
Abstract
Chondrogenesis is the process of differentiation of stem cells into mature chondrocytes. Such a process consists of chemical, functional, and structural changes which are initiated and mediated by the host environment of the cells. To date, the mechanobiology of chondrogenesis has not been fully elucidated. Hence, experimental activity is focused on recreating specific environmental conditions for stimulating chondrogenesis and to look for a mechanistic interpretation of the mechanobiological response of cells in the cartilaginous tissues. There are a large number of studies on the topic that vary considerably in their experimental protocols used for providing environmental cues to cells for differentiation, making generalizable conclusions difficult to ascertain. The main objective of this contribution is to review the mechanobiological stimulation of stem cell chondrogenesis and methodological approaches utilized to date to promote chondrogenesis of stem cells in vitro. In vivo models will also be explored, but this area is currently limited. An overview of the experimental approaches used by different research groups may help the development of unified testing methods that could be used to overcome existing knowledge gaps, leading to an accelerated translation of experimental findings to clinical practice.
Introduction
Chondrogenesis includes the differentiating process by which stem cells undergo chemical, functional, and structural changes to become mature chondrocytes. It begins with the aggregation of the chondroprogenitors into condensations, which is signaled by the presence of transforming growth factor beta (TGF-β) [1]. Chondrocyte differentiation and proliferation is marked by increases in chondrogenic gene expression, such as SOX9, aggrecan, and collagen II [1]. Levels of other biomarkers can indicate poor chondrogenesis, or hypertrophy, such as increased expression of collagen I and X [1].
It is well known that chondrocytes are sensitive to their mechanical environment. As in vivo chondrocytes function within an environment constantly subjected to a range of mechanical loading stimulations, these conditions are thought to accelerate chondrogenesis of progenitor cells [2 –4]. Thus, these conditions are mimicked in in vitro experimentation. A range of studies have shown how mechanical stimulation can be used to modulate cell metabolism. In particular, the type and magnitude of mechanical load may affect proliferation and apoptosis rates, as well as gene expression in chondrocytes and differentiation of stem cells.
To date, the mechanobiology of chondrogenesis has not been fully elucidated, despite the numerous studies published. Specifically aimed at providing mechanistic interpretation of the mechanobiological response of cells in the cartilaginous tissues, in vitro studies on cells or on tissue explants have been carried out. However, these studies vary considerably in their experimental protocols used for load application (eg, static vs. dynamic) and type of load [eg, mechanical vs. chemical using hydrostatic pressure (HP)] making generalizable conclusions difficult to ascertain [2 –4]. Moreover, the development of organ-on-a-chip technology provides a closer look at the interactions of cells with their environment [5 –8]. However, even in this case, experimental protocols for mechanical loading greatly vary from one study to another.
Research has also been conducted on animal models and human subjects to further test mechanobiological driven research hypotheses and potential treatments for cartilaginous degenerative diseases [2 –4]. Similar to in vitro studies, the methods adopted to modulate the mechanical environment of cells varied between passive (through robotic systems and/or external manipulation) and active loading mechanisms (eg, specific rehabilitative protocol) [2 –4].
The objective herein is to review the mechanobiological stimulation of stem cell chondrogenesis and methodological approaches utilized to date to promote chondrogenesis of stem cells in vitro. The limited number of current in vivo studies will also be explored, mainly in terms of future directions for the field of cartilage regeneration. In turn, the adoption of standard testing methods could be used to overcome existing knowledge gaps leading to an accelerated translation of experimental findings to clinical care. Current research on the optimal culture conditions for chondrogenic differentiation seeks to elaborate on the principles of environmental cues and cell cultures. The studies reviewed henceforth represent recent work and novel discoveries in regard to the effects of these factors on the chondrogenesis of stem cells. The different techniques for simulating chondrogenic mechanobiology, as well as their success in inducing chondrogenesis, will be explored.
Cell Environment
Earlier studies of various types of stem cells have shown that their differentiation varies according to mechanical cues from their culture environment [9]. In embryonic development, both intrinsic and extrinsic forces influence cell fate. Intrauterine breathing creates tensile forces in the bronchial epithelium, which supports smooth muscle development in the lungs [10]. The development of cardiac tissues and fetal hematopoiesis are influenced by shear forces from maternal blood flow [11,12], which in turn signals kidney morphogenesis through the primary cilia [13,14]. Finally, stress, strain, HP, and fluid flow have all been shown to contribute to proper ossification and bone collar formation in limb development [15].
Studies of the effects of chemical immobilization of chick embryos have proven that mechanical stimulation is vital for chondrogenesis and joint development [13,14]. Compared to embryos that were allowed to develop normally, immobilized embryos showed impaired joint cavity formation, abnormal limb shape, and failure to form synovial joint elements when paralysis was induced during early development [16]. When immobilized at later points of development, the chick embryos displayed a decrease in cartilage matrix production, cartilage volume, and hyaluronan content of articular surfaces, as well as decreased mechanical properties [17].
The differentiation of progenitor cells is heavily influenced by the mechanical properties of their culture environment, with the most notable property being matrix stiffness. In a study by Zeng et al., the addition of stiffening components to fibrous scaffolds seeded with porcine chondrocytes led to increased new cartilage formation in vivo after 16 weeks, compared to fibrous scaffolds that did not contain stiffening agents [18]. When substrate stiffness is similar to that of brain, muscle, or bone tissue, mesenchymal stem cells (MSCs) have been shown to differentiate into neurons, myoblasts, and osteoblasts, respectively, demonstrating that the mechanical properties of the culture material also influence MSC fate [14]. In general, osteogenic differentiation is more likely to occur in elastic materials with a storage modulus (G′) between 25 and 40 kPa, and differentiation of brain cells occurs in the softest matrices, with a G′ of less than 1 kPa. Chondrogenic differentiation has been shown to occur at intermediate substrate stiffness, and ongoing research has aimed to pinpoint the ideal matrix stiffness for chondrogenesis [9].
Recent studies have observed enhanced chondrogenesis across a wide range of matrix storage moduli. In static cultures, MSCs cultured in hydrogels of varying stiffnesses show the highest expression of chondrogenic markers in the stiffest hydrogels, with G′ values of 21 and 25 kPa [19,20]. When combining different matrix stiffnesses with cyclic HP, chondrogenic markers were also increased in stiffer matrices (17.5 kPa), but better morphological outcomes were observed in softer matrices (5.2 kPa) [21]. These results are supported by an in vivo study of bone-marrow derived stromal cell (BMSC) laden hydrogels implanted in mice after a 2-week preculture, with enhanced chondrogenesis observed in the stiffer constructs that had a G′ of 10 kPa compared to those with a G′ of 1 kPa [22].
Although the previous studies point toward an ideal G’ for chondrogenesis in the kPa range, a study by Feng et al. examining the effect of crosslinking (CL) density of microgels prepared three gelatin-hyaluronic acid (Gel-HA) systems with average G′ values of 238 Pa (low CL density), 510 Pa (medium CL density), and 772 Pa (high CL density). After a 4-week culture, all three gels produced high cell viability, with BMSCs showing trends of differentiating into hyaline cartilage in gels with low crosslinking density and fibrocartilage in gels with medium and high crosslinking densities [23]. This indicates that further research should be conducted to determine the ideal G′ ranges for differentiation of different types of cartilage.
Another important matrix property is the 3D network microstructure. In a study by Yang et al., fibrous and porous collagen hydrogels with identical chemical composition and similar mechanical strengths demonstrated varied effects on BMSC culture. Both in vitro and in vivo experiments suggested that the fibrous network hydrogel facilitates better chondrogenic differentiation and may slow down extracellular matrix (ECM) calcification [24]. Further research should seek to confirm these results and identify the best combination of network structure and mechanical strength for successful chondrogenesis.
Environmental Cues to Chondrogenesis
This section provides a comprehensive review of chondrogenic responses of stem cells to various physical stimuli. A summary of previous findings is provided in Table 1.
Chondrogenic Responses of Stem Cells to Environmental Cues
ADSC, adipose-derived stem cell; BMSC, bone-marrow derived stromal cell; MSC, mesenchymal stem cell; iPSC, induced pluripotent stem cell; HP, hydrostatic pressure; RCCS, rotary cell culture system; TGF-β, transforming growth factor beta; PGA, polyglycolic acid.
Dynamic/intermittent versus static cultures
During daily activities, articular cartilage experiences a combination of extrinsic and intrinsic compressive, tensile, and shear forces [25,26]. In healthy individuals, major load-bearing joints such as the knee and hip are subjected to thousands of compressive loading cycles per day [27]. For example, compressive forces can be static (up to 1 MPa while standing) or dynamic (up to 20 MPa when jumping) [28]. The presence of static pressure is also necessary within joints to control the growth and differentiation of chondroprogenitor cells into chondrocytes. If proliferation and ECM deposition continue to increase without the introduction of sufficient surrounding pressure, such as provided by the perichondrium in developing tissues, an accelerated hypertrophy may occur, leading to ectopic cartilage formation [25,29,30]. It then follows that dynamic stem cell cultures may be more suitable than static cultures to mimic the in vivo conditions that both promote and regulate chondrogenesis.
Dynamic loading on human chondrocyte cultures has been shown to increase cartilage matrix deposition and improve the mechanical qualities of engineered cartilage tissues [31]. In uniaxial loading studies, chondrocytes have been known to respond positively to moderate dynamic compressive loading in vitro, in the range of 2%–10% strain [32,33], 0.5–1.0 MPa [34,35] applied at 0.1–1.0 Hz [25,26]. In contrast, HP and shear loading have produced mixed effects on chondrocyte macromolecule synthesis and metabolism, respectively [36 –40]. More recent studies simulating multiaxial loading by combining compression and shear loading have observed superior chondrogenesis and increased glycosaminoglycan (GAG) and collagen II deposition compared to static culture [41], and even the suppression of factors that contribute to joint destruction in osteoarthritis (OA) [42]. In dynamic in vitro cultures, loading methods combined with chondrogenic induction media mimic the conditions that differentiating chondrocytes are subjected to in vivo. The benefits of dynamic culture over static culturing have previously been observed for inducing chondrogenesis in stem cells [43].
In a review by Fahy et al., studies of human, rabbit, and bovine MSCs all demonstrated increases in chondrogenic markers when the cells were exposed to cyclic compression, HP, or a combination of cyclic compression and surface shear [13]. Mechanical agitation of 3D cultures has corroborated these results, with the added benefits of increased proliferation and chondrogenic differentiation of BMSCs compared to 2D cultures [44]. Loading methods, when combined with chondrogenic induction media, mimic the conditions that differentiating chondrocytes are subjected to in vivo. Mechanical agitation of 3D cultures has corroborated these results, with the added benefits of increased proliferation and chondrogenic differentiation of BMSCs compared to 2D cultures [44].
Several groups have sought to evaluate whether dynamic culture conditions would enhance chondrogenesis, rather than how preculturing in static conditions would affect the outcome. As such, some recent studies have shifted toward determining if allowing for a static differentiation period before dynamic culture is beneficial, and whether there is an ideal amount of priming time which will best encourage chondrogenesis and cartilage maturation. Subjecting cultures to 7 days of cyclic HP or dynamic compression after a 7- or 21-day preculture, respectively, showed enhanced chondrogenesis and suppressed hypertrophy compared to loading without a preculture [21,45]. McDermott et al. varied the priming times before dynamic compression even further and found that shorter priming times may preserve chondrocyte homeostasis, while longer priming times may support cartilage maturation [46]. In all cases, applying loading at the beginning of a culture period tended to drive cells toward osteogenesis, while allowing for a priming period encouraged chondrogenesis, suggesting that priming time does affect the effectiveness of physical loading.
Compressive loading and strain
Some groups evaluated the effect of unconfined compressive strain on chondrogenesis. This loading may impact chondrogenesis through the entraining of the molecular clock within cells [47]. For these compression experiments, samples were exposed to a strain with a magnitude of 2%–20% (or less than 10 kPa) at a frequency of 1 Hz or lower [45,47 –59]. The strain cycle was (1) cyclic through sinusoidal loading [45,47 –49,52,54,58,59], (2) dynamic strain superimposed on static loading [51,53,55], or (3) a brief, initial static preload followed by a longer period of sinusoidal loading [50,56]. Custom-made bioreactors within incubators were used to generate the compressive forces, several of which were driven by air pressure [45,47 –57,60 –62]. Groups tested the ideal length of time that cell cultures should receive loading, which was around 1–4 h per day [49,57].
In addition, it was determined that cells should not be placed in a loading environment until after they have had adequate time to undergo chondrogenic differentiation [45,53]. Delayed time point dynamic compression (beginning around day 20) enhanced chondrogenesis and suppressed hypertrophy, while early time point compression suppressed chondrogenesis [45,51,53]. Temperature increases from compression are an unavoidable by-product of loading. Nasrollahzadeh et al. examined this widely neglected effect and found that the coexistence of thermomechanical cues had a superior effect on chondrogenic gene expression compared to either signal alone [62].
In addition, it seems that the need to add TGF-β to cultures is dependent on the type of stem cell used [49]. For example, MSCs and induced pluripotent stem cells (iPSCs) were found to show increased expression of cartilage-related markers in response to loading, independent from the presence of TGF-β [48,49,58,61]. Yet, it is believed that addition of TGF-β would only synergistically improve the chondrogenic response, to an extent [51,53,58,61]. However, embryonic stem cells required the addition of TGF-β to the existing loading to achieve these positive results [49].
Both the type of hydrogel and the concentration of cells within it are also important components of load testing. Scaffolds with higher dissipation levels lead to greater upregulation of chondrogenic markers [56]. As shown by Pelaez et al., fibrin gel is suitable for supporting cyclic compression-induced chondrogenesis of MSCs, and the gel works optimally at a concentration of 40 mg/mL [50]. Bian et al. concluded that MSCs in hyaluronic acid (HA) hydrogels with higher seeding densities (60 million cells/mL) led to increased mechanical properties and chondrogenesis [55].
Studies of shear strain induced by mechanical agitation of 3D cultures [44] or through low magnitude, high frequency vibration [63] supplemented with chondrogenic induction media have resulted in enhanced chondrogenesis within their dynamic culture groups. Surface shear strain superimposed on axial compression also has the potential to enhance chondrogenesis.
A 32 mm diameter ceramic hip ball is part of a bioreactor that is used to apply this complex dual loading to cells [64 –69]. The ball is pressed onto a cell-seeded scaffold and oscillates at ±25° at 1 Hz about an axis perpendicular to the scaffold axis, producing interface shear motion [64 –69]. This oscillation is in addition to the normal 1 Hz 10%–20% sinusoidal strain and/or 10% static strain [64 –69]. A representation of this type of loading apparatus is depicted in Fig. 1. Results showed that this type of loading promotes chondrogenesis of MSCs through the TGF-β pathway by upregulating TGF-β gene expression and protein synthesis [64,66,69]. The combination of compression and shear leads to significant increases in chondrogenesis, while either stimulation alone was insufficient [67]. In addition, asymmetrically seeded scaffolds led to improved tissue development [68].
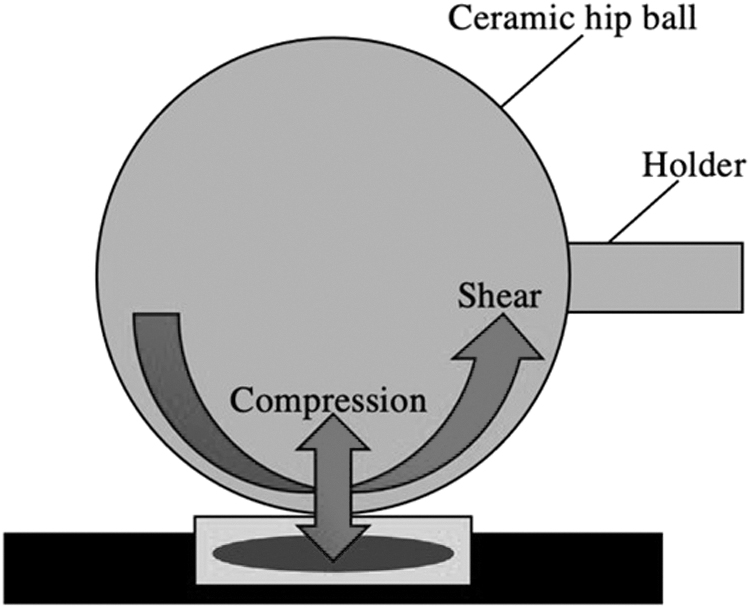
A compressive loading and strain bioreactor: A 32 mm diameter ceramic hip ball bioreactor is held by a mobile metallic holder. The hip ball is pressed against an MSC-seeded scaffold and compresses the cell seeded scaffold dynamically at a strain amplitude that is a percentage of its height. It also oscillates at ±25° perpendicular to the scaffold to produce mechanical shear stress. The resulting complex dual loading upregulates TGF-β expression to increase chondrogenesis significantly. MSC, mesenchymal stem cell; TGF-β, transforming growth factor beta.
A slight alteration to the ceramic hip ball bioreactor is the metal roller [70]. This device can also apply both shear and cyclic compressive forces to the cell culture [70]. Results showed that this bioreactor upregulates positive chondrogenic markers only when both shear and compressive forces are administered [70].
Hydrostatic pressure
Repetitive HP is one component of the mechanical stresses present within joints, and the physiologic pressure range for humans is 3–10 MPa [71,72]. HP does appear to induce chondrogenesis in vivo, where chemical signals for multiple differentiation pathways are present [73]. Thus, in vitro HP can mimic the natural joint loading environment [74]. To create this dynamic mechanical stimulation, groups followed a similar experimental setup. Cell aggregates were placed in tubes with flexible caps that were then placed in the stainless steel HP vessel within a water-filled temperature-controlled incubator bath (set at 37°C) or aggregate pellets or tubes were placed in sterile plastic bags before being placed into the chamber (Fig. 2) [21,71 –85].
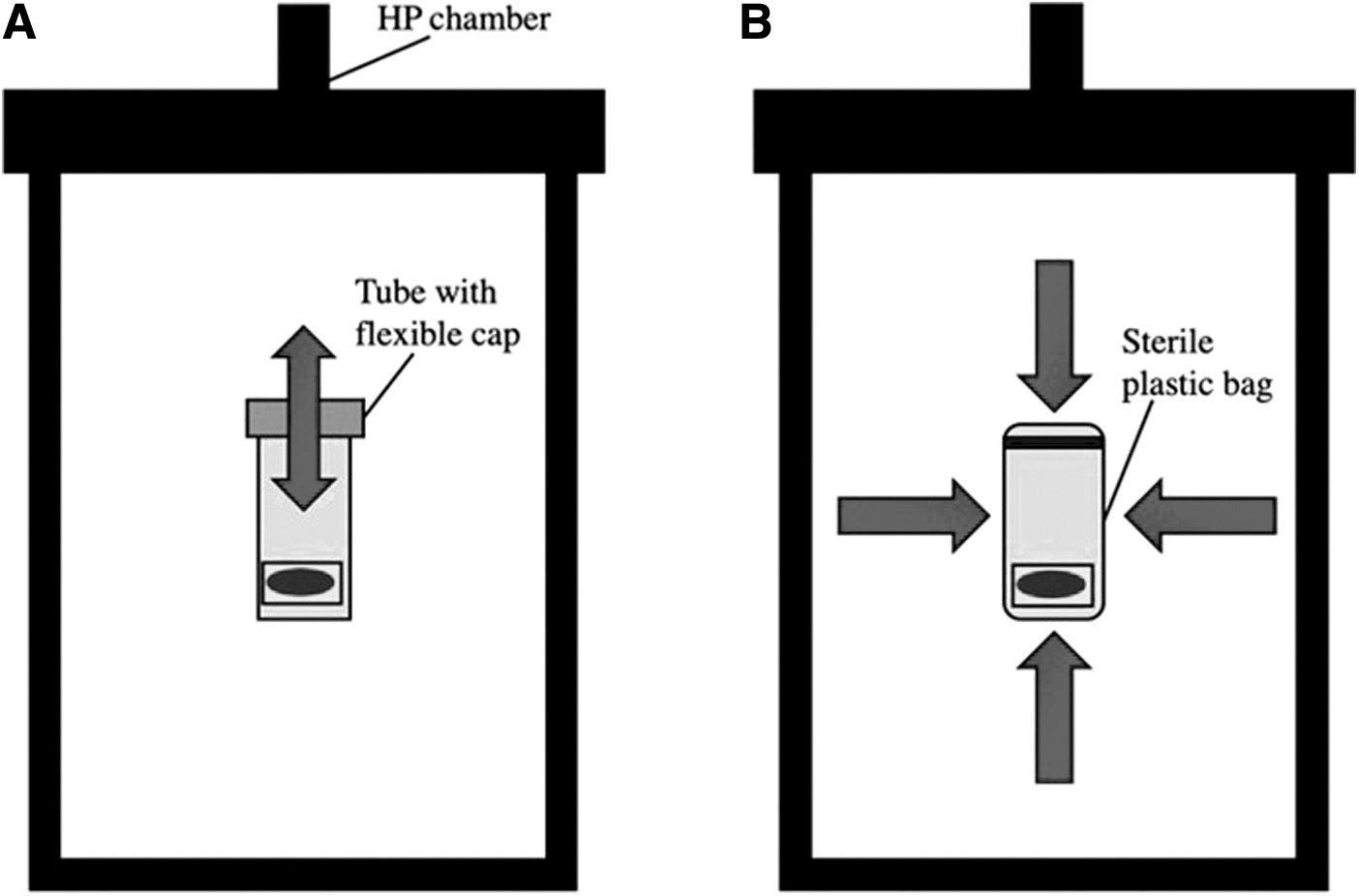
Examples of HP bioreactors:
Some systems were powered by electromagnets [82,86]. Cyclic HP was consistently applied for 4 h per day at a frequency of 1 Hz [21,71 –82,85]. Although early studies examined a range of pressures from 0.1 to 10 MPa [71,75,77], by 2006 most studies chose to use (1) under 5 MPa [21,73,74,80 –82,87,88], (2) 7.5 MPa [76,78], or (3) 10 MPa [72,79,83 –85,89]. As shown by Finger et al. in 2007, this pressurization could either be instantly achieved or progressively ramped up to the final intended value in daily increments, with the steady application producing better effects [76]. In addition, HP could either be applied immediately on day 0 of culture or delayed to a later day further along in the differentiation process of the cells. Interestingly, delayed application of HP resulted in improved chondrogenesis [21,89].
Finally, the stiffness of the scaffold in which cells are encapsulated in also likely plays a role in the results, with softer hydrogels mimicking the in vivo cartilage matrix [21,83,84]. Even the type of stem cell and the donor have been shown to influence how loading impacts chondrogenesis [85,87]. Only two studies were identified in which static pressure was generated within a pressure chamber. In these reports, done by the same group, cells were loaded into wells and transferred into the machine producing 50–200 kPa of static pressure for 0–4 h [90,91].
Many groups sought to develop an in vitro loading system that could examine the effect of cyclic HP on a particular line of stem cells undergoing chondrogenesis with adequate growth medium [21,71,73 –75,77,80 –89,92,93]. Results showed that multiday loading could enhance proteoglycan and collagen production of the differentiating stem cells compared to unloaded controls [71,73,75,81,82,88]. Chondrogenic markers such as SOX9, type II collagen, and aggrecan levels were upregulated to different degrees by different loading pressures [73,74,77,82,86]. Size changes were noted in HP samples, with these constructs being more uniform and compact [71,73,75,77]. Similar results were obtained with static pressure, as chondrogenic marker expression increased for pressures of around 100–150 kPa applied for 3 h [90,91]. However, all of these studies only looked at HP indirectly since conditions that promoted chondrogenic differentiation were used. Thus, other groups looked at whether cyclic HP is a functional stimulus for chondrogenesis, either alone or in the presence of TGF-β [72,76,78,79].
Even without TGF-β, groups found that stem cells exposed to cyclic HP showed early increased SOX9, type II collagen, and aggrecan levels [72,78,87,88]. Effects were additive if TGF-β was added, with greater concentrations producing better effects to an upper limit [72,79]. Finger et al. found that HP without TGF-β initially increased collagen I expression, then later increased SOX9, suggesting that HP alone may be sufficient to begin the induction of chondrogenesis in stem cells [76]. Puetzer et al. came to a similar conclusion, as chondrogenic expression levels were only upregulated with HP for the first 7 days of loading [78]. In contrast, Vinardell et al. concluded that HP alone cannot induce robust chondrogenesis of stem cells, but can act synergistically with TGF-β to increase chondrogenic differentiation and inhibit collagen type X [79]. Still, HP, with or without TGF-β, was also shown to create cell pellets that were more compact and uniform in structure [72,79].
In addition, HP superimposed on another mechanical stimulation, such as fluid shear stress, upregulates chondrogenesis more than either loading alone [86,87,92]. This is likely due to complex loading patterns that better mimic the native in vivo cartilage environment.
Electromagnetic stimulation
Magnetic forces do have excellent prospects for application as a noninvasive physical stimulation for chondrogenic differentiation [94]. These forces can be used to power mechanical stimulation bioreactors [95,96]. They can also be incorporated into nanoparticles that are internally incorporated into cell cultures and stimulated by external magnetic fields [97,98]. The external magnetic forces that are directly applied to stem cell constructs, with or without magnetic nanoparticles, can be generated by either (1) static magnetic fields (SMFs) or (2) pulsed magnetic fields (PMFs). These two stimulation techniques are discussed below.
Biophysical forces such as SMF have attracted interest for their effect on cartilage regeneration [99]. SMF changes the structure and function of cells by exposing them to permanent magnets that are stable in direction and intensity [94]. Although some reports at the time suggested that SMF may be useful in stimulating chondrogenic differentiation of precursor cells, a study by Amin et al. in 2014 provided a detailed look into the chondrogenesis-promoting in vitro technique with adult human BMSCs [99]. The group cultured BMSCs in a SMF of 0.1, 0.2, 0.4, and 0.6 T of SMF for 3 weeks, and they also exposed samples to 0.4 T SMF for 1, 2, or 3 weeks since this field strength was shown to be optimal in previous reports [99]. Results showed that SMFs stimulated sulfated-GAG (sGAG) production (a key indicator of terminal chondrogenic differentiation) and collagen II deposition, and maximal effects were seen in the 14–21 days 0.4 T groups [99].
In addition, SOX9, aggrecan gene (ACAN), and collagen type I gene (COL2A1) expression levels were elevated in SMF conditions compared to controls, and the TGF-β pathway was even stimulated by SMF [99]. Thus, a moderate-strength SMF promoted chondrogenic differentiation of BMSCs [99]. Zhang et al. found similar results in 2021 when the group investigated the effects of another moderate-strength SMF on the chondrogenesis of a mandibular bone marrow mesenchymal stem cell (MBMSC)/mandibular condylar chondrocyte (MCC) coculture [94]. The group used a 0.280 T SMF and found that this regimen promoted proliferation of cells, as well as chondrogenesis [94]. Within a week, the cells displayed Alcian blue staining and elevated SOX9, COL2A1, and ACAN expression [94].
In addition, bioelectricity plays a key role in a multitude of physiological processes, with recent evidence suggesting that cells respond to bioelectric signals during embryonic development and regenerative repair [100]. Current research on the electromagnetic mechanobiology of stem cells has centered around finding the ideal combination of electrical and magnetic field stimulation for enhanced chondrogenesis in vitro.
In 2013, Chen et al. examined a pulsed electromagnetic field (PEMF) and a single-pulsed electromagnetic field (SPEMF) to compare chondrogenic induction of adipose-derived stem cells (ADSCs) cultured in both 2D and 3D [101]. For the PEMF condition, the following characteristics were implemented: magnetic flux density of 2 mT, frequency of 15 Hz, magnetic field of 20 G, and pulsed period of 67.1 ms, all for 8 h per day [101]. For the SPEMF condition, the following characteristics were implemented: period of 5 ms, magnetic field magnitude of 0.6–1 T, magnetic flux density of 1 T, all for 3 min per day [101]. In both the 2D and 3D conditions, both PEMF and SPEMF stimulation increased chondrogenic marker genes and increased sGAG synthesis [101]. The group concluded SPEMF to be a superior treatment since it had reduced administration times and did not induce osteogenesis [101].
In 2019, Huang et al. expanded the knowledge of PEMF by examining its combined benefit with hydrogel embedded with magnetic nanoparticles [102]. The group found that the hydrogel together with the PEMF treatment promoted chondrogenesis, as evidenced by strong Toluidine blue staining and increased collagen II, SOX9, COL2, and ACAN expression [102]. It was hypothesized that these positive results were due to PEMF enhancing the cellular interactions and nutrient perfusion [102].
Also adding to the knowledge of the effects of PEMF on stem cells, Parate et al. varied the field amplitude, exposure duration, and dosage to determine the briefest and lowest amplitude exposure to produce a significant outcome [103]. MSCs were subject to 0–4 mT for 10 min, with 2 mT producing maximal effects in terms of SOX9, aggrecan, and COL2 expression, leading this strength to then be tested for 5–60 min [103]. The 10 min treatment proved to be best, and it was also found that the PEMF produced the best effects (increased collagen II levels) when administered once per day [103].
Celik et al. expanded on the work of Parate et al. by examining the influence of directionally applied PEMFs on chondrogenic differentiation of MSCs onto topographically-variant scaffolds [104]. MSCs were exposed to 1 mT of PEMF for 10 min per day in the X, Y, and Z directions (Fig. 3) [104]. A testing protocol of 1 mT and 10 min per day was chosen because these parameters optimized the upregulation of SOX9, COL2, and ACAN [104]. Results showed that PEMF in the Z-direction elevated cartilage markers, while the X-direction had little effect and the Y-direction inhibited chondrogenesis [104]. Most recently, as reviewed by Varani et al., rabbit, bovine, and human MSCs have shown enhanced chondrogenic differentiation, paracrine activity, migration, and proliferation in response to PEMFs at various magnitudes and frequencies.
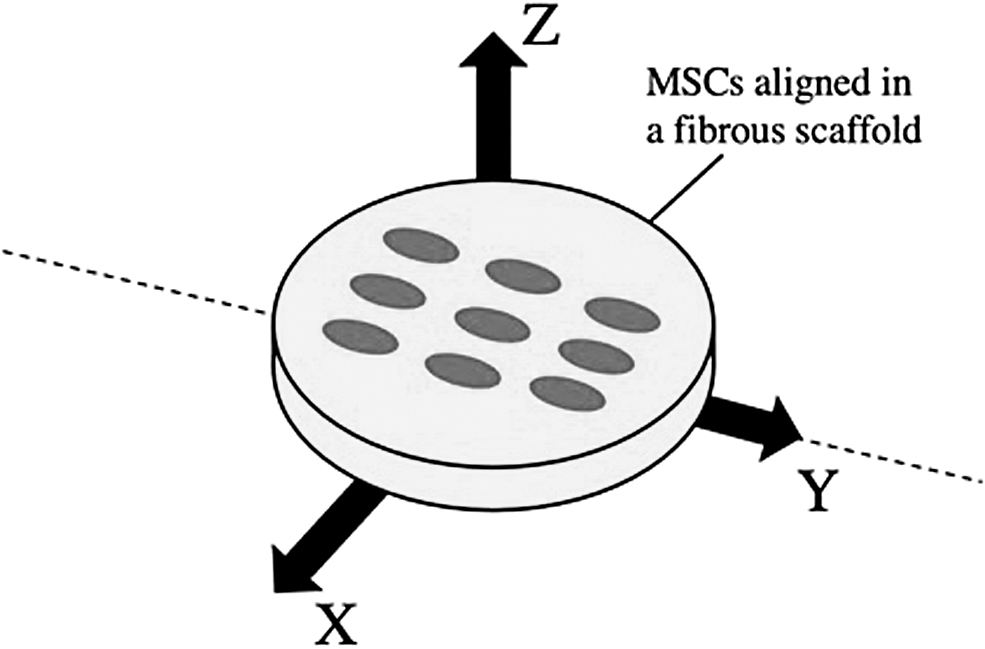
An electromagnetic stimulation bioreactor: The directionality of the magnetic field influences chondrogenesis for MSCs in a fibrous scaffold of different alignments. For an aligned scaffold, Y-directed fields (parallel to cells) demonstrated strong chondrogenic inhibition. X-directed fields (perpendicular to cells) had no effect on chondrogenesis when cells were aligned and did increase hypertrophy. Z-directed fields significantly increased chondrogenic expression, but only for MSCs randomly aligned on a scaffold.
Yet, the impact of SMF, PMF, SPEMF, and PEMF is not wholly proven. Electromagnetic stimulation has been found to both enhance and inhibit chondrogenesis [104 –106]. In a study with rat BM-MSCs, PEMF exposure resulted in the inhibition of maintaining a cartilaginous phenotype [105]. While both exposure to electrical fields and the use of conductive polymer scaffolds have led to increased chondrogenesis separately [100,107], these results are yet to be expanded to in vivo conditions, and the effects of external electrical stimulation in conjunction with conductive scaffolds are yet to be investigated.
In studies focusing on magnetic field stimulation, the effectiveness of SMF or PMF exposure at encouraging chondrogenesis has been mixed. One study found that even after allowing for a chondrogenic induction period, SMF exposure only increased chondrogenesis in human ADSCs, while it decreased chondrogenesis in Wharton Jelly MSCs, demonstrating that not all types of stem cells react equally to MF exposure [106]. Another study focusing on the directionalities of magnetic fields found that Z-directed fields had the best effect on chondrogenesis, while X-directed fields inhibited chondrogenesis in randomly oriented scaffolds. Interestingly, the same study observed that PEMFs did not promote chondrogenic differentiation on either polystyrene tissue culture plastic or aligned scaffolds, irrespective of field directionality, and that multiple PMF exposures could also inhibit chondrogenesis in all types of scaffolds tested [104].
Dikina et al. found in 2017 that neither static nor variable magnetic fields generated by 1.44–1.45 T permanent magnets affected cartilage formation in a scaffold-free MSC sheet [98]. Even those samples that used magnetic nanoparticles did not make an impact on chondrogenesis [98]. These results suggest that the success of chondrogenesis in response to MF exposure depends on many different factors. Overall, further research should be conducted to determine ideal stimulation parameters, categorize the effects of MF exposure on the chondrogenesis multiple types of stem cells, and to confirm the efficacy of the conditions presented above for enhancing chondrogenesis.
Ultrasound signaling
Ultrasound is a mechanically vibratory form of acoustic wave energy that travels through a given medium [108,109]. Over the past few decades, medical use of ultrasound has been extended beyond imaging and diagnosis toward therapeutic applications [110]. Ultrasound can be used to induce piezoelectric stimulation, which has been shown to facilitate MSC chondrogenesis [111]. Yet, most notably, low intensity ultrasound (LIUS) has been used to induce localized chondrogenic differentiation and enhance cell viability and cartilage matrix synthesis [112,113]. This type of ultrasound uses a frequency of 0.75–1.15 MHz and an intensity lower than 200 mW/cm2 [110]. These waves are characterized as either (1) continuous (cLIUS) or (2) pulsed (pLIUS). These two stimulation wave forms are discussed below.
All groups that used a cLIUS wave found that this biomechanical stimulation treatment can accelerate chondrocyte differentiation and improve cellular viability in vitro or in vivo [108,112 –121]. An example of this type of apparatus is provided in Fig. 4. In 2004, Ebisawa et al. tested a continuous signal with intensities of 15, 30, 60, and 120 mW/cm2, finding the 30 mW/cm2 treatment to produce the best effects [113]. Subsequent groups chose an intensity of 200 mW/cm2 [108,112,114,116,117] or less than 30 mW/cm2 [115,120]. Signal frequency and treatment time ranged from 0.8 MHz to 8.5 MHz and 1 min/day to 80 min/day, respectively [108,112 –121].
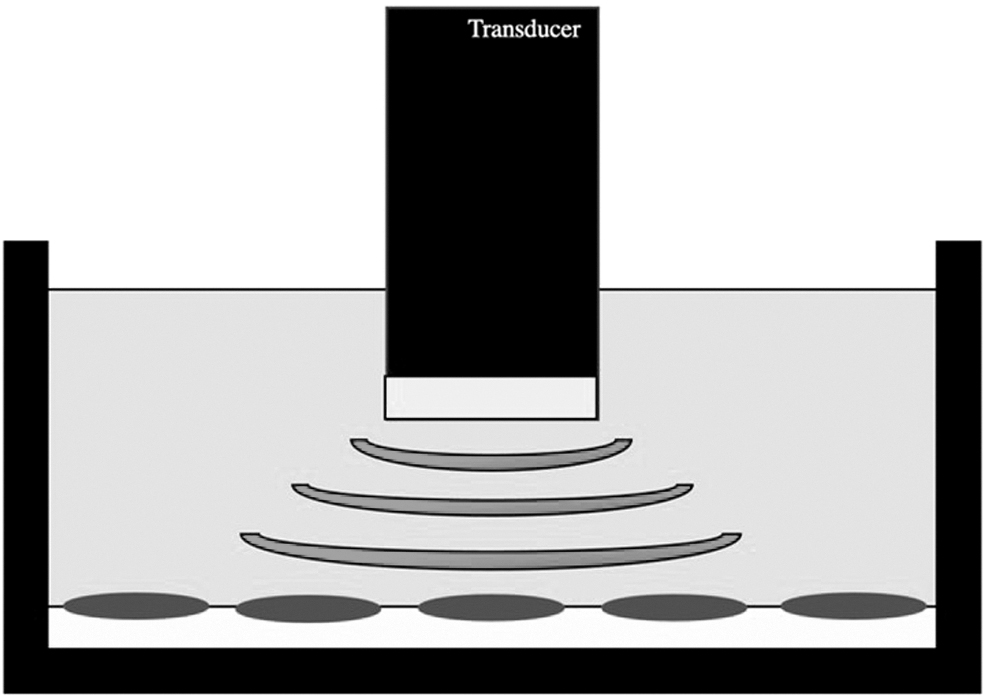
An ultrasound signaling bioreactor: A transducer emits the low-intensity ultrasound waves through the culture medium to stimulate the cell-laden hydrogel scaffold or cell pellet culture. The transducer surface is aligned perpendicularly to face the cells. Treatment is either pulsed or continuous and varies in terms of intensity, frequency, and time per day.
Results for cLIUS treatments showed that chondrogenesis was significantly upregulated, as evident through increased GAG content [108,112,116], ECM synthesis [108,116], aggrecan expression [108,114 –116], SOX9 expression [114,116,118 –121], collagen type II content [112,114 –117,119 –121], and cellular viability [114,119]. Osteogenic differentiation, as measured by markers for type X collagen, was also downregulated [116,117]. Yet, there are conflicting findings on whether cLIUS can act in the absence of TGF-β. Ebisawa et al. reported in 2004 that application of cLIUS alone showed little chondrogenic enhancement [113]. They found that cLIUS could only enhance the effects of TGF-β mediated chondrocyte differentiation [113]. Yet, experiments conducted by others showed that cLIUS treatment could positively impact chondrogenesis in the absence of exogenously added growth factors [114,118,119]. cLIUS was shown to perhaps even be better than TGF-β treatment, which results in unwanted production of the osteogenic collagen type X [117,121].
pLIUS uses low energy and pulsates it with very short intervals to produce pulsed pressure waves [122]. Similar to cLIUS, pLIUS is also shown to enhance chondrogenic differentiation and repair [109,110,122 –127]. Most groups chose an intensity of around 20–50 mW/cm2 [109,110,122,123,125 –127] or 200 mW/cm2 [124]. A frequency of 1.5 MHz was commonly used, with 200 μs-burst sine waves repeated at 1 KHz [110,122,123,127]. Yet, some groups did use 1–5 MHz [109,124,125] with a 100 Hz repetition rate [124]. pLIUS treatments were administered once per day, for 20–40 min/day [109,110,122 –127]. Results showed that pLIUS promotes the proliferation and stimulates differentiation of chondrocytes [110,122,123]. Expression of collagen type II [122,123,127], aggrecan [122], ECM [123],7 SOX9 [127], and proteoglycans [110,123] was increased, while collagen type X levels decreased [122]. However, most groups determined that TGF-β was required for pLIUS to produce these positive effects [109,122,124,126].
Alternative Cell Culture Approaches
This section provides a comprehensive review of alternative cell culture approaches. A summary of previous findings is provided in Table 2.
Alternative Cell Culture Approaches
On a chip
Microfluidic-based microdevices accelerate the execution of experimental testing by decreasing the sample size and platform and better controlling the surrounding microenvironment compared to conventional methods. These devices have been used to examine the effects on extrinsic factors, such as mechanical stimulation, on cell fate. Recently, several groups have used this technique to examine the effect of (1) shear stresses or (2) strains on the chondrogenesis of stem cells.
Shear stresses within the microfluidic device can potentially play a role in chondrocyte differentiation, as demonstrated by Zhong et al. in 2013. These stresses mirror fluid flow stimuli within the cartilage matrix in vivo that regulates cell behavior and functions [128]. The microdevice used by Zhong et al. consisted of four cell culture chambers connected to different resistance channels, and fluid flow was driven by a pump (Fig. 5A) [128]. Both articular cartilage chondrocytes and primary MSCs were isolated from rats and injected into a cell chamber [128]. Average stresses administered to the cell cultures ranged from 1.089 to 0.009 dyne-cm2 [128]. Results showed that the proliferation rates of both chondrocytes and MSCs increased with increasing magnitude of fluid shear stress [128]. However, SOX9 levels in the differentiating MSCs were not altered for the different shear stresses [128]. With increasing levels of fluid flow, chondrocytes began to synthesize collagen I, while collagen II expression diminished [128].
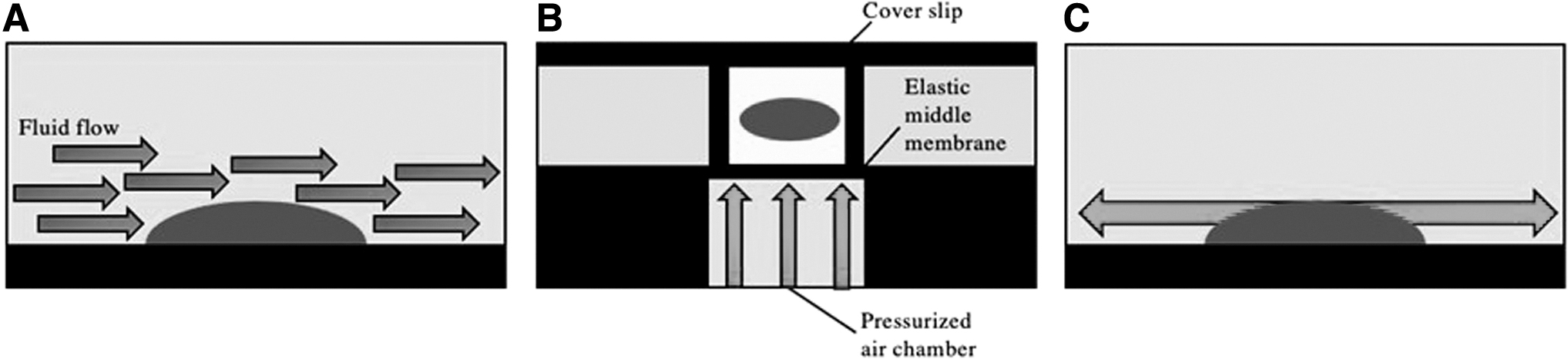
Examples of organ on a chip bioreactors:
In 2018, Kowsari et al. published the first known report on the effect of different uniaxial strains on chondrogenic differentiation [129]. The group used a novel semiconfined design that applied a wide range of static and dynamic mechanical compressive strains to cell-laden cylindrical hydrogels [129]. The microbioreactor is a 5 × 4 well array of 20 actuation cavities that enable simultaneous various compressive strains by a single pressurized air source [129]. When pressure in the air chamber of the microdevice increases above atmospheric pressure, an elastic polydimethylsiloxane membrane moves upwards and creates uniform cyclic axial strain in the hydrogel [129]. A diagram of this organ on a chip model is given in Fig. 5B.
Numerical methods and equations were used to ensure that the strain distribution was uniform and the microenvironment was suitable for the vitality of the cell culture [129]. For testing, the group used ADSCs from rabbits that were expanded and mixed with an alginate-sodium solution before being poured into the microwells of the microdevice [129]. Loading occurred for 2 h a day, 6 days a week, at 1 Hz with strain magnitudes of 20%, 15%, 10%, 5%, and 0% in each row of the array [129]. Results showed that loading tended to increase cell viability compared to controls [129]. In addition, collagen II was maximally upregulated in the 10% strain condition, while aggrecan and SOX9 were more highly expressed in all of the loaded groups [129].
This publication was followed by a 2020 report by Chiu et al. that looked at chondrogenesis under uniform and nonuniform 2-axial strain in another membrane-type microdevice that was also driven by a pneumatic system [130]. The microdevice used 1:1 circular, 1:2 oval, and 1:3 oval culture wells to generate uniform and nonuniform strains on cultured rabbit periosteal cells within the wells (Fig. 5C) [130]. Results showed that cell viability was not reduced by loading and that SOX9 expression was upregulated when cells were stretched under 6 kPa pressure in the 1:3 oval well [130]. Thus, chondrocytes were differentiated from stem cells under a large and nearly single dimensional strain [130].
Shaking
To date, there is only one study that uses shaking culture methods for chondrogenic induction of stem cells. The 2020 study by Limraksasin et al. used 3D mouse iPSC cultures that were treated with hydrodynamic stress at specific frequencies in a simple see-saw shaker (Fig. 6) [131]. The single cell suspensions were added to low-cell attachment 96-well U bottom plates, where they were maintained for 24 h before chondrogenic induction medium was added [131]. After 3 days, the cells were transferred to low-cell attachment plates and subjected to suspension shaking culture at a frequency of 0 (static), 0.3, or 0.5 Hz [131].

A shaking bioreactor: The see-saw shaker bioreactor administers noncontact hydrodynamic stimulation to iPSCs in vitro. Shaking cultures at 0.3 and 0.5 Hz increased expression of chondrogenic-related genes (compared to static cultures) by increasing TGF-β expression and Wnt signaling. iPSC, induced pluripotent stem cell.
The frequencies of 0.3 and 0.5 Hz were chosen as the shaking frequencies because 0.1–1 Hz has been well established by previous studies on mechanical loading and the present apparatus did not allow for shaking above frequencies of 0.5 Hz due to medium overflow [131]. Results showed that the shaking cultures produced chondrogenically induced iPSC constructs that were larger in size, surrounded by more extensive ECM lacunae, and more differentiated and mature as determined by gene expression compared to static cultures [131].
Sliding contact
Physiological joint loading often includes sliding contact between two contacting cartilage layers, such as in the knee and hip [132]. This type of loading is characterized by moving contact and fluid pressurization [132]. Thus, in 2012, Huang et al. constructed a displacement controlled bioreactor that applies sliding contrast in both the X and Y directions to MSC-seeded hydrogel strips [132]. The cell-hydrogel strips were subjected to 5%–20% axial strain applied at a velocity of 2.5 mm/s [132]. Results showed that short term sliding contact improved chondrogenic gene expression [132]. However, this success was dependent on the axial strain applied and supplementation with TGF-β [132].
Microgravity
Both a 3D culture environment and appropriate mechanical stimulation are important for chondrogenic differentiation of stem cells to mimic in vivo conditions [133]. A rotary cell culture system (RCCS) is a 3D microgravity culture system that allows culture materials to establish a suspension track similar to a homogenous fluid on a horizontal axis (Fig. 7) [134,135]. It is thought to potentially induce chondrogenic differentiation, and the device was first used by NASA for microgravity testing [133,136,137]. In this machine's container, gravity, buoyancy, and shear force achieve a balance in a suspension orbit, which constitutes an environment conducive to cell aggregation [133,134]. A free fall status is maintained, similar to if the culture was in space, and cells attach to microcarriers cultured in a 3D scaffold material that can freely rotate with the rotation of the base [134,135,138]. Specific biomechanical stimuli, such as shear stress, joint compression force, and HP, that support tissue development are also provided by the RCCS system [135,139,140].
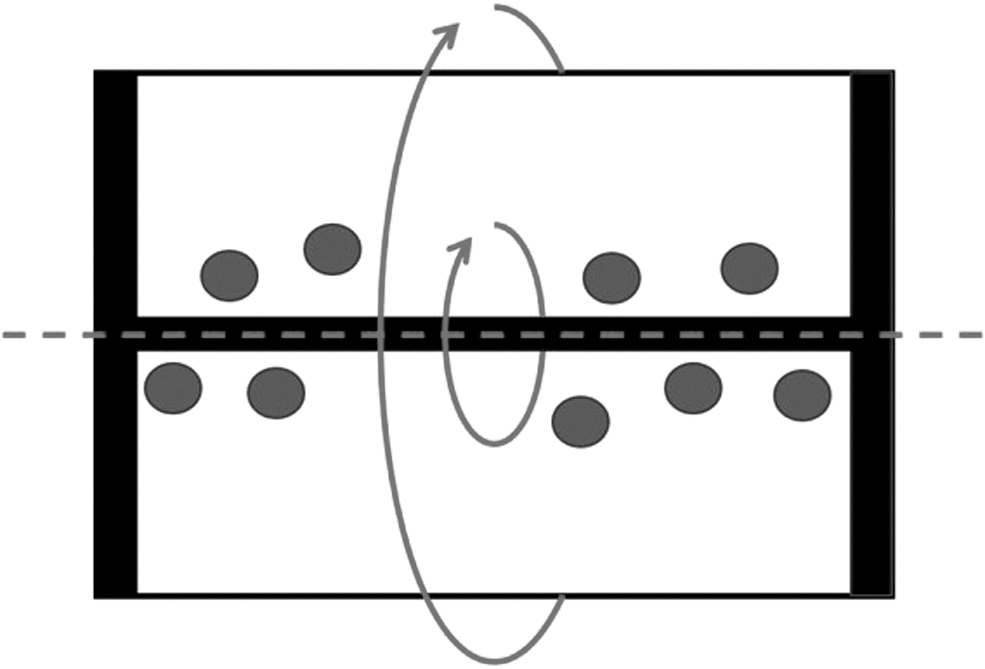
A microgravity bioreactor: The RCCS is used to rotate cell-microcarrier cultures or cell pellets and create a microgravity environment. Rotation speeds are increased over time to ensure that the cells do not come in contact with the bioreactor's walls and instead are in free-fall. SHH and IHH likely play a role in regulating chondrogenesis under microgravity conditions. RCCS, rotary cell culture system; SHH, Sonic Hedgehog; IHH, Indian Hedgehog.
All groups increased the rotation speed of the RCCS container from the initial speed over time to create a true microgravity environment. Several groups did this in a stepwise manner. These groups initially set the rotation speed of the container to 10–12 rpm to ensure full contact between the cells and their microcarriers [133 –135]. Then, after 24 h, the rotation speed was increased to 12–14 rpm [133 –135]. This was done so that the culture material in the container no longer came into contact with the container and remained in free-fall during rotation [133 –135]. Other groups increased rotation speed gradually as time progressed. Luo et al. initiated studies at a speed of 6.5 rpm, which was increased as aggregates expanded [141]. Mayer-Wagner et al. varied rotational speed between 4.2 and 7.2 rpm during experimentation to maintain cells in a free floating position [138,142]. Yu et al. varied rotation speed between 11 and 25 rpm and increased it steadily over time for 21 days [137]. Still, some groups began with a speed of 20–30 rpm, which was further increased over the course of the study [139,140].
Most reports on the use of an RCCS to generated microgravity deem that it is an effective technique for chondrogenesis induction in stem cells. The dynamic 3D culture seems to increase the growth rate and distribution of seeded cells [141,143]. In addition, the rotating action continuously transfers culture medium, gasses, and nutrients to the differentiating stem cells, producing higher metabolic activity and accelerated tissue growth [139,143]. Perhaps this explains why cells in microgravity conditions tend to be larger in size and express upregulation of type II collagen, some proteoglycans, and some chondrogenic signaling proteins [137,139 –141].
Some reports, such as that by Mayer-Wagner, still conclude that microgravity produces reduced expression of collagen II and aggrecan [138]. Although it is shown by some to suppress hypertrophic differentiation by reducing collagen X and its gene (COL10A1) [138], other reports show that RCCS does not produce these intended effects, as COL10A1 and matrix metalloproteinase 13 (MMP-13) expression was enhanced [139]. Thus, the chondrogenic and hypertrophic effects of microgravity are still quite disputed.
Several groups examined the effects of RCCS on Indian Hedgehog (IHH) and/or Sonic Hedgehog (SHH) induced chondrogenic differentiation to elucidate the role of microgravity in this process [134]. Chen et al. found that transfection of BMSCs with IHH and SHH induced a significant chondrogenic response over time in the RCCS group, compared to nonmicrogravity controls, as levels of collagen II and ACAN were elevated [134]. Similarly, the same group found that BMSCs transfected with SHH and cultured in RCCS resulted in significantly upregulated mRNA levels of SOX9, ACAN, and collagen II [133]. Liu confirmed that IHH transfected BMSCs in RCCS produced increased mRNA expression of collagen II and aggrecan compared to conventionally cultured cells [135].
The benefit of microgravity appears to be that chondrogenic hypertrophy and aging are alleviated during differentiation, which could be due to continued mechanical stress from medium flow and low shear stresses [134]. The free-fall environment is thought to facilitate cellular aggregation and enhance cell–cell and cell–matrix communication [135]. In the RCCS, cells and nutrients are homogenously distributed, improving the transport of nutrients and removal of metabolic wastes in the microenvironment and thus improving the culture [133 –135].
Yet, the chondrogenic effects of simulated microgravity are still conflicting, especially compared to more proven mechanical stimuli techniques such as electromagnetic forces (EMFs) and HP. Mayer-Wagner et al. sought to determine the single and combined effects of low-frequency EMF and microgravity on chondrogenesis of hMSCs [142]. Results showed that microgravity alone reduced collagen II, COL2A1, and aggrecan levels significantly, compared to EMF and combined groups [142]. In addition, Mellor et al. compared cyclic HP with simulated microgravity to better understand the effect of these stimuli on chondrogenesis of human adipose stem cells (hASCs) [136]. Again, results showed that the microgravity group had reduced aggrecan and sGAG compared to the HP controls [136]. Wnt signaling was increased in the microgravity condition, which has been suggested to inhibit chondrogenesis and stimulate chondrocyte hypertrophy [136].
Conclusion and Future Directions
Chondrocytes and chondroprogenitors transduce and respond to a wide array of in vivo environmental mechanical stimuli during development and throughout adulthood [144]. Joint loading produces direct stresses and strains, as well as indirect factors such as HP and fluid flow [144]. As described in this review, the various cell culture techniques that model these stimuli have shown much promise in inducing chondrogenesis in stem cells. Cell ECM metabolism on the gene level, including remodeling and organization, is likely very influential in cartilage tissue engineering [144]. Mechanical stimuli induce ion channel changes, such as with calcium or potassium, which could be an underlying mechanical signaling pathway [144]. The primary cilium is also suggested to contribute to mechanical signal transduction [144]. Nuclear or cytoskeleton deformation also has been shown to play a role in the cellular response to biophysical stimuli [144]. These underlying cellular changes induced by individual or combined physical stimulants could be accelerating chondrogenesis.
Models that use compression, sliding, fluidics, and HP clearly directly mimic forces found in the natural cartilage environment. However, factors such as electromagnetism, ultrasound, shaking, sliding, and microgravity are less traditional in how they constitute mechanical stimulation. Electromagnetism stimulation works by promoting cell differentiation and proliferation through the modulation of extracellular calcium entry to the cells [99,103]. Ultrasound is a mechanically vibratory form of energy that creates displacement waves within a medium, likely generating a deforming compressive environment [108]. Shaking cultures create noncontract hydrodynamic forces that impact cellular properties [131]. Microgravity conditions are created with an RCCS container in which shear force, gravity, and buoyancy are balanced to create an environment conducive to cell aggregation [133].
Mechanical stimuli for cartilage formation during embryonic and fetal development
Previous studies using models of cartilage growth and skeletal development have demonstrated that development of the joints is modulated by muscle-generated forces during the embryonic and early fetal stages. Cells residing in cartilaginous biphasic ECM can experience HP, shear, compression, and tension under loading. Those mechanical stimuli orchestrate the control of chondrogenesis of stem cells during the embryonic and early fetal stages. For example, when morphology of the joint is changing due to mechanical stimulation [145,146], articular cartilage may regenerate in some locations and calcify to form bone or degenerate in others. A better understanding of micromechanical environments during joint development at the embryonic and early fetal stages could enable development of new mechanobiological approaches to promote cartilage regeneration of stem cells. To do so, advanced computational methods, such as multiphasic models, could be the best approach that allows us to predict in situ biomechanical environment favoring cartilage formation to aid cartilage tissue engineering strategies.
Mechanotransduction of stem cells—a combined approach
Signaling pathways have extensively studied for cellular mechanotransduction under mechanical stimulation (eg, HP, shear, compression, and tension) [147]. As described previously, cells may experience a variety of mechanical stimuli simultaneously in tissues. Different mechanical stimuli may affect stem cell synergistically. For instance, HP can further enhance ECM synthesis of chondrocytes promoted by shear stress [148] while inhibiting nitric oxide release from chondrocytes induced by shear stress [149]. A combination of shear and dynamic compressive forces enhanced chondrogenic differentiation in MSCs [67,70]. Also in MSCs, shear stress combined with cyclic HP showed increased collagen II and SOX-9 [87].
Chondrocytes subjected to an electromagnetic field combined with mechanical stimulation from a ceramic hip ball had increased gene expression for the collagen II/I ratio and a more chondrogenic phenotype [150]. Therefore, combining the mechanical systems described earlier can allow us to study interplay between signaling pathways in stem cells under complex mechanical environments and optimize mechanobiological strategies for chondrogenesis of stem cells.
There is some understanding on the mechanisms underlying the effects of mechanical cues on stem cells. Mechanical forces activate signaling pathways within the stem cells, thus transducing mechanical messages into actions within the cells [151]. These actions can include responses such as but not limited to the production of growth factors or the synthesis of ECM proteins [151].
There are a number of mechanotransduction signaling pathways, and each relies on concentration changes of intracellular ions and molecules in response to a particular mechanical stimulus [151]. Tension, compression, shear stress, HP, stiffness, elasticity, and viscoelasticity are all stimuli with specific proteins that serve as transmitters in mediating the cellular response to them [147]. Downstream signaling events that are activated by these mechanical forces include YAP/TAZ, Rho/ROCK, FAK, mitogen activated protein kinase, and G protein related calcium signaling [151]. These proteins mount the cellular response to the particular mechanical stimulus [151]. A comprehensive summary of the protein response for each different mechanical stimulus covered in this review is provided in Table 3.
Signaling Pathways Triggered by Different Mechanical Stimuli
Translation of in vitro findings into clinical applications
Articular cartilage plays a crucial mechanical role in diarthrodial joints providing a low friction surface to minimize wear. The tissue further acts to absorb and distribute force within the joint. In vitro studies have demonstrated that chondrogenesis of stem cells can be promoted by mechanical stimulation. After an analysis of the different cell types used, as specified in Tables 1 and 2, it appears that the most suitable stem cell type for chondrogenesis studies are MSCs derived from adipose or bone marrow. These cells are collected from humans or animals such as rabbits or rats. However, present methods for differentiating MSCs into stable articular cartilage chondrocytes that contribute to joint regeneration have not been effective to date, despite extensive investigation.
Cell-based tissue engineering methods are being explored as a solution for the treatment of chondral defects, often an early precursor to OA. Fundamental to any successful technique will be an understanding of the loading environment that is required for the maintenance of a healthy articular cartilage phenotype. Herein may be the major obstacle from extrapolating from in vitro studies to the human condition.
Some in vivo studies have shown promise in using mechanical stimuli to better induce chondrogenesis. For example, Cui et al. used US to noninvasively condition rabbit bone-marrow derived MSCs in polyglycolic acid scaffolds that have been implanted into mice. In the US treatment group, collagen and GAG content increased significantly, and mechanical tests showed that these cells were stronger and the tissue was more cartilage like. This study suggests that US could be used to stimulate chondrogenesis in vivo [108].
Regenerative rehabilitation is an emerging field that integrates regenerative medicine with rehabilitation approaches and principles. Recent preclinical studies have demonstrated that exercise and continuous passive motion enhance cartilage repair after introducing stem cells into osteochondral defects [152,153]. However, current rehabilitative protocols for joint injury involve complex mechanical stimulation that may not provide consistent therapeutic effects as described in previous in vitro studies.
Our recent study uses an internally designed and fabricated porcine knee joint loading device which can apply controlled dynamic compression to articular cartilage by mimicking in vitro loading conditions to demonstrate similar therapeutic effects on articular cartilage as described previously [154]. Developing new rehabilitative protocols or devices to closely translate therapeutic in vitro loading conditions to in vivo settings may (1) enable us to precisely translate in vitro findings into clinical applications and (2) provide controlled mechanical stimulation for us to quantitatively optimize therapeutic effectiveness clinically. A prototype human device is under development at this time that is based on anatomic scaling of the porcine to human anatomy. Finally, it is evident that the biological and mechanical environments both play a role and interact and should be considered when devising novel strategies to possibly stimulate chondrogenesis in the human.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Funding for this work was provided by The Soffer Family Foundation and the Upwelling Foundation.