
Abstract
Triple-negative breast cancer (TNBC) is a highly aggressive and invasive type of breast cancer. In addition, type 2 diabetes mellitus (T2DM) is recognized as a risk factor for cancer metastasis, which is associated with mortality in patients with breast cancer. Cancer-associated fibroblasts (CAFs) generated from adipose tissue-derived mesenchymal stem cells (AT-MSCs) play a vital role in the progression of TNBC. However, to date, whether T2DM affects the ability of AT-MSCs to differentiate into CAFs is still unclear. In this study, we found that in coculture with TNBC cells [breast cancer cells (BCCs)] under hypoxic conditions, AT-MSCs derived from T2DM donors (dAT-MSCs) were facilitated to differentiate into CAFs, which showed fibroblastic morphology and the induced expression of fibroblastic markers, such as fibroblast activation protein, fibroblast-specific protein, and vimentin. This was involved in the higher expression of transforming growth factor beta receptor 2 (TGFβR2) and the phosphorylation of Smad2/3. Furthermore, T2DM affected the fate and functions of CAFs derived from dAT-MSCs. While CAFs derived from AT-MSCs of healthy donors (AT-CAFs) exhibited the markers of inflammatory CAFs, those derived from dAT-MSCs (dAT-CAFs) showed the markers of myofibroblastic CAFs. Of note, in comparison with AT-CAFs, dAT-CAFs showed a higher ability to induce the proliferation and in vivo metastasis of BCCs, which was involved in the activation of the transforming growth factor beta (TGFβ)-Smad2/3 signaling pathway. Collectively, our study suggests that T2DM contributes to metastasis of BCCs by inducing the myofibroblastic CAFs differentiation of dAT-MSCs. In addition, targeting the TGFβ-Smad2/3 signaling pathway in dAT-MSCs may be useful in cancer therapy for TNBC patients with T2DM.
Introduction
Breast cancer is the most commonly diagnosed cancer worldwide. According to global cancer statistics, breast cancer accounted for 11.7% of newly diagnosed cancers in 2020 [1]. Approximately 15%–20% of breast cancer patients are diagnosed with triple-negative breast cancer (TNBC), which is highly invasive and metastatic [2]. Metastasis, which reduces survival time in 46% of TNBC patients, is a multistage biological process whereby cancer cells spread to other parts of body [2,3]. In cancer metastasis, intercellular signaling, together with the hypoxic condition inside the tumor microenvironment, regulates the tumor behavior and enhances the migratory ability of tumor cells [4 –7].
The tumor microenvironment, which consists of several types of cells [eg, immune cells, endothelial cells (ECs), cancer-associated fibroblasts (CAFs), and mesenchymal stem cells (MSCs)], provides a favorable environment for tumor growth, and regulates tumor progression and metastasis [6]. Among the cell populations inside the tumor, CAFs play a vital role in support of tumor progression and metastasis [7]. Several studies suggest that the CAFs enhance the migratory ability and the aggressive features of cancer cells [8,9]. Therefore, targeting CAFs is considered to be a promising approach in cancer therapy.
CAFs represent the largest heterogeneous cell population; myofibroblastic CAFs and inflammatory CAFs represent the major subtypes in tumors [7]. Myofibroblastic CAFs, which are identified based on the expression of alpha smooth muscle actin (α-SMA), are located at the center of the tumor, which directly interacts with and supports the tumor cells [7]. Meanwhile, inflammatory CAFs, which express inflammatory cytokines, such as CXCL1 and interleukin (IL)6, surround the tumor and secrete inflammatory cytokines to support tumor development [7].
Previous studies have suggested that in the tumor microenvironment, CAFs are generated from various types of progenitors, including fibroblasts, ECs, and MSCs, in response to signaling from cancer cells and the induction of hypoxia [10 –12]. Several studies have suggested that hypoxia facilitates the reprogramming of normal fibroblasts into CAFs, which support tumor growth and metastasis [13,14]. Furthermore, hypoxic induction enhances both the tumor-promoting functions of CAFs, which elevates the migratory ability of tumor cells and creates a favorable environment for tumor metastasis [15,16].
MSCs are a population of tissue stem cells that reside in the tumor microenvironment, and which participate in the regulation of tumor progression and metastasis. Bone marrow MSCs have been reported to increase the migratory ability of breast cancer cells (BCCs) through CCL5 signaling [17]. Other studies reported that the secretions of adipose tissue-derived MSCs (AT-MSCs) promote the epithelium–mesenchymal transition (EMT) and tumor growth of BCCs [18,19]. Of note, several studies have reported that MSCs are a progenitor of CAFs in the tumor [20]. MSCs possess the ability to differentiate into CAFs through the transforming growth factor beta (TGFβ) signaling secreted by cancer cells [21]. In addition, CAFs generated from AT-MSCs induced by the secretome of TNBC cells enhance the invasive ability of these TNBC cells [22].
Numerous studies have reported the effects of diseases of donors, such as type 2 diabetes mellitus (T2DM), on the characteristics and functions of MSCs. T2DM has been recognized as a risk factor that induces metastasis and leads to poor outcomes in patients with breast cancer [23]. Of note, we previously reported that T2DM induces paracrine effects of AT-MSCs to promote metastasis of TNBC cells [24]. However, to date, the relationship between T2DM and CAFs differentiation of AT-MSCs is still unclear. In this study, we investigate how T2DM affects the ability of AT-MSCs to differentiate into CAFs. In addition, the mechanisms involved are clarified to suggest a candidate treatment for TNBC.
Materials and Methods
Ethics statement
The experiments were conducted according to the amended Declaration of Helsinki and were approved by the Ethics Committee of the University of Tsukuba. The collection of human samples was performed with the informed consent of the donors. The in vivo experiments were approved by the Animal Care Committee of the University of Tsukuba.
Cell isolation and culture
Human adipose tissues were collected from healthy donors (female, n = 4; average age, 80 years, HbA1c <6) or T2DM donors (female, n = 4; average age, 80 years, HbA1c >6.5) from the Department of Cardiovascular Surgery, University of Tsukuba Hospital, Tsukuba, Japan. Adipose tissues were minced and digested with 0.1% collagenase (Invitrogen, Waltham, MA) in phosphate-buffered saline (PBS) at 37°C for 1 h. After that, the mixture was centrifuged at 1,600 rpm in 7 min, and the cell pellets were harvested. Cells were cultured in MSCs culture medium containing Iscove's Modified Dulbecco's Medium (IMDM, Invitrogen) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Invitrogen), 2 mg/mL
MDAMB231 cells (BCCs) were purchased from the American Type Culture Collection (ATCC, Manassas, VA). BCCs were cultured in IMDM medium, supplemented with 1% Penicillin–Streptomycin, 5% FBS, and 2 mg/mL
Proliferation assay
MSCs or BCCs were collected from the 24-well cell culture dish. The cell number was determined by staining MSCs with 0.4% trypan blue stain (Gibco, Waltham, MA) and counting the live cells by a hemocytometer.
CAFs differentiation
AT-MSCs (1 × 105 cells/well) were seeded in a 24-well cell culture dish and cultured at 37°C under 5% CO2 overnight. Transwells (3.0 μm; Corning, New York) were placed in each well of 24-well cell culture dish, and BCCs (1 × 105 cells/well) were placed in the transwells. The transwell coculture assay was performed at 37°C under 1% O2 for 14 days. The media in the 24-well cell culture dish and transwells were changed to fresh medium every 2 days. After 14 days, the cells were prepared for the subsequent examination.
Immunofluorescence staining
CAFs derived from MSCs were washed with PBS two times, then 1 mL of 4% paraformaldehyde was added to fix the cell. After that, 1 mL of perforation buffer was added for 10 min, and cells were washed with PBS. Then, 1 mL of blocking buffer was used to block the cells on glass coverslips for 30 min under room temperature. After removing the blocking buffer, cells were incubated with rabbit anti-vimentin as primary antibody (1:500, ab137321; Abcam, Cambridge, United Kingdom) at 4°C overnight. After being washed, the cells were incubated with goat antirabbit IgG second antibody as secondary antibody and DAPI (1:500, DAPI 1:1,000, ab150077; Abcam) for 1 h at room temperature. The antibody solution was removed, and the cells were washed three times for 5 min. The fluorescence signal of cells was observed under a microscope (BZ-X710; Keyence Corporation, Osaka, Japan).
Quantification of immunofluorescence
All images with a size of 640 × 480 pixels were obtained by a fluorescent microscope (Keyence Corporation) at 40 × magnification. The fluorescence signals of vimentin-GFP were taken at channel 1, and of nuclei-DAPI were taken to examine the single cells in the field. The fluorescence signals of vimentin were analyzed by measuring corrected total cell fluorescence (CTCF), as described in a previous study [25]. To measure CTCF, images with GFP signals were applied with gray scaling by a FIJI software (version 2.1.0/1.53c).
In each image, a single cell will be randomly selected, and the area, integrated density, and mean gray value were measured. To remove the background, three areas without fluorescence signals, which are near to the targeted cells, were selected with the same size, and mean fluorescence of background readings was measured to normalize against the autofluorescence. Then, CTCF was calculated as CTCF = integrated density − (area of selected cell × mean fluorescence of background readings).
Cell migration assay
BCCs were seeded with a number of 105 cells/well in the 24-well cell culture dish, and cultured at 37°C under 1% O2 and 5% CO2 for 24 h to reach 100% confluence. After that, the cells were treated with 10 μg/mL mitomycin C for 2 h at 37°C under 1% O2 and 5% CO2. A scratch was created; then cells were washed with PBS three times. The migration of cells was observed every 6 h under a microscope (BZ-X710; Keyence Corporation).
Gene expression analysis
To isolate total RNA, Sepasol-RNA I Super G (Nacalai Tesque, Kyoto, Japan) was used in accordance with the manufacturer's protocol. Then, cDNA was synthesized using a reverse transcription polymerase chain reaction (RT-PCR) kit (Toyobo, Osaka, Japan) with 1 μg of total isolated RNA. To examine the expression of target genes, a quantitative polymerase chain reaction (PCR) was performed with SYBR green reagent (Toyobo) using a GeneAmp 7500Fast Real-Time PCR System (Applied Biosystems, Waltham, MA).
The sequences of the primer sets used for PCR are shown in Table 1. β-Actin was used as an internal control in all samples. The expression levels of the target genes were analyzed using comparative CT method, also referred to as the 2−ΔΔC T method, as described previously [26]. To compare the gene expression in two different samples, the fold change was calculated as: fold change = 2−ΔΔC T, in which
Primers Used for Quantitative Polymerase Chain Reaction
α-SMA, alpha smooth muscle actin; FAP, fibroblast activation protein; FSP, fibroblast-specific protein; IL, interleukin; LIF, leukemia inhibitory factor; TGFβ, transforming growth factor beta; TGFβR2, transforming growth factor beta receptor 2.

Cellular interaction assay
BCCs (1 × 105 cells/well) were seeded in a 24-well cell culture dish and cultured at 37°C under 1% O2 for 6 h. Transwells (3.0 μm) were placed in each well, and AT-MSCs or AT-MSC-differentiated CAFs (1 × 105 cells/well) were seeded in the transwells. After culturing at 37°C under 1% O2 for 48 h, the cells were harvested for the following examination.
Western blotting
Total protein was extracted from BCCs according to a previously described report [27]. After that, an amount of 30 μg total protein was applied to a SDS (sodium dodecyl-sulfate)–polyacrylamide gels for electrophoresis, then transferred onto polyvinylidene fluoride membranes (Merck Millipore, Burlington, MS). The membranes were then blocked with 5% skim milk in TBST and incubated with rabbit anti-Smad2/3 antibody (sc-398844; Santa Cruz Biotechnology, Dallas, TX), anti-pSmad2/3 antibody (sc-11769; Santa Cruz Biotechnology), or rabbit anti-β-actin antibody (GTX109639; GeneTex, Irvine, CAS) at 1:1,000 dilution at 4°C overnight.
After that, the membranes were washed with TBST and incubated with secondary antibody horseradish peroxidase (HRP)-conjugated goat antirabbit IgG (Invitrogen), at a dilution of 1:10,000 at room temperature for 2 h. To detect the expression of protein, membranes were incubated with chemiluminescence HRP substrate (EMD Millipore) for 1 min and detected by an Image Quant LAS 4000 System (GE Healthcare, Chicago, IL).
In vivo metastatic assay
Female C57BL/6 mice were purchased from Charles River Japan, Inc. (Yokohama, Japan). Mice were maintained on a 12-h light/dark cycle with ad libitum access to food and water. Eight-week old mice were used in this experiment. In brief, a number of 105 BCCs were intravenously injected into mice through the tail vein. Then, mice were injected with cyclosporin A (20 mg/kg; Wako) every day for 2 weeks. Mice were sacrificed, and lung tissues were collected and fixed in 4% paraformaldehyde after being embedded in OCT compound (Sakura Finetek, Tokyo, Japan) and frozen in liquid nitrogen. The migration of BCCs to lung tissues was determined by hematoxylin and eosin staining.
Statistical analysis
Data were analyzed by the Mann–Whitney U-test using the GraphPad Prism 5 software program (GraphPad Software, San Diego, CA) and are presented as the mean ± standard deviation. P values of <0.05 were considered to indicate statistical significance.
Results
Coculture with TNBC cells (BCCs) induced the differentiation of AT-MSCs to CAFs
Coculture with cancer cells is reported to induce the differentiation of MSCs to CAFs [22]. In addition, hypoxia plays a vital role in the tumor microenvironment, which stimulates paracrine signaling between tumor cells and other cells, facilitating fibroblast reprogramming and CAFs differentiation [16]. Therefore, we first examined the ability of AT-MSCs derived from healthy donors, at the average of 80, to differentiate into CAFs in coculture with BCCs, MDAMB231 cells under normoxic condition and hypoxic condition. The cell morphology showed that at day 14 post-coculturing in either normoxic or hypoxic conditions, nAT-MSCs cocultured with BCCs (AT-CAFs) showed a spindle shape that was similar to human fibroblasts (Fig. 1A).
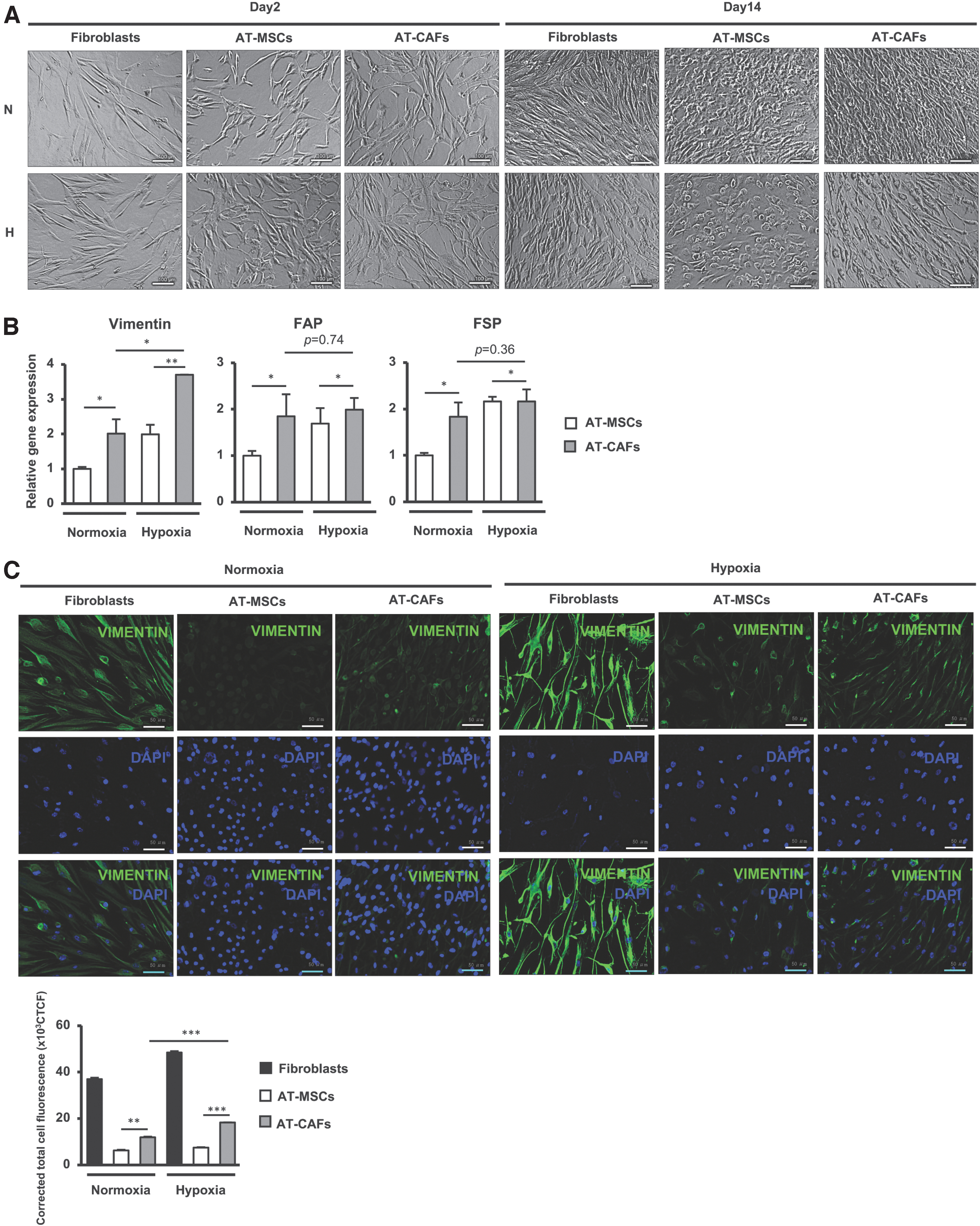
Hypoxia induced the ability of AT-MSCs to differentiate into CAFs.
Next, we examined the expression of genes related to CAFs differentiation in AT-MSCs cocultured with BCCs after 14 days. As shown in Fig. 1B, in comparison with AT-MSCs, AT-CAFs showed the induced expression of genes related to CAFs differentiation [28 –31], including fibroblast activation protein (FAP, under normoxic conditions: 1.85-fold increase, under hypoxic conditions: 2-fold increase), fibroblast-specific protein (FSP, under normoxic conditions: 1.83-fold increase, under hypoxic conditions: 2.16-fold increase), and vimentin (under normoxic conditions: 2.01-fold increase, under hypoxic conditions: 3.7-fold increase).
Since the expression of vimentin, one of the markers of CAFs, was strikingly upregulated in AT-CAFs under hypoxic conditions, we further confirmed the expression of vimentin protein by staining of AT-MSCs with an antivimentin antibody. The results showed that in comparison with AT-MSCs, AT-CAFs expressed a higher level of vimentin protein under either normoxic or hypoxic conditions (Fig. 1C). Of note, consistent with the mRNA expression, AT-CAFs under hypoxic conditions showed the upregulation of vimentin protein in comparison with those under normoxic conditions (P < 0.05, n = 3, Fig. 1C).
Taken together, these data suggested that coculturing with BCCs induced the differentiation of AT-MSCs to CAFs, and that this was facilitated under hypoxic conditions.
AT-MSCs derived from T2DM showed a high ability to differentiate into CAFs
T2DM is recognized as a risk factor of BCCs progression and metastasis [23]. Indeed, our previous study showed that AT-MSCs derived from T2DM (dAT-MSCs) highly induced metastasis of BCCs, suggesting that dAT-MSCs may have higher potential to support the progression of breast tumors [24]. Hence, in this study, we determined whether dAT-MSCs possess higher ability to differentiate into CAFs in comparison with AT-MSCs under hypoxic conditions, the major environment inside the tumor.
First, the characteristics of dAT-MSCs were examined, which showed the comparative proliferation, MSC markers, and differentiation ability to osteocytes, in comparison with AT-MSCs derived from healthy donors (Supplementary Fig. S1A–C). In addition, dAT-MSCs exhibited the higher ability to differentiate to adipocytes, in comparison with AT-MSCs derived from healthy donors (Supplementary Fig. S1B).
These results are consistent with our previous study [32]. Moreover, dAT-MSCs secreted a higher amount of inflammatory cytokines, such as IL6 (4.3-fold higher), IL1β (2.2-fold higher), and TNFα (3.7-fold higher), as shown in Supplementary Fig. S1D. Next, the ability of dAT-MSCs to differentiate into CAFs was compared with AT-MSCs derived from healthy donors. The results showed that after 14 days of coculturing with BCCs under hypoxic conditions, cells derived from both AT-MSCs (AT-CAFs) and dAT-MSCs (dAT-CAFs) showed a similar proliferation rate (Fig. 2A) and fibroblast-like morphology. Of note, in comparison with the morphology of AT-CAFs, the morphology of dAT-CAFs was more spindle-like and much more similar to human fibroblasts (Fig. 2B).

dAT-MSCs showed higher ability to differentiate into CAFs in comparison with nAT-MSCs.
In addition, the expression of fibroblast markers in both AT-CAFs and dAT-CAFs was examined. As shown in Fig. 2C, in comparison with AT-CAFs, dAT-CAFs showed the higher expression of fibroblast markers, including FAP (1.44-fold increase), FSP (2.24-fold increase), and vimentin (3.13-fold increase). Consistently, the immunostaining data showed that the vimentin expression of dAT-CAFs was markedly higher than that of AT-CAFs (1.3-fold increase, P < 0.001, n = 3, Fig. 2D). CAFs are a heterogeneous population that are classified into several subpopulations based on the expression of specific biomarkers [28,31]. Therefore, we further examined whether AT-CAFs and dAT-CAFs belong to the same subpopulation by analyzing the expression of biomarkers of CAF subpopulations.
Interestingly, Fig. 2E shows that in comparison with AT-MSCs, AT-CAFs showed the upregulation of markers of inflammatory CAFs, including CXCL1 (4.9-fold increase), IL6 (2.6-fold increase), and leukemia inhibitory factor (LIF) (2.1-fold increase). Meanwhile, in comparison with dAT-MSCs, dAT-CAFs showed the upregulation of markers of myofibroblastic CAFs, including α-SMA (1.6-fold increase) and TPM1 (1.3-fold increase).
Taken together, these data suggest that dAT-MSCs possess a higher ability to differentiate into CAFs under signaling of BCCs under hypoxic conditions in comparison with nAT-MSCs. Notably, AT-CAFs and dAT-CAFs may belong to two different subpopulations, in which dAT-CAFs are inclined to exhibit a myofibroblastic CAFs phenotype.
The activation of TGFβ paracrine signaling induced dAT-MSCs to differentiate into CAFs
A previous study suggested an important role of TGFβ signaling in the generation of CAFs, in which TGFβ from the paracrine singling of BCCs is a key factor in inducing MSCs to differentiate into CAFs [22]. In addition, several studies have suggested that the activation of TGFβ signaling is responsible for the generation of myofibroblastic CAFs [29]. Therefore, we next examined the role of TGFβ signaling in the differentiation of dAT-MSCs to dAT-CAFs.
First, we compared the expression of TGFβ receptors (TGFβR) between AT-MSCs and dAT-MSCs under normoxic or hypoxic conditions. As shown in Fig. 3A, in comparison with AT-MSCs, dAT-MSCs showed significantly higher TGFβR2 expression levels (6.1-fold increase); this was upregulated under hypoxic conditions (2.43-fold increase).
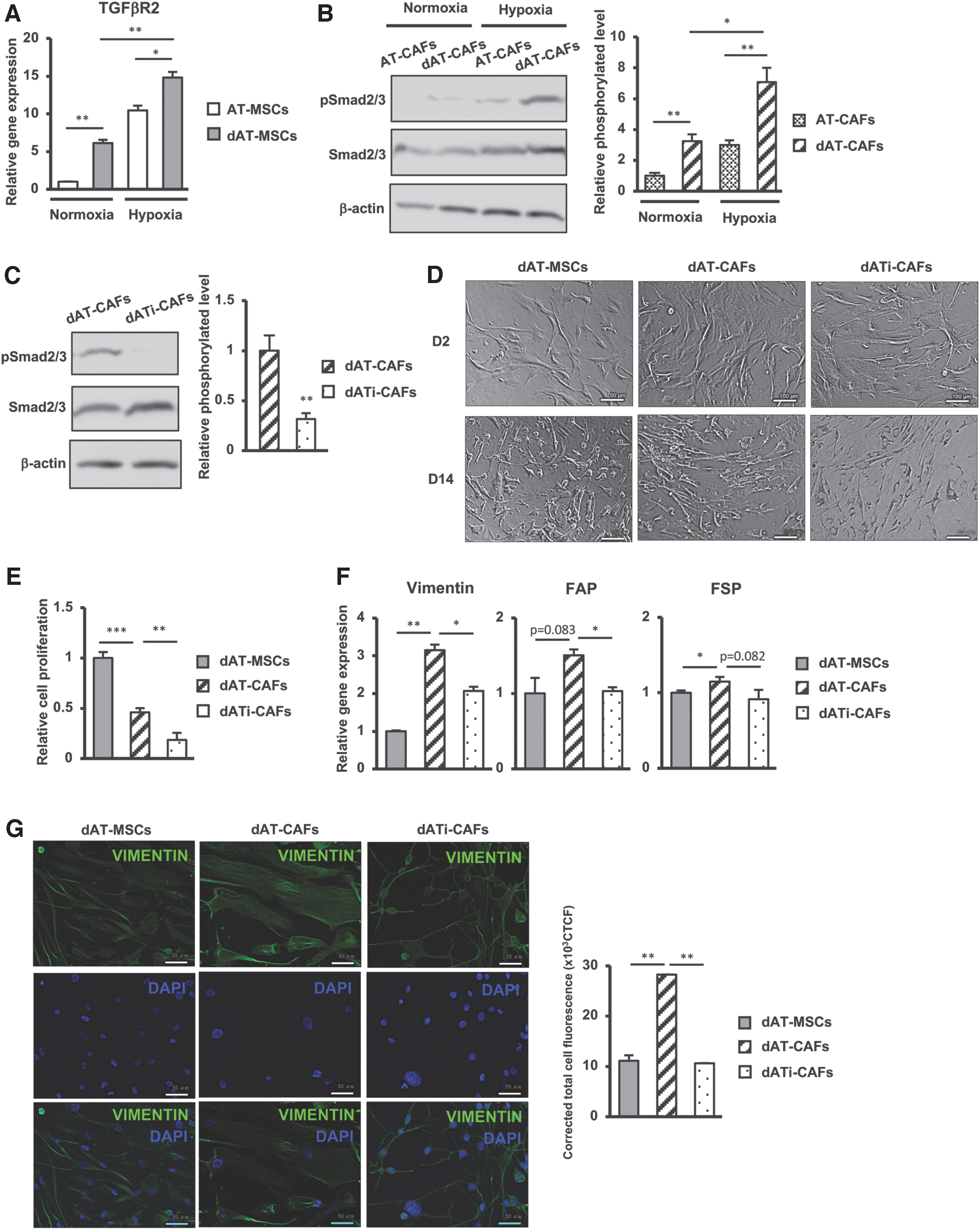
Activation of the TGFβ-Smad2/3 signaling pathway was involved in the high ability of dAT-MSCs to differentiate into CAFs under hypoxic conditions.
Next, we examined the activation of Smad2/3 pathway, a key signaling molecule in the TGFβ signaling pathway involved in the differentiation of CAFs [33], in the AT-CAFs or dAT-CAFs that were derived after 14 days of coculturing with AT-MSCs or dAT-MSCs with BCCs. As shown in Fig. 3B, consistent with the higher expression of TGFβR2 in dAT-MSCs, dAT-CAFs showed the higher expression of phosphorylated Smad2/3 protein in comparison with AT-CAFs (3.2-fold increase), which was upregulated under hypoxic conditions (2.2-fold increase).
To examine the role of the Smad2/3 signaling pathway in the CAFs differentiation of dAT-MSCs, TGFβ inhibitor was added to the coculture of dAT-MSCs with BCCs. As a result, TGFβ inhibitor treatment led to the impaired phosphorylation of Smad2/3 protein in dAT-CAFs (Fig. 3C). In addition, both dAT-CAFs and dAT-CAFs generated in the presence of TGFβ inhibitor (dATi-CAFs) showed a similar fibroblastic morphology (Fig. 3D), whereas the proliferative activity of dATi-CAFs was lower than that of dAT-CAFs (2.56-fold decrease; Fig. 3E). Of note, in comparison with dAT-CAFs, dATi-CAFs showed a significantly decreased expression of fibroblast markers, such as FAP (1.5-fold decrease) and vimentin (1.57-fold decrease; Fig. 3F). Correspondingly, the immunostaining data showed that the protein expression of vimentin was downregulated in dATi-CAFs compared with dAT-CAFs (P < 0.001, n = 3; Fig. 3G).
Taken together, these data suggested that T2DM might induce the CAFs differentiation of AT-MSCs by activating TGFβ signaling pathway. In comparison with AT-MSCs, dAT-MSCs showed the higher expression of TGFβR and activation of Smad2/3, which are involved in the differentiation to CAFs with the signaling from BCCs.
In comparison with AT-CAFs, dAT-CAFs more strongly promoted the metastatic ability of BCCs
We next examined whether the function of dAT-CAFs in supporting the progression of TNBC is stronger in comparison with AT-CAFs, by performing coculturing of BCCs with AT-CAFs (AT-CAFs/BCCs), dAT-CAFs (dAT-CAFs/BCCs), and dATi-CAFs (dATi-CAFs/BCCs), and examined the effects of these CAFs on the phenotypes of BCCs. After coculturing, BCCs were collected and characterized, including the migration, gene expression, in vitro migration, and in vivo metastasis. First, the effects of AT-CAFs, dAT-CAFs, and dATi-CAFs on the proliferation of BCCs were examined.
The results showed that while AT-CAFs/BCCs and dATi-CAFs/BCCs showed a similar proliferation rate with the original BCCs, dAT-CAFs/BCCs exhibited a significantly induced proliferative ability (1.3-fold higher in comparison with the original BCCs; Fig. 4A), suggesting that dAT-CAFs promoted the proliferation of BCCs more than nAT-CAFs. Next, we examined the effects of AT-CAFs, dAT-CAFs, and dATi-CAFs on facilitating the metastatic ability of BCCs.

dAT-CAFs showed a high ability to induce metastasis of BCCs, which was impaired by the inhibition of TGFβ-Smad2/3 signaling pathway.
As shown in Fig. 4B, while dATi-CAFs/BCCs showed similar expression levels of genes related to the EMT in comparison with the original BCCs, AT-CAFs/BCCs and dAT-CAFs/BCCs showed the upregulation of TGFβ (AT-CAFs/BCCs: 1.8-fold increase, dAT-CAFs/BCCs: 2.4-fold increase) and Snail (AT-CAFs/BCCs: 1.7-fold increase, dAT-CAFs/BCCs: 1.8-fold increase), respectively. These data suggested that coculturing with either AT-CAFs or dAT-CAFs significantly induced the expression of genes related to the EMT (eg, TGFβ and Snail) in BCCs.
Next, we examined the effects of AT-CAFs, dAT-CAFs, and dATi-CAFs on the in vitro migratory ability of BCCs. As a result, while AT-CAFs/BCCs, dAT-CAFs/BCCs, and dATi-CAFs/BCCs showed increased migratory ability in comparison with the original BCCs, the induced migration was highest in dAT-CAFs/BCCs (Fig. 4C). These data suggested that dAT-CAFs possess higher ability to promote the migration of BCCs in comparison with AT-CAFs and dATi-CAFs. Furthermore, the effects of AT-CAFs, dAT-CAFs, and dATi-CAFs on the in vivo metastasis of BCCs were examined in the lung metastatic mouse model.
The original BCCs, AT-CAFs/BCCs, dAT-CAFs/BCCs, or dATi-CAFs/BCCs were injected into mice through tail veins, then metastasis of the injected BCCs to lungs was examined. The results showed that mice injected with either AT-CAFs/BCCs or dAT-CAFs/BCCs showed the increased number of tumor foci and tumor size in lungs, in comparison with mice injected with the original BCCs (Fig. 4D).
Of note, in comparison with AT-CAFs/BCCs, dAT-CAFs/BCCs exhibited a higher in vivo metastatic ability (Fig. 4D), suggesting that dAT-CAFs have a higher ability to induce in vivo metastasis of BCCs in comparison with AT-CAFs. Interestingly, mice injected with dATi-CAFs/BCCs showed a similar number of tumor foci in lungs in comparison with mice injected with the original BCCs, although the tumor size is higher (Fig. 4D), suggesting that dATi-CAFs possess the ability to inhibit metastasis of BCCs in vivo.
Taken together, these data suggested that dAT-CAFs had a higher ability to induce metastasis of BCCs in comparison with nAT-CAFs. Notably, the impairment of TGFβ signaling resulted in decreasing CAFs differentiation and declining the effects of dAT-CAFs on metastasis BCCs.
Discussion
In this study, we demonstrated the effects of T2DM on the ability of AT-MSCs to differentiate into CAFs under the paracrine effects of BCCs. In comparison with AT-MSCs, dAT-MSCs showed the higher ability to differentiate into CAFs, which was involved in the upregulation of TGFβR2 and the activation of Smad2/3 in TGFβ signaling pathway. Of note, while AT-MSCs differentiated to inflammatory CAFs, dAT-MSCs differentiated to myofibroblastic CAFs, which showed the higher ability to promote the proliferation, migration, and in vivo metastasis of BCCs (Fig. 5).
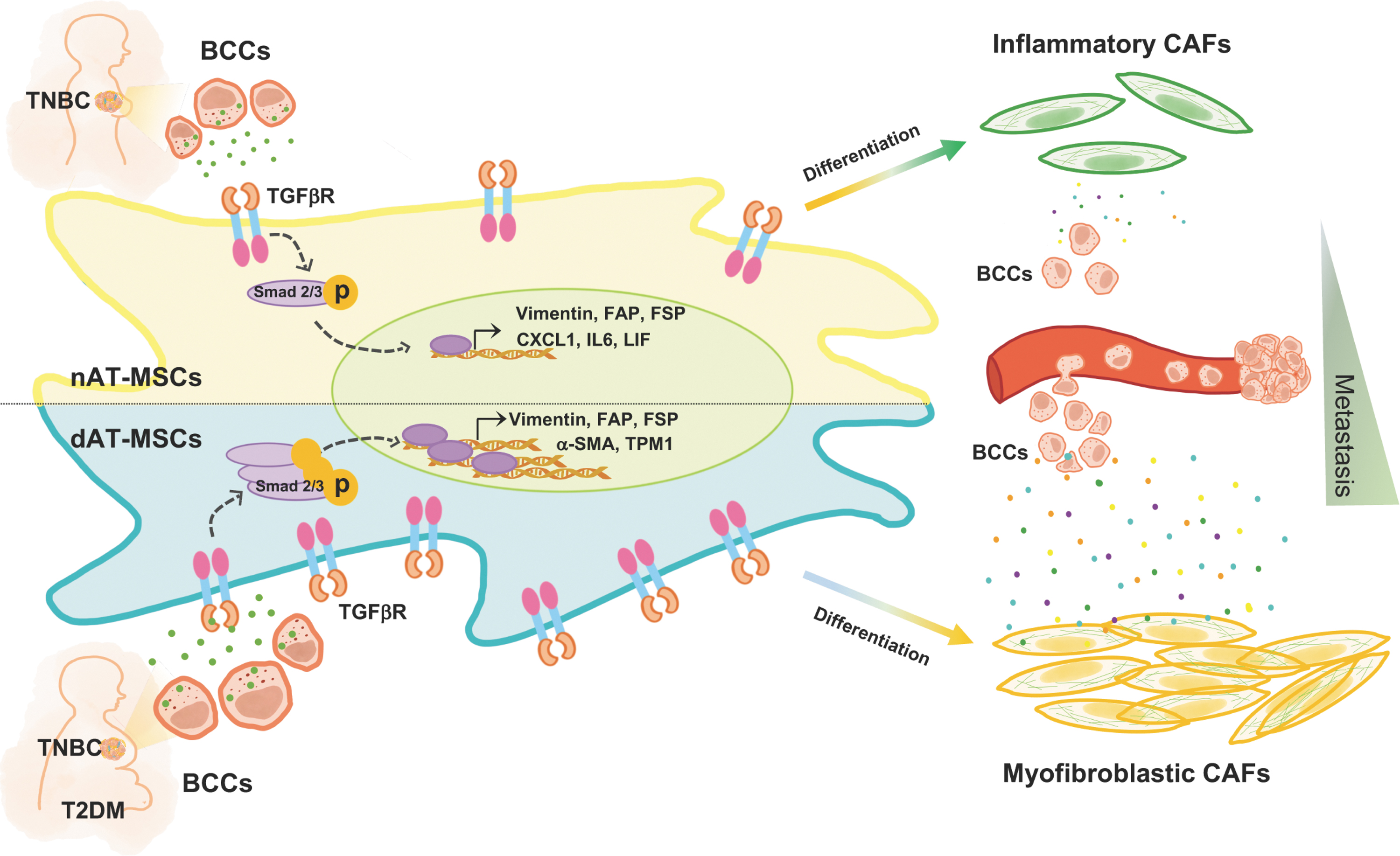
T2DM facilitates the differentiation of AT-MSCs into CAFs under hypoxic conditions by the activation of the TGFβ-Smad2/3 signaling pathway. In comparison with AT-MSCs, dAT-MSCs show a higher expression of TGFβR2 under hypoxic condition. Therefore, dAT-MSCs highly respond to the signaling from BCCs and promote the differentiation of dAT-MSCs into myofibroblastic CAFs, which support metastasis of BCCs. T2DM, type 2 diabetes mellitus.
The tumor microenvironment consists of different types of cells, in which MSCs are recognized to have a crucial role in promoting tumor growth and metastasis [34]. Given that breast tumors are mainly surrounded by adipose tissues, several studies have suggested that among the types of MSCs, AT-MSCs might be more involved in the stimulation of breast cancer progression [35,36].
Furthermore, our previous study reported that AT-MSCs enhance the migratory ability of BCCs in vitro and the metastatic ability of BCCs in vivo [24]. In addition to the direct effects of AT-MSCs on cancer cells, this study demonstrated that AT-MSCs also possess the ability to differentiate into CAFs in response to paracrine signaling from BCCs in coculture, which is consistent with a previous report [22], and that this differentiation was facilitated under hypoxic conditions.
In addition, we examined the CAFs differentiation of AT-MSCs derived from healthy donors at the younger age, average of 30 (yAT-MSCs). The results showed that yAT-MSCs express the cell surface markers of MSCs, such as positive with CD73, CD105, CD166 and negative with CD45 and CD14, while possessing the higher proliferation and differentiation abilities into adipocytes and osteoblast in comparison with AT-MSCs derived from elderly donors (Supplementary Fig. S2A–C). In addition, after being cocultured with BCCs for 14 days, the cells derived from yAT-MSCs showed a similar morphology with fibroblast and a significant upregulation of CAFs markers, such as vimentin, FAP, and FSP, as similar as AT-MSCs derived from elderly donors (Supplementary Fig. S2D, E).
CAF is the largest heterogeneous cell population in the tumor microenvironment [9], which induces the EMT and the metastasis of cancer cells [8,37]. Moreover, a study suggested that CAFs derived from AT-MSCs enhance the invasive ability of BCCs [22]. In comparison with MSCs, CAFs share many similarities, including the same surface markers of MSCs; the ability to differentiate into osteocytes, chondrocytes, and adipocytes; and the expression of cytoskeleton and extracellular matrix proteins [38,39]. Our results are consistent with previous reports, and demonstrate that both AT-MSCs and AT-CAFs showed a similar expression of MSC markers, such as positive with CD73, CD166, CD105, while negative with CD14 and CD45 (Supplementary Fig. 3A).
Furthermore, several reports identified α-SMA, vimentin, FAP, FSP as the traditional biomarkers of CAFs [28,37,40], which showed the higher expression in CAFs, in comparison with MSCs. Therefore, the upregulation of α-SMA, vimentin, FAP, and FSP was used to identify the generation of CAFs derived from MSCs in several studies [29,37]. Consistently, our data showed that, after being cocultured with BCCs for 14 days, cells derived from AT-MSCs exhibited the upregulation of vimentin, FAP, FSP (Fig. 1B, C), α-SMA, TPM1, CXCL1, IL6, and LIF (Fig. 2C–E), in comparison with those derived from AT-MSCs without coculturing with BCCs. Importantly, our findings also showed that AT-CAFs had the higher ability to induce the migration (Supplementary Fig. 3B) and in vivo metastasis of BCCs (Supplementary Fig. 3C), in comparison with AT-MSCs.
Previous reports have suggested that hypoxia facilitates the generation of CAFs from various progenitors, including fibroblasts, ECs, and MSCs [41], by inducing numerous signaling pathways, including the Wnt, PDGF, and TGFβ pathways [16,42]. Indeed, our results demonstrated that under hypoxic conditions, AT-MSCs showed the upregulation of the TGFβR2 expression.
In addition, AT-CAFs derived from AT-MSCs cocultured with BCCs under hypoxic conditions showed induced Smad2/3 phosphorylation and the higher expression of vimentin, a fibroblast marker, in comparison with normoxic conditions. As TGFβ is regarded as one of the key factors secreted by BCCs, which activates the differentiation of AT-MSCs into CAFs [22], our results suggested that hypoxic stimuli may induce the differentiation of AT-MSCs into CAFs, which might be involved in the activation of TGFβ signaling pathway.
Several studies indicate that T2DM is associated with worse outcomes in breast cancer patients [43 –47]. In addition, a study suggested that T2DM may alter the cellular behaviors of adipose tissue-derived cells, which promotes tumor growth and metastasis of breast cancer [35]. In this study, we showed that T2DM promotes the ability of dAT-MSCs to differentiate into CAFs under hypoxic condition. Moreover, our data suggested that T2DM affects the fate of CAFs, in which CAFs derived from AT-MSCs and dAT-MSCs belong to two different subpopulations; AT-CAFs are inflammatory CAFs that show the high expression of IL6, CXCL1, and LIF [28,48], while dAT-CAFs are myofibroblastic CAFs that show the high expression of a-SMA and TPM1 [28,40].
Inflammatory CAFs and myofibroblastic CAFs are two major subtypes of CAFs. Inflammatory CAFs support tumor development through the secretion of inflammatory factors [48]. Meanwhile, myofibroblastic CAFs locate adjacent to the tumor cells and have close contact with tumor cells, promoting metastasis and the EMT of tumor cells [28,40,49]. Of note, our findings suggested that dAT-CAFs exhibited an enhanced ability to induce proliferation, as well as the migratory and metastatic abilities of BCCs in comparison with nAT-CAFs.
These different functions might be because AT-CAFs and dAT-CAFs belong to different subpopulations. However, the signaling pathways involved in the induced effects of dAT-CAFs on TNBC are still not clear. A previous study determined that secreted factors derived from CAFs induce the EMT, and the migratory and invasive ability of BCCs, resulting in an increased risk of metastasis [8]. Therefore, a further study is needed to compare the growth factors secreted by dAT-CAFs and nAT-CAFs, and clarify the signaling pathways involved in the functions of these cells in TNBC metastasis.
The TGFβ signaling pathway plays an important role in promoting tumor progression of pancreatic cancer in diabetic patients [50]. In addition, several studies indicate that the generation of myofibroblastic CAFs is activated by TGFβ signaling pathway [49]. Consistent with these studies, our current results indicated that T2DM promoted the upregulation of the TGFβR2 expression in dAT-MSCs under hypoxic conditions.
Consequently, dAT-CAFs showed the higher phosphorylation of Smad2/3 in comparison with nAT-CAFs, which may give rise to the myofibroblastic CAFs fate. By adding TGFβR inhibitor to the coculture of dAT-MSCs and BCCs under hypoxic conditions, the CAFs differentiation ability of dAT-MSCs significantly declined. Even more noteworthy is the fact that in dATi-CAFs, derived from dAT-MSCs in coculture with BCCs in the presence of TGFβR2 inhibitor, the ability to support BCCs was reduced. These data suggested that targeting TGFβ-Smad2/3 signaling pathway to impair the CAFs differentiation of dAT-MSCs would be a promising candidate of anticancer therapy for DM patients with breast cancer.
Conclusion
Our study demonstrated that in comparison with AT-MSCs, dAT-MSCs possess the higher ability to differentiate into myofibroblastic CAFs under hypoxic conditions, which play a strong role in promoting the proliferation and metastasis of BCCs. Therefore, these findings suggested that T2DM might increase the risk of metastasis in BCCs by enhancing the generation of dAT-CAFs. Of note, impairment of the TGFβ-Smad2/3 signaling pathway in dAT-MSCs can be considered for the treatment of breast cancer of T2DM patients to reduce their risk of metastasis.
Acknowledgments
We thank the Japanese Ministry of Education, Culture, Sports, Science & Technology (MEXT) for their support.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.