
Abstract
Neuroinflammation is one of the typical events in multiple neurodegenerative diseases, whereas microglia are the critical participants in the pathogenesis of neuroinflammation. Several studies suggest that neural stem cells (NSCs) present immunomodulatory benefits due to their paracrine products, which contain mounting trophic factors. In the current study, the anti-inflammatory effects of NSC secretome (NSC-S) on lipopolysaccharide (LPS)-induced neuroinflammatory models were evaluated in vivo and the underlying mechanism was further investigated in vitro. It was revealed that NSC-S significantly attenuated the severity of LPS-induced behavior disorders and inflammatory response in mice. In vitro studies found that NSC-S significantly promoted the polarization of microglia from proinflammatory M1 to anti-inflammatory M2 phenotype, and reduced the production of proinflammatory cytokines, whereas elevated anti-inflammatory cytokines in BV2 cells. NSC-S promoted peroxisome proliferator-activated receptor gamma (PPAR-γ) pathway activation. However, these effects of NSC-S were abrogated by PPAR-γ inhibitor GW9662. Notably, the fatty acid-binding protein 5 (FABP5) in NSC-S may mediate PPAR-γ activation and inflammation remission. In summary, NSC-S promotes the regression of LPS-induced microglia-mediated inflammation through the PPAR-γ pathway. This function might be achieved through FABP5.
Introduction
Neuroinflammation is a reaction of the central nervous system in response to endogenous or exogenous stimulation. It involves the activation of microglia, astrocytes, and other immune cells as well as the consequent changes in the levels of inflammatory mediators and cytokines, leading to degeneration or loss of neurons [1]. It has been reported that neuroinflammation is involved in triggering a variety of neurodegenerative diseases, including multiple sclerosis, Alzheimer's disease, Parkinson's, and amyotrophic lateral sclerosis [2 –5]. Hence, antineuroinflammation is a key factor in neurodegenerative disease therapy.
Microglia are innate immune cells in the brain with the function of proliferation, migration, and phagocytosis and are important participants in the process of neuroinflammation [6,7]. Under normal physiological conditions, acting as the central nervous system monitor, microglia sense the changes of the surrounding environment with its special movement process to maintain the stability of the immune system [8]. In response to external stimuli or internal environmental changes, microglia are activated into different phenotypes: proinflammatory M1 and anti-inflammatory M2, showing inflammatory or protective effects by secreting different mediators [9]. The M1 phenotype mainly secretes inflammatory cytokines and mediators, such as induced nitric oxide synthase (iNOS), interleukin-6 (IL-6), interleukin-1β (IL-β), tumor necrosis factor α (TNF-α), and nitric oxide (NO), leading to an inflammatory cascade and neuronal degeneration and loss.
In contrast, the M2 phenotype mainly secretes anti-inflammatory factors, such as interleukin-10 (IL-10) and arginase-1 (Arg-1), to promote the regression of neuroinflammation and remove neuronal debris, resulting in neuronal damage repair and neuroprotection [10,11]. Research has shown that suppressing M1 and promoting M2 phenotype of microglia contribute to decreased neuroinflammation [12]. Therefore, the mediation targeting microglia phenotype may be an effective strategy for the treatment of neuroinflammation-related neurodegenerative diseases.
Stem cell therapies have received increasing attention in recent years. Neural stem cells (NSCs) are capable of proliferating, self-renewing, and differentiating into neurons, astrocytes, and oligodendrocytes. Studies have shown that transplanted NSCs are involved in the process of tissue repair and rescue of neurodegeneration in various central nervous system diseases and injuries through cell replacement, immunosuppression, neuronutrition, and other means [13 –15]. Although stem cell therapy has great potentials, the use of stem cell derivatives as an alternative therapy appears to be a more convenient and safe approach to treatment [16,17]. NSC secretome (NSC-S) derivatives have been reported to have distinct neuroprotective and immunosuppressive effects [18 –20]. Our previous studies have shown that NSC-S can ameliorate peripheral nerve injury in rats [21], which supports the therapeutic effects of NSC-S.
On this background, this study aimed at investigating the role of NSC-S in the treatment of neuroinflammation, further exploring the effect of NSC-S on microglia phenotype and the potential mechanism involved.
Materials and Methods
Cell culture
Cell culture was carried out as outlined in our previous studies [22]. NSCs were differentiated from human embryonic stem cells and cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 (Gibco, USA) containing 1% penicillin–streptomycin, 20 ng/mL basic fibroblast growth factor (PeproTech, USA), and 20 ng/mL epidermal growth factor (PeproTech). BV2 and N2a were purchased from the Cell Resource Center, Peking Union Medical College (Headquarter of National Infrastructure of Cell Line Resource, NSTI) and were cultured in DMEM (Gibco) with 10% fetal bovine serum (Gibco).
Since previous studies showed that the morphology did not change and the biological characteristics were relatively stable in P2-P18 NSCs [22], this study collected NSC-S from P7-P15 NSCs. At 80%–90% confluence, P7-P15 NSCs were washed with serum-free DMEM five times and maintained in serum-free DMEM for 6 h. NSC-conditioned medium was collected and centrifuged at 2000 rpm for 20 min to remove cell debris, then, the collected supernatant was filtered with the 0.22 μm membrane to create NSC-S, and stored at −80°C for the next experiments. For animal experiment, 50 mL NSC-conditioned medium was concentrated into dry powder with a lyophilizer (Labconco FreeZone, England), and then dissolved with 5 mL sterile ultrapure water to create 10 × NSC-S.
Animal model establishment and treatment
Male C57BL/6 mice were purchased from the Laboratory Animal Center of Jiangsu University (Zhenjiang, China). They were raised in plastic cages under a 12-h light–12-h dark cycle with food and water ad libitum. All animal experiments were approved by the Institutional Animal Care and Use Committee of Jiangsu University (Permit Number: SCXK 2018-0012). As described previously [23], neuroinflammation model was induced by intraperitoneal injection of lipopolysaccharide (LPS, 5 mg/kg, from Escherichia coli O55:B5; Sigma-Aldrich, Germany). The animals were divided into four groups randomly: DMEM-treated group, LPS-induced group, LPS-induced treated with NSC-S group, and LPS-induced treated with dexamethasone (DEX; Beyotime, China) group. There were eight mice in every group. After LPS induction for 2 h, 50 μL 10 × NSC-S or DEX (5 mg/kg) was administered through the caudal vein. Fifty microliters of 10 × DMEM was used as control. Subsequent animal experiments were conducted 24 h later.
Tail suspension with head swing test
Tail suspension with head swing test was performed using previous method [24] to evaluate the effect of 10 × NSC-S on the reaction capacity of mice. The tails of mice were briefly fixed with their heads pointing down, and the number of head swings in 1 min were recorded.
Open field test
Open field test (OFT) was performed to evaluate spontaneous locomotor activity of mice [25]. Mice were placed in a relatively quiet and low-light box. The bottom area of the box was cleaned up with ethyl alcohol before the test to avoid the effect of previous tests. The experimental mice were placed in the open field for 30 min. The trajectory, total traveled distance, and time in the center square within the 30 min were recorded using the Ethovision 8.5 system (Noldus, the Netherlands).
Hematoxylin and Eosin staining
Coronal section of brain tissue was performed to obtain a brain section of 5 μm, and then subjected to Hematoxylin and Eosin (H&E) staining. Briefly, sections were dried and cleared in xylene, dehydrated in an ethanol gradient, and then stained with H&E at room temperature, respectively. Finally, the sections were mounted by neutral gum.
Quantitative real-time polymerase chain reaction
Total RNA was extracted using RNA-easy™ isolation reagent (Vazyme, China) according to the manufacturer's instructions. The complementary DNA (cDNA) was prepared by using the Quantscript RT Kit (Vazyme). The cDNA was diluted at a ratio of 1:5 and mixed with the SuperReal PreMix Plus Kit (SYBR Green; Vazyme) for quantitative real-time polymerase chain reaction (qRT-PCR). The results were analyzed by the 2−ΔΔCt method. The primer pair sequences for gene amplification are listed in Table 1.
Primer Sequences Used for Quantitative Real-Time Polymerase Chain Reaction
Arg-1, arginase-1; CSF3, colony-stimulating factor 3; IL-1β, interleukin 1β; IL-10, interleukin 10; IL-6, interleukin 6; iNOS, inducible nitric oxide synthase; LCN2, lipocalin 2; TNF-α, tumor necrosis factor alpha.
Western blot
Total protein was extracted using RIPA buffer, which contains the protease and phosphatase inhibitors (Beyotime). The western blot was performed as previously described [26]. The primary antibodies used were anti-β-actin (Cell Signaling Technology, USA), anti-Histone3 (BOSTER, China), anti-GAPDH (Mutisciences, Hangzhou), anti-IL-6 (Wanleibio, China), anti-TNF-α (Wanleibio), anti-iNOS (Wanleibio), anti-Arg-1 (Wanleibio), anti-nuclear factor kappa B inhibitor alpha (IKBα) (Wanleibio), anti-phosphorylation-IKBα (p-IKBα) (Wanleibio), anti-nuclear factor kappa B (NF-κB) (BOSTER), anti-IL-1β (BOSTER), anti-IL-10 (BOSTER), and anti-peroxisome proliferator-activated receptor gamma (PPAR-γ) (BOSTER). The protein bands were detected by the High-Sensitivity ECL Chemiluminescence Detection Kit (Vazyme), and gray analysis of the immunoblots were performed by Lane 1 after image exposure.
Peripheral blood neutrophil cell count
The mice were anesthetized and immobilized, and the chest was exposed to collect blood from the heart using a vacuum needle. Blood smear was performed immediately after blood collection. Wright-Giemsa (Baso Diagnostics Inc., Zhuhai, China) staining was conducted according to the manufacturer's instructions, and the ratio of neutrophils in 100 white blood cells were counted under an ordinary light microscope at magnification of 1000 times.
Immunofluorescence staining
BV2 cells were stimulated with LPS (100 ng/mL) and NSC-S for 24 h. The immunofluorescence staining of cultured cells was carried out according to standard protocols. Briefly, cells were fixed with 4% paraformaldehyde for 30 min and permeabilized with phosphate buffered solution (PBS) containing 0.1% Triton X-100 for 20 min. For CD86, the cell membranes did not need to be permeabilized, and then the cells were blocked with 5% bovine serum albumin for 1 h at room temperature. This was followed by incubation with the immunofluorescence primary antibodies, including anti-CD86 (BOSTER), anti-CD206 (ProteinTech, USA), and anti-ionized calcium-binding adapter molecule 1 (Iba-1) (Wanleibio) at 4°C overnight, and staining with the corresponding secondary antibodies for 1 h at room temperature. The sections or cells were counterstained with 4,6-diamidino-2-phenylindole for 5 min and mounted with antifluorescence quenching agent. Images were acquired using a Nikon fluorescence microscope or confocal microscope.
Nissl staining
With reference to the instructions of the Nissl Staining Kit (Solarbio, Beijing), the brain sections were immersed in Cresyl Violet Stain at 56°C for 1 h and then differentiated for 1 min using the Nissl differentiation solution. The sections were quickly dehydrated with anhydrous ethanol, cleared with xylene, and finally sealed with neutral gum. A scanning machine was used to observe sections.
Griess test
Production of NO was detected by the Griess Kit (Nanjing Jiancheng, China). In short, the cell supernatants were collected after 24 h of treatment with LPS in the presence or absence of NSC-S. One hundred microliters of the supernatant of each well was collected, and Griess reagents were added successively according to the protocol. The absorbance was measured at 550 nm wavelength using a microplate reader. Sodium nitrite solution with a concentration of 20 μmol/mL was used as a standard in the assay.
Flow cytometry analysis
The Reactive Oxygen Species (ROS) Assay Kit (Beyotime) was used for ROS production in BV2 cells. Briefly, cells were stimulated by LPS in the presence or absence of NSC-S for 4 h, and then incubated with diluted 2,7-dichlorodi-hydrofluorescein diacetate (DCFH-DA) at 37°C for 20 min. Finally, cells were resuspended in PBS for flow cytometry.
NanoLiquid chromatography–tandem mass spectrometry analysis
After concentrates of NSC-S were obtained, we performed protein extraction, quantification, reduced alkylation, enzymolysis, and peptide analysis. For each sample, 2 μg of total peptides were separated and analyzed with a nano-UPLC (EASY-nLC1200) coupled to a Q Exactive HFX Orbitrap instrument (Thermo Fisher Scientific) with a nano-electrospray ion source. Vendor's raw mass spectrometry files were processed using Proteome Discoverer (PD) software (Version 2.4.0.305) and the built-in Sequest Hyper-Threading search engine.
Statistical analysis
All quantitative results are presented as the mean ± standard error of the mean. One-way analysis of variance was conducted to determine statistical significance among multiple experimental groups with GraphPad Prism software version 6.01. P values <0.05 were considered statistically significant.
Results
NSC-S improves behavioral disorders in LPS-induced neuroinflammation in mice
After LPS injection, the number of mice head swing to the left or right was decreased compared with the 10 × DMEM group, while 10 × NSC-S treatment significantly increased head swing frequency (Fig. 1A). Mice were also subjected to OFT to evaluate their excitability and exploration behavior. The results showed that LPS injection caused a reduction in total distance of movement in the open field and stay time spent in the central area of the field (center time). Following 10 × NSC-S treatment, the total distance covered and center time increased significantly (Fig. 1B–D).
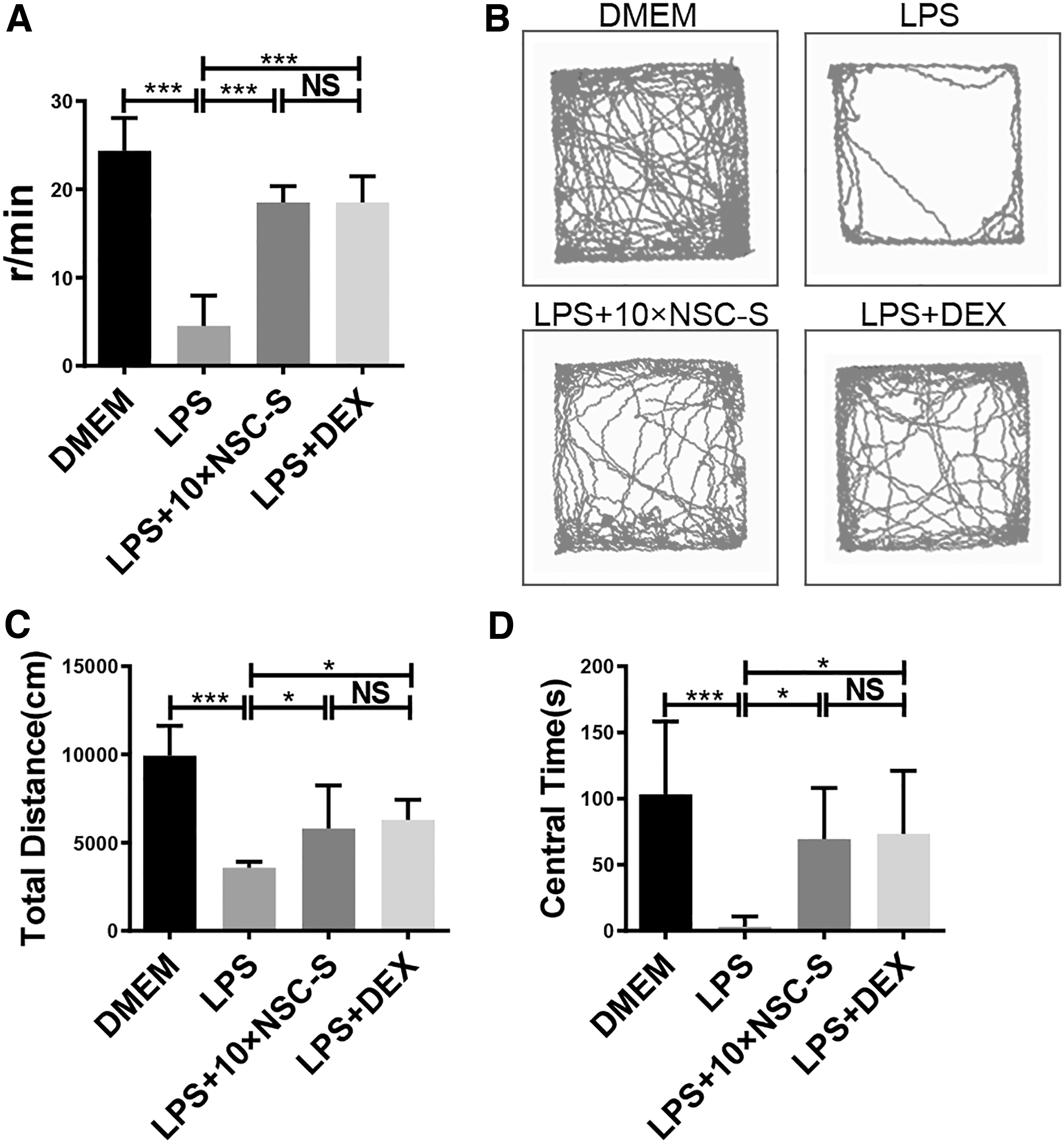
Effects of NSC-S on LPS-induced behavioral disorders.
NSC-S suppresses systemic and neuroinflammation in mice
Peripheral blood neutrophil count results showed that 10 × NSC-S reduced LPS-induced neutrophil elevation (Fig. 2A). In the LPS-induced group, mice exhibited white secretions around their eyes. However, 10 × NSC-S treatment remarkedly improved this symptom (Fig. 2B). H&E staining revealed that LPS caused focal gliosis in the cortex, but 10 × NSC-S administration significantly abolished the gliosis in the brain (Fig. 2C). Nissl's staining showed that the Nissl substance in the cortex and hippocampus from the LPS-induced group exhibited significant loss compared with those from the 10 × DMEM group. While 10 × NSC-S treatment increased Nissl substance in neurons. The shrunken and deeply stained neurons induced by LPS were also greatly reduced in the cortex and hippocampus by 10 × NSC-S treatment (Fig. 2D).
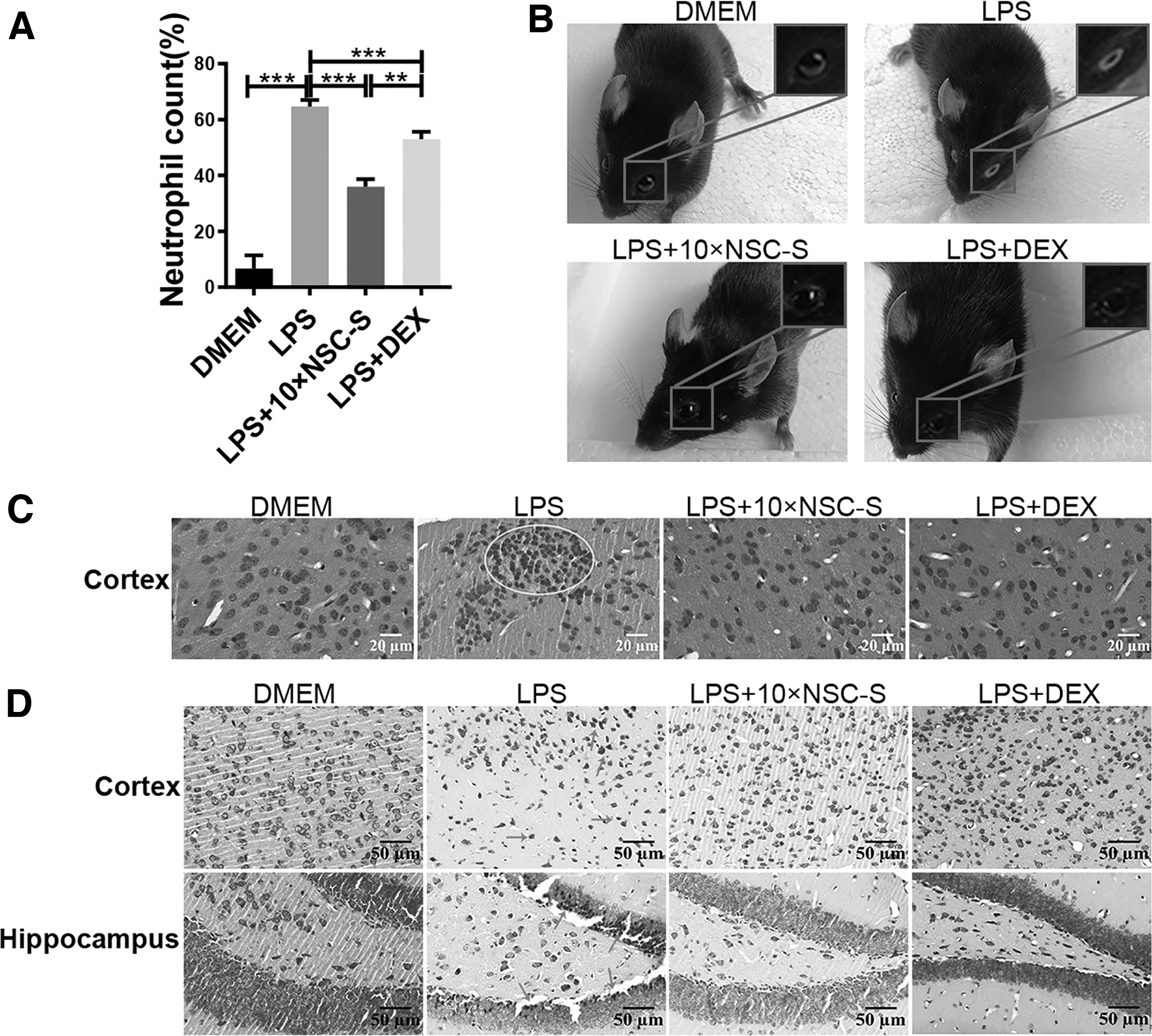
Effects of NSC-S on peripheral blood neutrophils and inflammatory infiltration of the brain.
Moreover, it was observed that LPS induced the upregulation of proinflammatory enzyme and cytokine (iNOS, IL-1β, IL-6, TNF-α) gene and protein levels in the cortex and hippocampus. These proinflammatory enzymes and cytokines were dramatically restored by 10 × NSC-S, while increasing the expression of the anti-inflammatory cytokines, Arg-1 and IL-10 (Fig. 3A–E).
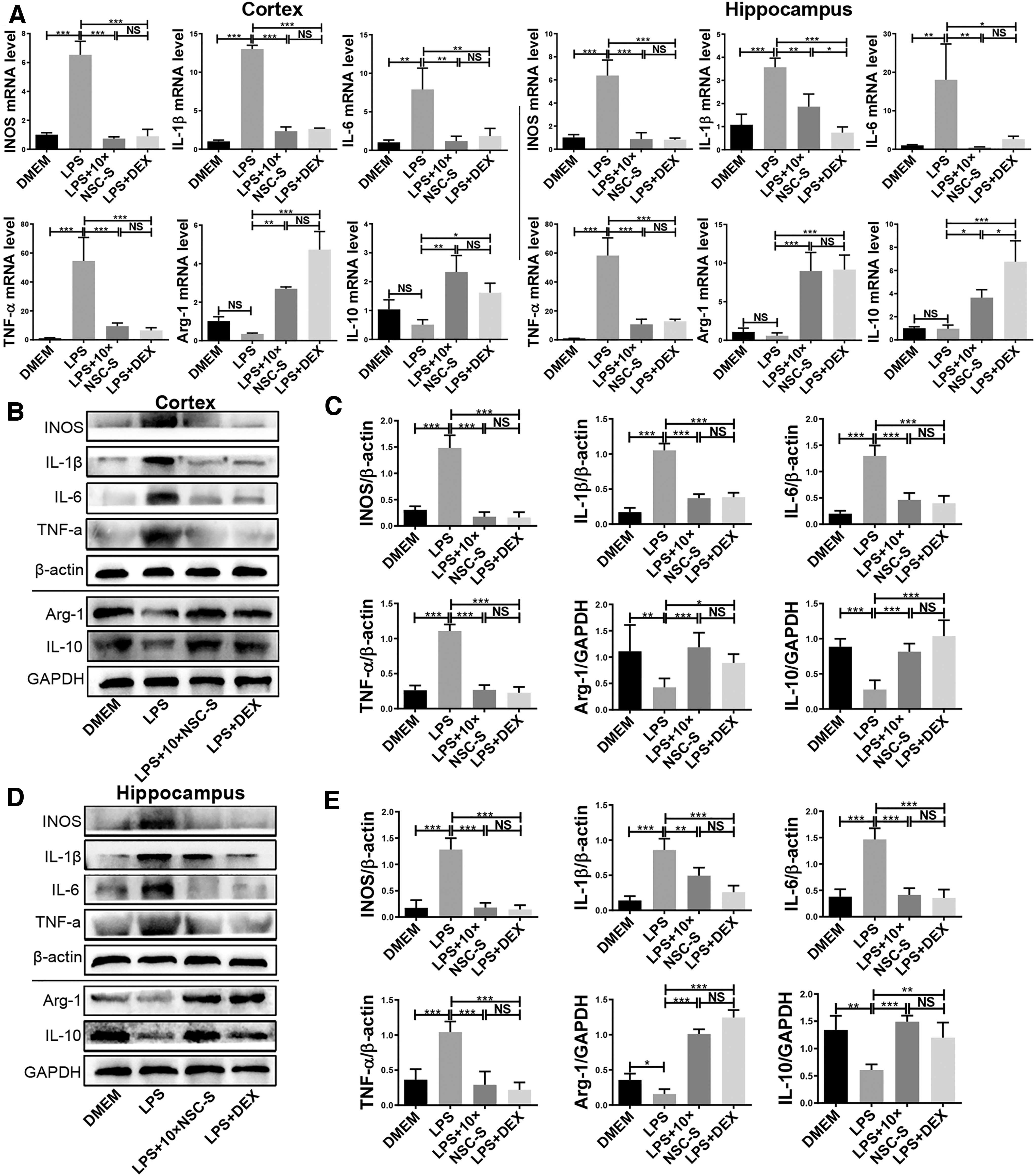
Effects of NSC-S on proinflammatory and anti-inflammatory cytokine expressions in the brain.
NSC-S exerts the polarization of BV2 cells from M1 to M2 phenotype
CD86 and CD206 are surface markers of M1 and M2 phenotypes of microglia, respectively, and their expression in BV2 cells was detected by immunofluorescence staining. The results showed that the expression of CD86 marker was enhanced, whereas CD206 was reduced by LPS stimulation, however, NSC-S treatment produced an opposite effect in the BV2 cells (Fig. 4A, B). Iba-1, which is the marker of microglia activation, was observed as a significantly decreased expression by NSC-S in the LPS-treated BV2 cells (Supplementary Figure S1A). Additionally, M1-related proinflammatory mediators (ROS, NO) and cytokines (iNOS, IL-1β, IL-6, TNF-α, colony-stimulating factor 3, lipocalin 2) were decreased, whereas M2-related anti-inflammatory cytokines (Arg-1, IL-10) were increased in BV2 cells treated with NSC-S, as the opposite effect was observed in the LPS group (Supplementary Figure S1B–N).
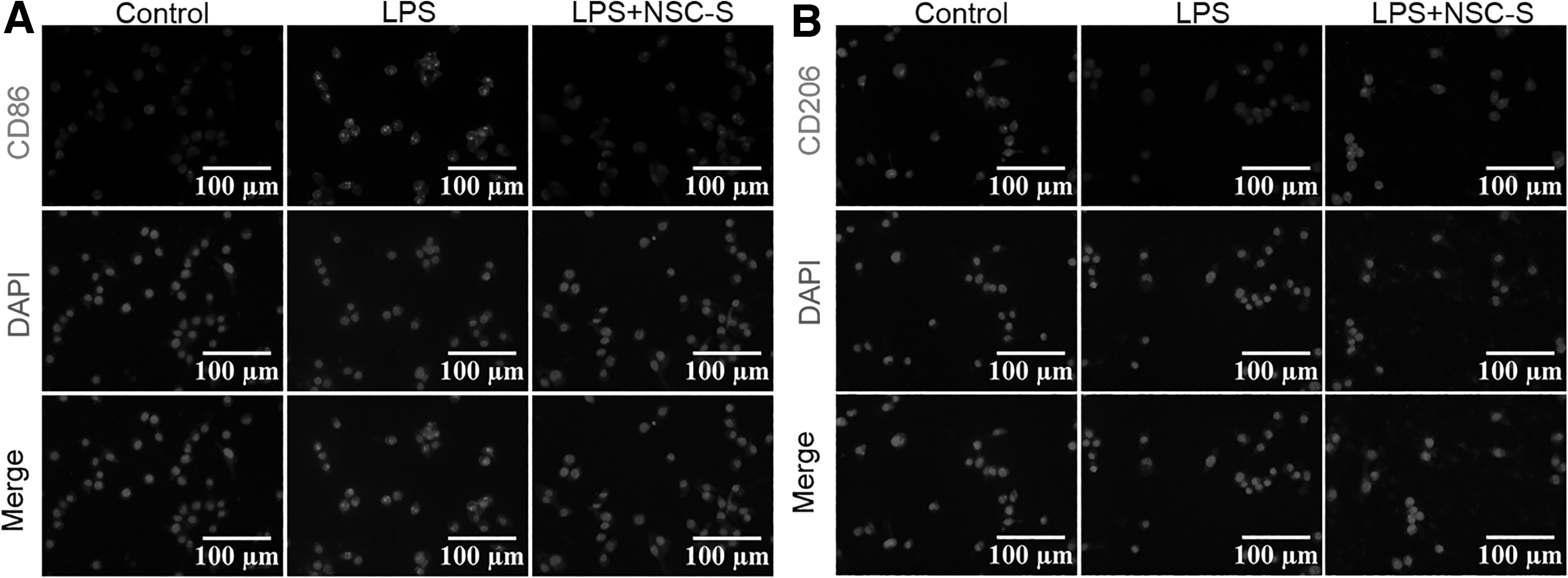
NSC-S exerts the polarization of BV2 cells from M1 to M2 phenotype.
NSC-S regulates neuroinflammatory response in LPS-induced BV2 cells by activating the PPAR-γ pathway
Further experiments revealed that NF-κB was the central regulator of inflammation. Moreover, protein–protein interaction analysis showed that NF-κB (rela proto-oncogene [REL]) and PPAR-γ participated in the regulation of inflammatory factors, where IKBα served as the link between NF-κB and PPAR-γ (Fig. 5A). GW9662, a specific antagonist of PPAR-γ, was applied to inhibit the activation of the PPAR-γ signaling pathway in NSC-S-treated BV2 cells. Western blot results showed that NSC-S administration significantly promoted PPAR-γ activation and repressed NF-κB activation. However, pretreatment with GW9662 blocked the activation of PPAR-γ and inhibition of NF-κB nuclear translocation exerted by NSC-S. In addition, the suppressive effect of NSC-S on the phosphorylation of IKBα, induced by LPS was partly abolished by GW9662 (Fig. 5B–F).
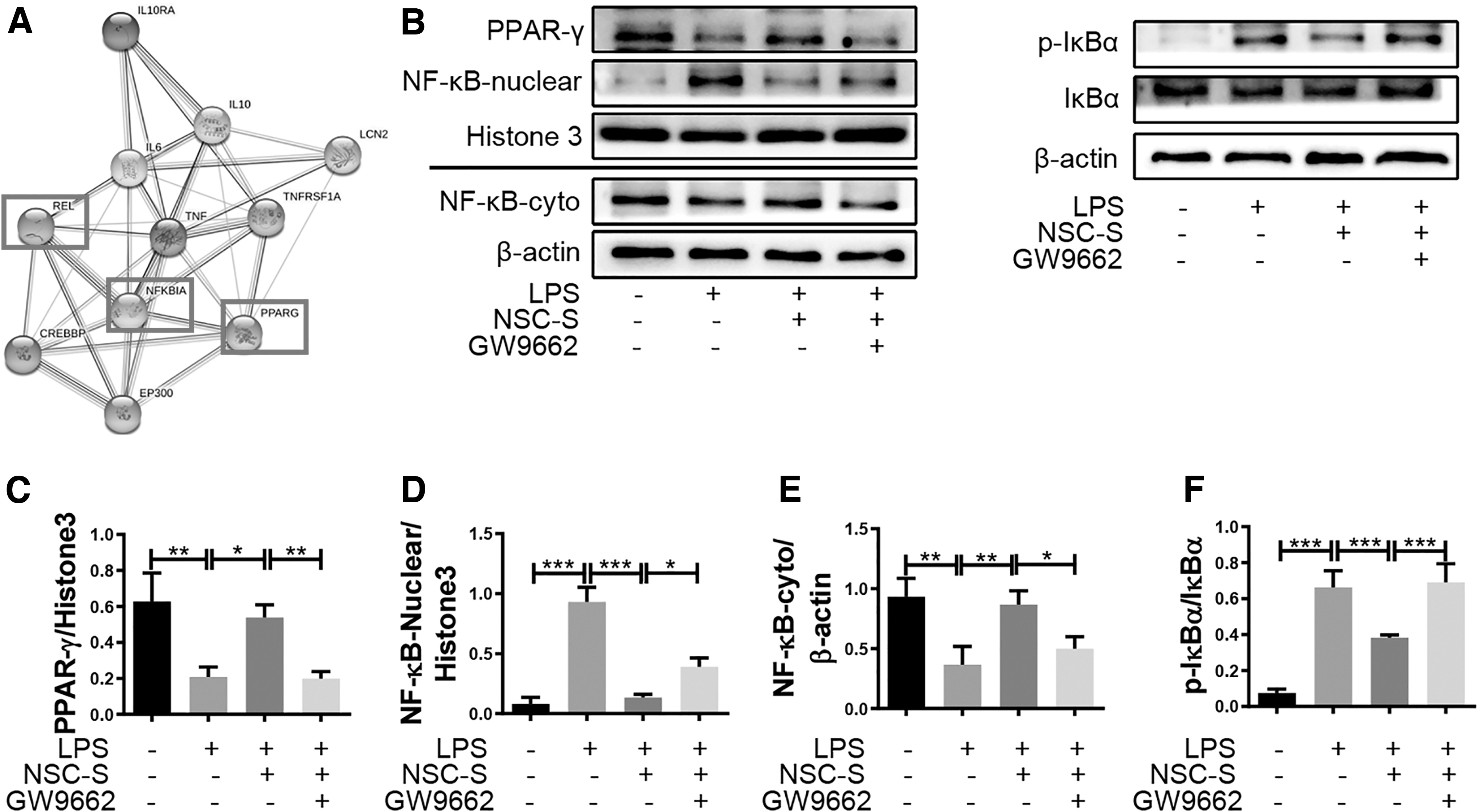
NSC-S inhibits nuclear translocation of NF-κB through promoting the activation of PPAR-γ in LPS-treated BV2 cells.
In an additional analysis, qRT-PCR and western blot results showed that GW9662 restrained the NSC-S-induced downregulation of proinflammatory cytokines, iNOS, IL-1β, IL-6, and TNF-α, as well as the upregulation of anti-inflammatory cytokines, Arg-1 and IL-10, in the LPS-induced BV2 cells (Fig. 6A–G). Immunofluorescence staining results also indicated that GW9662 pretreatment diminished the NSC-S-induced inhibition of M1 marker CD86 expression and the promotion of M2 marker CD206 expression (Fig. 6H, I).
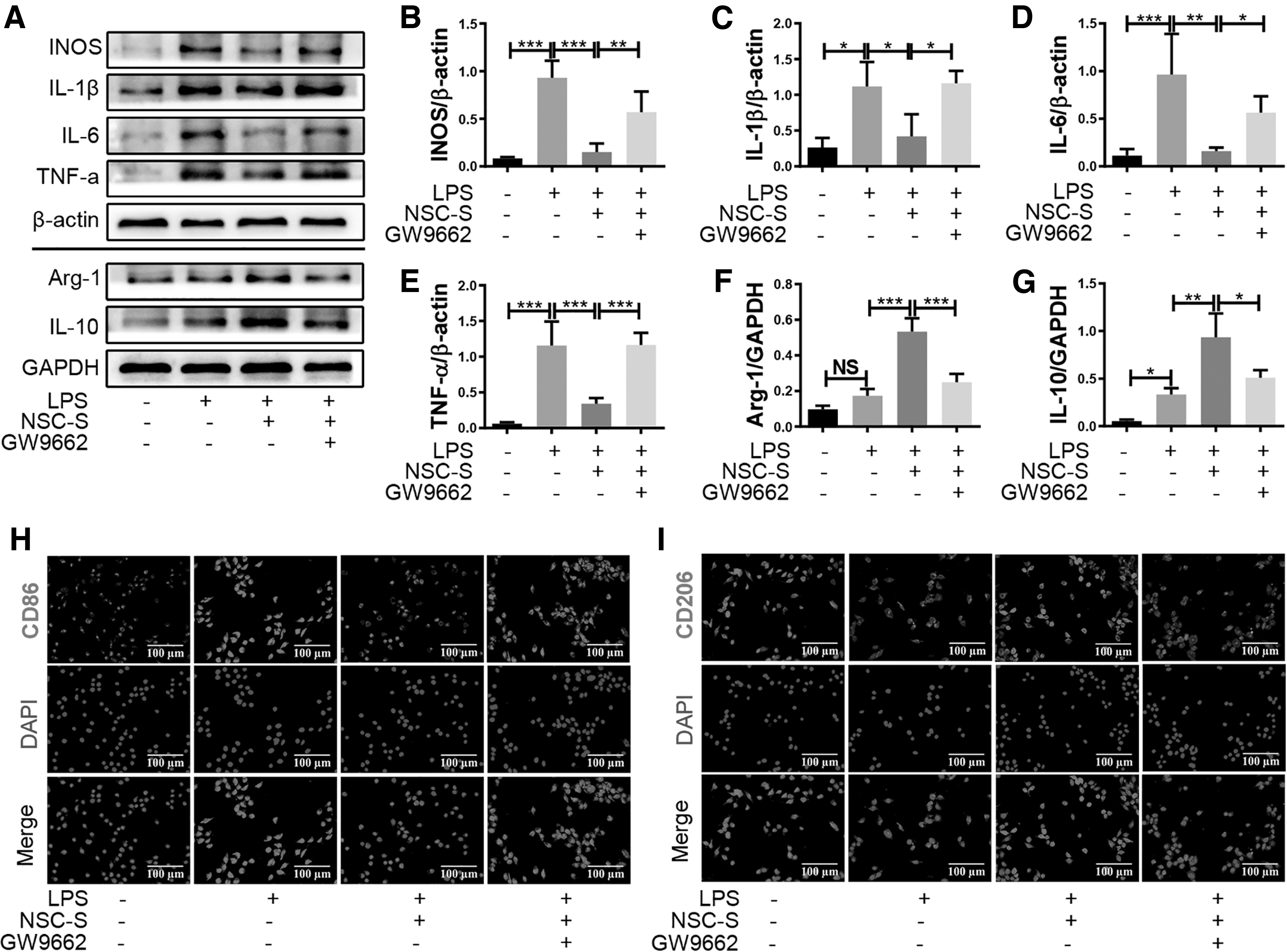
NSC-S regulates inflammatory cytokines and phenotypes in LPS-simulated BV2 cells by activating PPAR-γ.
NanoLiquid chromatography–tandem mass spectrometry analysis of NSC-S
Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was performed on the differently expressed proteins between NSC-S and control medium identified by NanoLiquid chromatography–tandem mass spectrometry (NanoLC-MS/MS). It was shown that four proteins, fatty acid-binding protein 5 (FABP5), acyl-coA-binding protein (DBI), lipoprotein lipase, and FABP7, were involved in the PPAR pathway (Fig. 7A). Gene Ontology (GO) analysis revealed that, of the four proteins, FABP5 and DBI had a localized extracellular form (Fig. 7B). Further analysis through protein–protein interactions showed that FABP5, rather than DBI was functionally associated with PPAR-γ and NF-κB, through the use of the STRING resource analysis (Fig. 7C). The relative concentration of FABP5 in NSC-S was about 11.59 times more than that of DMEM (Table 2).

The bioinformatics on NSC-S through NanoLC-MS/MS analysis.
Differently Expressed Fatty Acid-Binding Protein 5 in Neural Stem Cell Secretome (Mean A) and Dulbecco's Modified Eagle's Medium (Mean C)
The bold showed that the active proteins screened from NSC-S and their relative concentration differences.
FABP5 is involved in regulating LPS-induced inflammatory response in BV2 cells
qRT-PCR results showed that treatment of BV2 cells with recombinant human FABP5 protein (rhFABP5) reduced M1-related proinflammatory factor expression and elevated M2-related anti-inflammatory factor expression in a concentration-dependent manner (Fig. 8A). Notably, the protein level of nuclear PPAR-γ in rhFABP5-treated BV2 cells showed an increasing trend compared with LPS-induced BV2 cells (Fig. 8B, C).
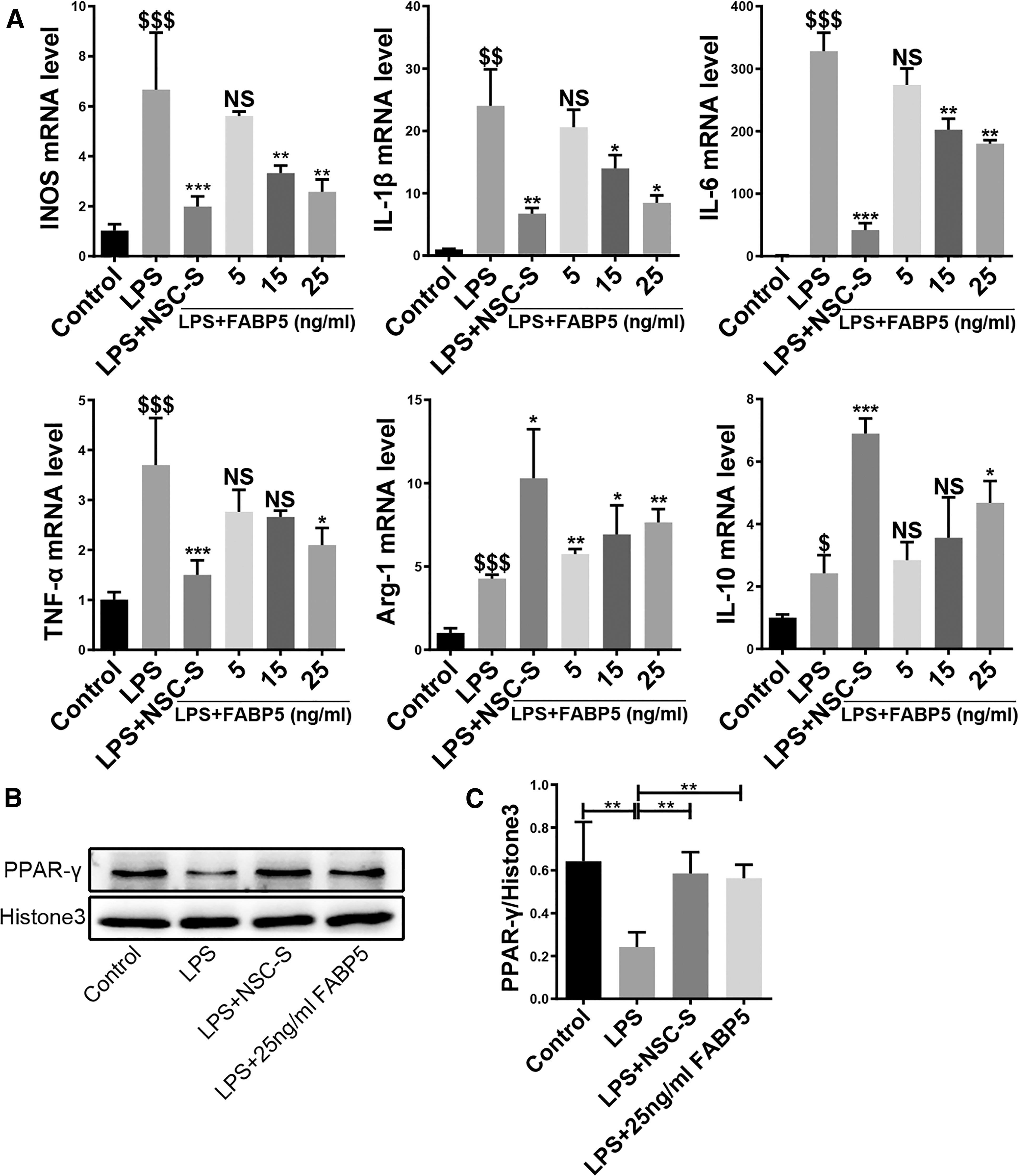
FABP5 inhibits inflammatory response in LPS-induced BV2 cells.
Discussion
Neuroinflammation is an important event and risk factor in many neurodegenerative diseases and is considered as a novel therapeutic target for neurodegenerative diseases [27 –29]. Stem cell therapy has attracted increasing attention in recent years. NSCs are regarded as seed players for transplantation due to their strong ability of self-renewal, multidirectional differentiation potential, immunosuppression, and immunomodulatory effects [30 –34]. However, stem cell transplantation still faces some challenges, which limit its application of stem cell transplantation as a routine choice for clinical treatment [35]. Therefore, there is an urgent need to seek alternative treatment options and strategies. A number of studies have found that the immunomodulatory effects of transplanted stem cells are mainly attributed to bioactive molecules (growth factors, chemokines, cytokines) produced by paracrine mechanisms [36 –38].
To that effect, stem cell secretome contains plentiful paracrine secretions and these substances have been shown to play important roles in the mitigation of diseases [39 –41]. Compared with stem cell transplantation therapy, secretome of stem cells is easier to obtain, preserve, cross the biological barrier, and has the advantages of high bioactivity and low immunogenicity [42,43]. Therefore, stem cell secretome is considered as a promising alternative tool for cell therapy.
In this study, the effect of NSC-S treatment on LPS-induced neuroinflammation was investigated. Previous studies have reported that peripheral challenge of LPS in mice resulted in sickness behavior accompanied by activation of microglia in the central nervous system. Therefore, LPS-induced neuroinflammation represents one of the standard models for studying neuroinflammation [44 –46]. Herein it was found that NSC-S significantly ameliorated sickness behavior and neuroinflammation in LPS-induced mice. Specifically, NSC-S treatment markedly increased the head swing frequency in tail suspension with head swing test, the total traveling distance, and the time spent at the center in OFT. Interestingly, white secretions observed from the eyes of the LPS group was ameliorated by NSC-S treatment. In comparison to DEX, a type of nonsteroidal anti-inflammatory drug commonly used to treat inflammation [47,48], NSC-S exhibited similar level of therapeutic effects, indicating that NSC-S facilitates the improvement of sickness behavior through anti-inflammatory induction in mice.
As the first defense line in the central nervous system's immunization, microglia activation plays a pivotal role in neuroinflammation [49]. Studies have reported that microglia can be functionally identified to M1 and M2 phenotypes, which are closely associated with the severity of neurodegenerative diseases [50 –52]. Hence, maintaining the balance between M1 and M2 immunophenotypes is the key to the treatment of neuroinflammation. In vitro results showed that NSC-S treatment decreased the expression of CD86 and increased the expression of CD206. Also, NSC-S treatment significantly inhibited the activation of microglia and release of M1-related iNOS, IL-1β, IL-6, and TNF-α, but promoted the expression of M2-related Arg-1 and IL-10. These results implicated that NSC-S can alleviate inflammatory response induced by LPS through modulating microglia immunophenotypes.
NF-κB, a critical transcription factor, modulates multiple expression of proinflammatory mediators and plays a crucial role in the development of various inflammation-associated diseases [53 –56]. PPAR-γ belongs to the nuclear hormone receptor superfamily and following activation by ligands, it migrates to initiate transcription [57]. PPAR-γ activation exhibits antioxidant and anti-inflammatory functions in various diseases [58 –61]. Emerging studies revealed that PPAR-γ agonists' administration could effectively suppress NF-κB activation and inflammatory response [62,63]. In this study, the functional connection between PPAR-γ and NF-κB was further confirmed. It was shown that NSC-S significantly elevates PPAR-γ expression and prevented NF-κB activation. Notably, GW9662, a PPAR-γ inhibitor, blocked the inhibition of NSC-S on NF-κB activation and inflammatory response in LPS-induced BV2 cells, indicating that NSC-S plays anti-inflammatory role in a PPAR-γ-dependent manner.
To explore the specific substances that play immunomodulatory role in NSC-S, the proteomic in NSC-S were identified by NanoLC-MS/MS and the bioinformatics analyzed. KEGG and GO analyses revealed FABP5 and DBI in NSC-S to be possibly involved in the PPAR pathway. However, whether these proteins are involved in the PPAR-γ pathway in microglia is worth exploring. Based on the protein–protein interaction analysis of the two proteins, it was found that the expression difference of FABP5 in NSC-S was about 11.59 times that of the control medium. Thus, our subsequent study focused on FABP5. FABP5, a member of the FABP family, has a high affinity for fatty acids and is involved in the activation of PPAR-γ [64,65]. It is reported that FABP5 deficiency significantly worsens inflammatory pathological changes and negatively modulates PPAR-γ activity in lung diseases [66,67]. In another report, FABP5 was identified as a critical anti-inflammatory protein because it exerts inflammation resolution and PPAR-γ activity.
Interestingly, this study showed that FABP5 protein levels were found to positively correlate with PPAR-γ activity in human peripheral blood mononuclear cells [68]. These findings support the positive role of FABP5/PPAR-γ in inflammatory regulation. The current study demonstrated that rhFABP5 inhibited M1-related proinflammatory cytokines and promoted M2-related anti-inflammatory cytokine expression in a dose-dependent manner, and has well-enhanced PPAR-γ activity in BV2 cells, indicating that NSC-S might regulate the anti-inflammatory effect and PPAR-γ activation through FABP5. Contrary to this observation, a study found FABP5 to induce inflammation [69]. The different outcomes could be attributed to differences in drug inducers, cell types, and disease models, which require further exploration.
Conclusion
In summary, this study demonstrates that NSC-S possesses antineuroinflammatory and neuroprotective effects against LPS-induced neuroinflammation models in vivo. The mechanism may be associated with the activation of the PPAR-γ signaling pathway, which subsequently inhibits the activation of the NF-κB signaling pathway, thus, promoting the phenotypic polarization of microglia from M1 to M2, reducing the secretion of inflammatory mediators, and increasing the expression of anti-inflammatory cytokines. It is worth noting that FABP5 may play a crucial role in these processes. Therefore, FABP5 might open novel perspectives for the treatment of neuroinflammation, however, further studies are needed (Supplementary Figure S2).
Footnotes
Acknowledgment
The authors thank Professor Weining Zhang and Associate Professor Jia Wang, Jiangsu University, for his guidance on this study.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
The authors declare no potential conflicts of interest.
Funding Information
This study was supported by the National Natural Science Foundation of China (Grant No. 81571221), the Science and Technology Cooperation Foundation of Health bioMed (Grant No. 20200605), and A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.