
Abstract
Stem cell replacement therapy has emerged as one of the most promising treatment options for retinal degenerative diseases, which are the main causes of irreversible vision loss. Three-dimensional (3D) retinal organoid culture is a cutting-edge technology for differentiating embryonic stem cells into retinal cells by forming a laminated retinal structure. However, 3D culture systems have strict requirements with respect to the experimental environment and culture technologies. Our study aimed to investigate the effect of retinal conditioned medium (RCM) at different developmental stages on the early differentiation of embryonic stem cells into retina in a 3D culture system. In this study, we added RCM to the 3D culture system and found that it could promote the differentiation of mouse embryonic stem cells (mESCs) into neuroretina. We further explored the possible mechanisms of RCM that regulate differentiation through proteomic analysis. RCM at different time points disclosed different protein profiles. Proteins which improved energy metabolism of mESCs might help improve the viability of embryonic bodies. We then screened out Snap25, Cntn1, Negr1, Dpysl2, Dpysl3, and Crmp1 as candidate proteins that might play roles in the differentiation and neurogenesis processes of mESCs, hoping to provide a basis for optimizing a retinal differentiation protocol from embryonic stem cells.
Introduction
Retinal degeneration diseases are the main causes of irreversible vision loss. Specifically, the degeneration of photoreceptor cells and retinal pigment epithelium (RPE) cells causes diseases like age-related macular degeneration, retinitis pigmentosa, and Stargardt's disease [1]. Degenerative damage of retinal ganglion cells (RGC) is the hallmark of diseases like glaucoma and Leber's hereditary optic neuropathy [2]. Due to the nonregeneration of retinal cells, clinical interventions only temporarily relieve symptoms and thus cannot completely reverse vision loss. Therefore, stem cell replacement therapy has become a promising approach to treating these diseases.
Pluripotent stem cells have self-renewal capacity and can differentiate into all three germ layer cells. Two-dimensional (2D) and three-dimensional (3D) culture systems have been developed for the differentiation of stem cells into retinal cells [3 –5]. 3D culture, which is also called organoid culture, has been established to cultivate laminated retinoid structure that contains various retinal neurons. To some extent, this structure can reflect an electrophysiological response under light stimulus [6,7]. Compared with 2D culture systems, 3D culture systems are more conductive to reflecting the orderly arrangement and the polarity of the cells and can form an intercellular introduction network [8]. And the process of organoids' differentiation is closer to the physiological development process of retina in vivo compared with 2D differentiation [8]. Thus, retinal organoid culture has enormous value in research on neural development, retinal disease model establishment, and organ transplantation therapy.
However, 3D culture systems have strict requirements on the experimental environment and culture technologies. The strain and state of the stem cells also have a huge impact on differentiation. Furthermore, cells in 3D culture systems are prone to general degenerative processes, so that the differentiation rate is unstable [9]. Therefore, it is still necessary to investigate a more stable and easy-operating culture system.
Retina is composed of various types of neural cells that can secrete abundant soluble factors that affect development, neurophysiology, neuroprotection, and disease states [10]. Since the development of retina follows a strict spatiotemporal sequence, the secreted soluble factors of retina also undergo a dynamic change process. Studies have reported that active substances secreted from mature cells and tissues can affect the differentiation of stem cells [2,4,11]. And the generation of particular retinal cells from stem cells can be altered by different environmental signals [12]. Retinal conditioned medium (RCM) contains various active substances secreted by retina, so we hypothesized that the RCM had the potential to promote the differentiation of stem cells into retinal cells. Therefore, the aim of this study was to investigate the effect of mouse RCM at different developmental stages on the differentiation of mouse embryonic stem cells (mESCs) into retina and to screen out the key proteins in RCM through proteomic analysis, hoping to provide a basis for optimizing retinal differentiation protocol.
Materials and Methods
Mice
C57BL/6J mice at embryonic day (E) 14, E18, and postnatal day (P) 10 (Animal Center, Southern Medical University, Guangzhou, China) were used in this study. The animal experiment was approved by Institutional Animal Ethics Committee Zhongshan Ophthalmic Center (ethical approval No. 2020-043).
Collection of RCM
Intact retinas from mice at E14, E18, and P10 were isolated following the protocol optimized by Suphansa and Johnson et al. [13,14]. Briefly, eyes were immersed in ice-cold Hank's balanced salt solution containing penicillin (100 U/mL) and streptomycin (100 μg/mL). Using very fine forceps, anterior segments, lens, and vitreous body were gently removed, and RPE was peeled away. The neuroretina was dissected and transferred into retina culture medium. The medium composition was in Supplementary Data. The cultures were maintained in cell incubators at 37°C and 5% carbon dioxide. The media was changed at 6 h and 24 h, and RCM was collected.
mESC culture and differentiation
The mESC line was derived from the C57BL/6J strain and was generously provided by Prof. Xiang Peng from Zhongshan Medical School, Sun Yat-sen University. The ESC line was maintained on feeder mouse embryonic fibroblast cells as previously described [5,15]. Retinal differentiation was optimized on previously published protocols [5,16]. In brief, on the day of differentiation (D0), mESCs were dissociated from feeders and plated into 96-well low-attachment plate (3000 cells per 100 μL per well). On D1, 2% growth factor reduced Matrigel (BD Bioscience, USA) was added. On D7, embryonic bodies (EBs) were transferred to low-binding plates (Corning, USA). And the medium was changed every 2–3 days; 1mM taurine (Sigma, USA) and 0.5 μM all-trans retinoid acid (Sigma, USA) were added from D10 to D14. A measure of 1 mM taurine was added into the medium from D14 onward. Media composition was in Supplementary Material.
On D0 and D7, retina culture medium (negative control) and RCM at different time points (E14, E18, and P10) were added into the culture medium at the concentration of 1:4, respectively.
Immunofluorescent staining
Cells or EBs at different time points were fixed in 4% paraformaldehyde for 30 min and then washed in phosphate-buffered saline (PBS). For EBs, dehydrated with 15% sucrose overnight at 4°C and embedded up to 10 EBs in a cryomold with optimal cutting temperature compound and frozen at −20°C. Made 10 μm cryosections using a cryostat (Leica, Germany). Dried the slides, washed with PBS thrice, and incubated in blocking solution (5% bovine serum albumin, 0.5% Triton X-100) for 1 h at room temperature. Then the slides were incubated with primary antibody overnight at 4°C (Supplementary Table S1). Washed with PBST thrice, the specimens were incubated with Alexa Fluor 555 and 488 secondary antibodies (Invitrogen, USA) for 2 h at room temperature. After washing with PBST for several times, the specimens were incubated with DAPI for 5 min. Washed with PBST, the slides were mounted with antifade mounting medium (CST, USA).
Image acquisition and analysis
Live images in bright field were captured using inverted microscope (Olympus, Japan). The surface area and the neuroepithelium area of EBs were measured using ImageJ software (NIH, USA). At least eight EBs were analyzed for each condition and for each biological replicate. Three biological replicates were included. Averages were plotted using GraphPad Prism software (GraphPad Prism Software Inc., USA). Fluorescent images were captured using an upright fluorescent microscope (Zeiss, Germany). Final images were adjusted for brightness and contrast in Photoshop software (Adobe Systems Inc., CA, USA).
Quantitative real-time polymerase chain reaction
RNA was extracted from EBs at D7 and D10 using the EZ-press RNA Purification Kit (EZBioscience, USA). One thousand nanogram of RNA was converted into cDNA using PrimeScript™ RT Reagent Kit (Takara, Japan). Quantitative polymerase chain reaction (qPCR) was performed using TB Green™ Premix Ex Taq Ⅱ Kit (Takara). All reaction systems were added following the manufacturer's protocols. Triplicated samples were analyzed by quantitative real-time PCR (LightCycler480Ⅱ, Roche, Switzerland). The PCR program was set as denaturation at 95°C for 10 min, followed by 40 cycles of denaturation at 94°C for 30 s, annealing at 60°C for 30 s, and extension at 72°C for 1 min. The relative expression of mRNA was calculated using the 2−ΔΔCt method. The primer sequences are listed in Supplementary Table S2.
Label-free proteomic analysis
The RCM samples for protein extraction were obtained by mixing RCM taken in three separate batches. Proteins were extracted from E14, E18, and P10 RCM samples, respectively, and were used for mass spectrometry-based label-free proteomics (performed by Fitgene Biological Technology, Guangzhou, China). Briefly, after measuring the protein concentration, the proteins were incubated with trypsin and transferred into peptides. The peptides were subjected to liquid chromatography tandem mass spectrometry analysis. The raw data were identified and quantified using the MaxQuant software based on the UniProt protein database. Significant proteins (peptides ≥2, coefficient of variation (CV) <0.5, P < 0.05) were screened out. Differential proteins were defined as average ratio (AVG) ≥1.2 or AVG ≤0.8. Bioinformatic analyses such as clustering analysis (
Statistical analysis
Statistical analysis was performed using the GraphPad Prism software. Data were described as mean ± standard deviation. Statistical significance was tested by One-way analysis of variance. P < 0.05 was considered statistically significant.
Results
Characterization of mESCs from C57BL/6J mice
The mESCs were maintained on the feeder layer cells in an undifferentiated state. The mESC clones showed a suborbicular shape with bright edges (Supplementary Fig. S1A). We evaluated the multipotential capacity of the mESC line from C57BL/6J mice evaluated by immunostaining using pluripotency markers. The expression of Nanog and OCT3/4 was detected positive in mESC clones (Supplementary Fig. S1B, C), showing that the mESCs were in a good state and retained pluripotency.
Identification of retinal structures during early differentiation
The morphological changes during the differentiation process were consistent with those demonstrated in previous studies. In the 3D culture system, on D1, the mESCs aggregated into spherical EBs (Supplementary Fig. S2A). On D4, the EBs formed continuous transparent neuroepithelial structures (Supplementary Fig. S2B). On D7, optic-vesicle-like (OV-like) structures were observed (Supplementary Fig. S2C). By D10, the size of the EBs had enlarged, and a few optic-cup-like structures had appeared (Supplementary Fig. S2D).
The EBs were fixed for immunofluorescence staining to detect the expression of retinal marker genes. On D7, EBs expressed early RPC markers such as Rx and MITF. On D10, early neuroretinal markers such as Pax6 and CHX10 were positive (Supplementary Fig. S3).
RCM promoted the early formation of neuroepithelium
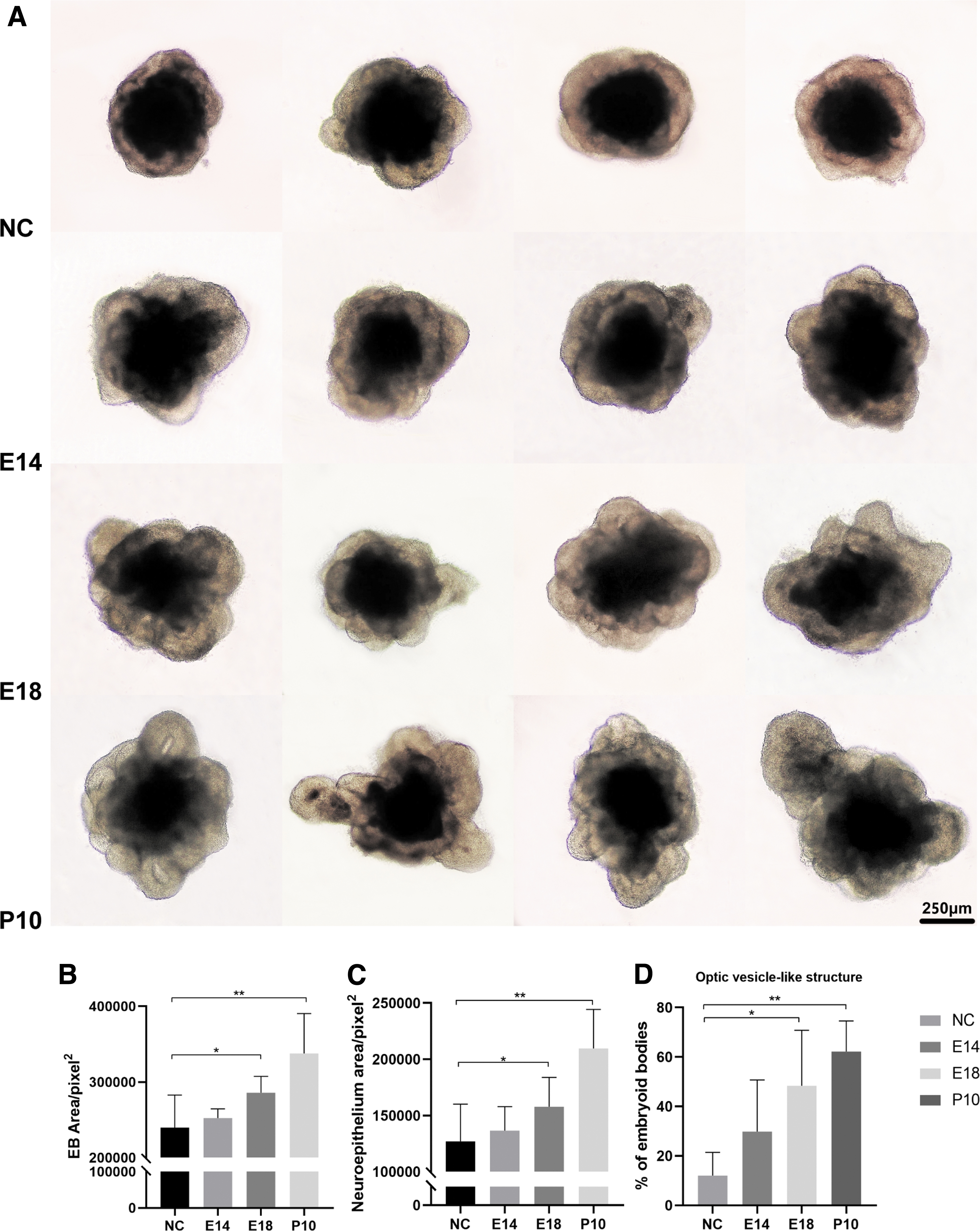
On D0, adding RCM into the culture system at a concentration of 1:4, morphological observation was performed on D7 (Fig. 1A). To objectively evaluate the observation, we quantified the area of EBs, the neuroepithelium area of EBs, and the percentage of OV-like structures. We found that RCM at E14, E18, and P10 significantly increased the area of EBs and promoted the formation of neuroepithelium and OV-like structures (Fig. 1B–D, P < 0.05). Adding RCM, the percentage of OV-like structures significantly improved from around 40% to above 60%; this percentage could be even increased to around 80% when RCM at P10 was added. We continued to add RCM on D7 and observed the morphological changes on D10 (Fig. 2A). Compared with the control group, RCM contributed to maintaining the neuroepithelium and OV-like structure, especially RCM at E18 and P10 (Fig. 2B–D, P < 0.05).

Morphological observations of EBs on D7.
Morphological observations of EBs on D10.
RCM promoted the early differentiation of mESCs into retinal cells
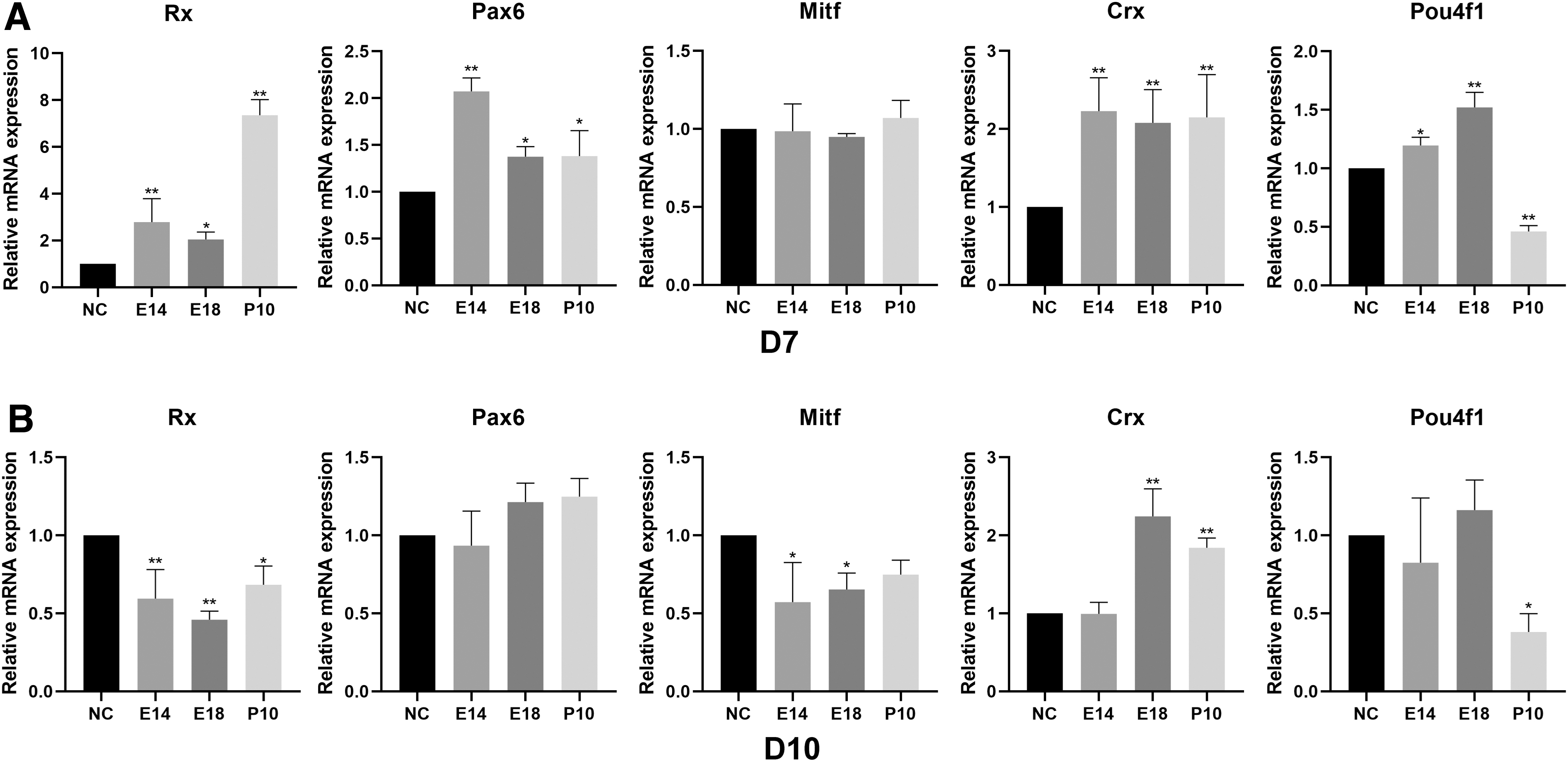
We collected EBs on D7 and D10 to extract RNA and detected the expression levels of various retinal marker genes (Rx, Pax6, Mitf, Crx, and Pou4f1) through qPCR. On D7, the expression level of Rx increased significantly in RCM groups, indicating that the addition of RCM had promoted the differentiation of mESCs into retinal progenitor cells (RPCs). The expression level of Pax6 in the RCM group slightly increased, whereas the Mitf expression level did not change significantly, indicating that to some extent, RCM helped mESCs differentiate into neural cell fate rather than epithelial cell fate. Furthermore, RCM might have benefits on the differentiation into photoreceptor precursors because Crx levels in RCM groups had significantly improved. We also found that the expression of Pou4f1 increased after adding RCM at E18, indicating that RCM's effect on differentiation direction might vary at different time points (Fig. 3A).
Relative expression of retinal marker genes.
On D10, the expression of Rx and Mitf in the RCM groups decreased, and the expression of Crx in E18 and P10 RCM groups increased. Compared with those in the control group, the RPC marker genes in the RCM group were downregulated, indicating that RPCs might have further differentiated. Adding E18 and P10 RCM, the differentiation of RPCs toward photoreceptor precursor fate was more obvious (Fig. 3B).
Proteomic profile of RCM
To further investigate the mechanism of RCM in its effect on differentiation, we performed proteomic analysis. We identified 198 proteins, mainly small molecular weight, acidic hydrophilic proteins (Fig. 4A–C). We further screened out 123 reliable proteins (number of peptides ≥2, CV <0.5, P < 0.05), in which 15 proteins were common to the three groups of RCMs (Fig. 4D). The identified common proteins belonged to five functional categories: vesicle-mediated transport, cellular response to chemical stress, regulation of supramolecular fiber organization, organelle localization, and hemostasis (Fig. 4E).

Characteristics of identified proteins.
Since the functional analysis of the common proteins revealed no beneficial effect on differentiation, to further find proteins that might contribute to the differentiation or neurogenesis of stem cells, we screened 105 differential proteins using pairwise comparison between groups. The heat map showed the abundance of the differential proteins (Fig. 5A). We observed that P10 RCM disclosed a different profile and had a much richer variety of proteins than E14 and E18 RCM. The differentially expressed proteins were presented as volcano plots (Fig. 5B–D).
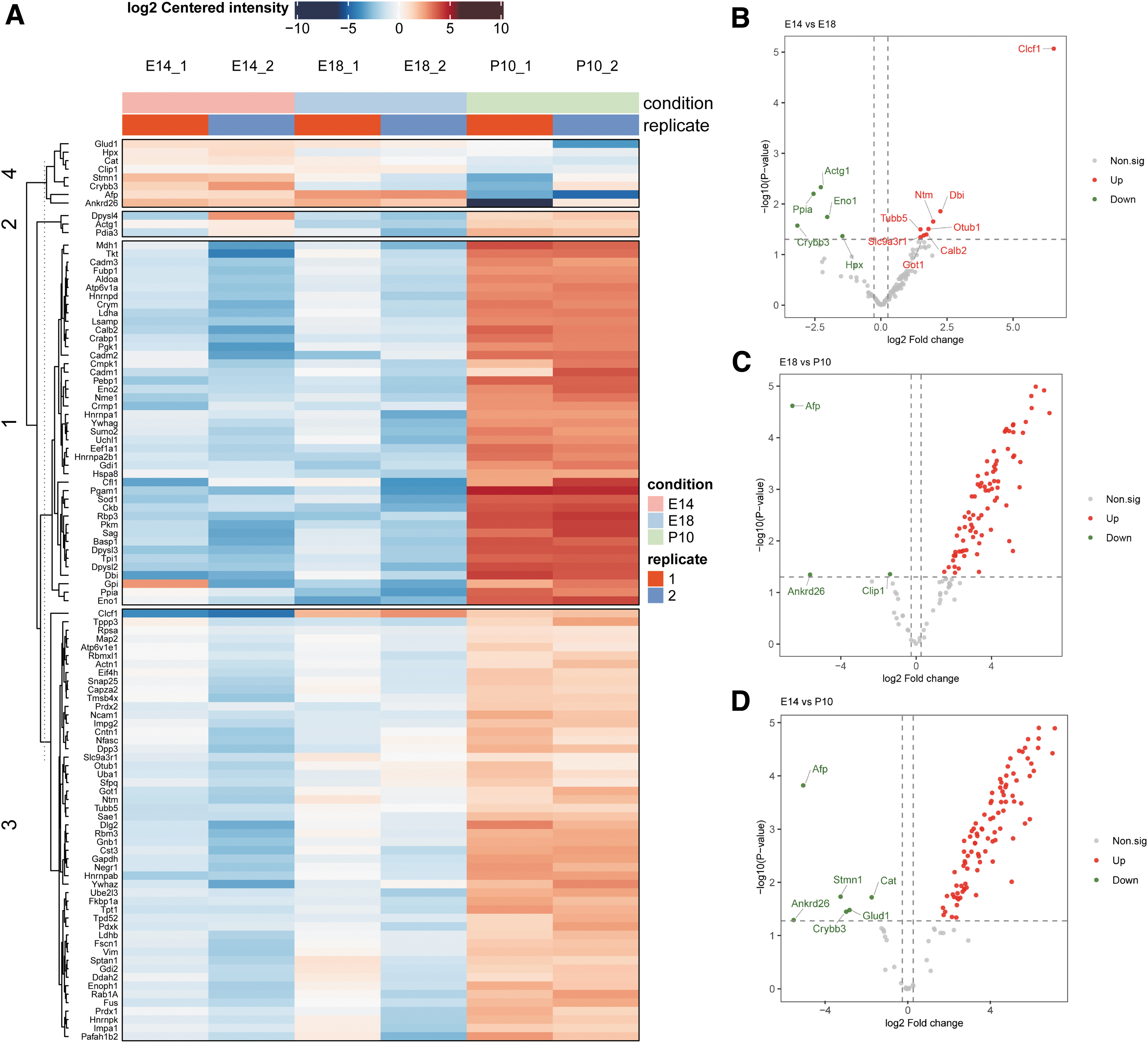
Expression of differential proteins.
GO Analysis and protein-protein interaction analysis
Among the differential proteins, we identified the GO items of 103 proteins. Biological process annotation revealed that the proteins were mainly associated with metabolic processes and response to stimuli. Cell composition annotation indicated that the proteins were mainly involved in cytosol, membrane structure, nucleus, and cell protrusion structure. Molecular function annotation showed that the proteins were related to binding activity, especially protein-binding activity and ion-binding activity (Fig. 6A). Pathway enrichment analysis showed that the proteins were mainly involved in carbon metabolism, regulation of neuron projection development, and aldehyde biosynthetic processes (Fig. 6B).

Bioinformatic analysis of differential proteins.
There were 8 subordinate terms in the term of neuron projection development, and they involved 17 proteins (listed in Supplementary Table S3). We carried out protein–protein analysis and grouped the network into five clusters using the Markov Cluster Algorithm (inflation parameter, 3). Proteins in two clusters had stronger association inside the cluster, and the two clusters were connected to each other. In the red and yellow clusters, the core proteins included synaptosome-associated protein 25 (Snap25), Cntn1 (Contactin/F3/F11), neuronal growth regulator 1 (Negr1), Dpysl2, Dpysl3, and collapsing response mediator proteins (Crmp)1 (Fig. 6C).
Discussion
Because of the indefinite self-renewal of ESCs, they have the possibility of providing an unlimited source of retinal cells, which makes cell replacement therapy a promising radical treatment for retinal degeneration disease. Methods for ESC differentiation are varied, and stem cells can be differentiated into RPE cells, RGCs, and photoreceptors. However, the cell function is immature, and the induction rates are inefficient and highly variable [4,17,18]. By adding Matrigel, a material enriched with extracellular matrix proteins, the EBs can spontaneously differentiate into retinal structures. This process generates OV-like structure that simulates the retinal development process in vivo [19,20]. However, the undefined nature and lot-to-lot variations of Matrigel have become a bottleneck for the stable reproducibility of organoids in 3D culture systems [21]. Thus, to achieve a robust and reproducible 3D culture system, it is still necessary to delve into the molecular regulation mechanism of organoid genesis and optimize the culture protocol.
It is widely known that the microenvironment can affect the differentiation of stem cells. The differentiation direction of ESCs can be manipulated by changing the extracellular matrix environment or by adding different small molecules [22]. Related studies have reported some effects of different conditioned mediums on cells. For example, RCM can promote the differentiation of RPCs into retinal neural cell fate [23], RGC conditioned medium promotes the differentiation of RPCs into RGCs, and Müller cell conditioned medium improves the survival of mESC-derived RGCs [2,24]. Thus, the effect of microenvironment on stem cells can be a good entry point for studying the molecular regulation mechanism of organoid genesis.
Retina can secrete various extracellular soluble factors that are active in multiple functions [10]. It has been reported that RCM can promote the expression of tight junction markers of RPE cells and induce the differentiation of lens epithelium cells [25,26]. However, there is a lack of research on the effect of RCM on the differentiation of ESCs into the retina. In our study, we collected RCM at E14 (the proliferation peak of cone precursors in mice), E18 (the proliferation peak of rod precursors in mice), and P10 (the period of increased expression of mature proteins of photoreceptors) [27] and added them into the early 3D culture system.
Due to the development of retinal cells in a strictly spatiotemporal manner, we speculated that RCM could provide a microenvironment closer to the physiological state for EBs, so that the viability and the differentiation efficiency would be improved. And our results showed that RCM improved the formation of neuroepithelial structures and increased the proportion of OV-like structures. The PCR results also confirmed that RCM had the potential to induce mESCs to differentiate into neuroretina, and the effect of RCM at different periods could be slightly different. Hence, the differentiation direction of RCM at different periods is worthy of more in-depth study.
To further investigate the possible active substance in RCM which promotes the activity and differentiation of mESCs, we performed a proteomic analysis. Through hierarchical cluster analysis and pairwise comparison, it was found that the protein abundance of the P10 RCM was significantly higher compared with the E14 and E18 RCM. This is because the development of retina follows a strict spatiotemporal paradigm, and at P10, all types of retinal cells are almost mature. Thus, the P10 RCM had more complex components and showed a more obvious effect on the mESCs. Since the RCM had a promoting effect, although to different degrees, on the formation of the neuroepithelial structure at three time points, we analyzed the common proteins at each time point. However, the analysis of these proteins showed functions related to basic cellular activities, rather than activities related to neural differentiation. We then performed a comparative proteome analysis to elucidate the common effects of different RCMs on stem cells. Differential proteins were then screened and analyzed.
Through pathway enrichment analysis of the differential proteins, we found that the proteins were closely related to carbon metabolism. ESCs self-renew rapidly with a shortened G1 cell cycle phase. Carbon metabolism, especially glycolysis, allows for the quick generation of ATP, which fuels the proliferation of ESCs and RPCs [28]. Carbon metabolism is also indispensable for cell differentiation and biosynthesis during the development of retina [29]. In addition to growth factors and the extracellular matrix, carbon metabolic pathways can contribute to establishing a balance between the self-renewal and differentiation of ESCs [30]. Thus, the promoting effect of RCM on the viability and neurogenesis of EBs might be due to the improved energy metabolism of mESCs. The cross-talks between carbon metabolism and signaling pathways thus deserve to be further defined.
Proteins related to the regulation of neuron projection development were included in the second most enriched pathway. We screened out six core proteins that might play important roles in the differentiation and neurogenesis processes of mESCs. Snap25 is involved in numerous cellular functions, such as neurotransmission, vesicle priming, axon outgrowth, and photoreceptor differentiation [31]. In the developing retina, a high expression level of Snap25 has been identified in photoreceptors [32]. A study also points out that different expression levels of Snap25 play a crucial role in the cell fate determination of photoreceptors; a relatively low level of Snap25 seems to favor the differentiation of cones [33]. Although the spatiotemporal expression of Snap25 in retina has not been fully clarified, a high level of Snap25 in the developing retina is essential to the correct differentiation of photoreceptors. The effect of Snap25 on retinal development and stem cell differentiation deserves further study.
Crmp1, Dpysl2 (Crmp2), and Dpysl3 (Crmp4) are homologous proteins of collapsing response mediator proteins (CRMP). CRMP are highly expressed during brain and retina development and downregulated in adults [34]. The functions of CRMP include neuronal polarity, axonal outgrowth, and proper positioning of neurons [35]. Cntn1 is a novel adhesion protein that plays crucial roles in neuronal migration and retinal axon regeneration [36,37]. Negr1 is a cell adhesion molecule that is reported to enhance axonal outgrowth from neural stem cells [38]. The Cntn1, Negr1, Dpysl2, Dpysl3, and Crmp1 are reported to have relationship with neural regeneration, neuronal migration, and neural differentiation. These proteins might have relationship with retinal development, but their specific effects and mechanisms need to be further verified.
Conclusion
Our study found that RCM could promote the differentiation of mESC into the neuroretina and that RCM at different time points might have different differentiation effects. Improving the energy metabolism of mESCs might help maintain the viability of EBs. There were some proteins in the RCM that influenced the process of neurogenesis, but further studies are needed to verify these effects and mechanisms and to further optimize the differentiation plan. Nevertheless, our study sheds some light on how a stable 3D culture system can be improved at the early stage.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China to Y.L. (Grant No. 81770971) and the Natural Science Foundation of Guangdong Province, China to Y.L. (Grant No. 2017A030313787).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.