
Abstract
Reserve mesenchyme cells (RMCs) are a type of antler stem cells (ASCs) that contribute to the rapid growth of deer antlers, the only known mammalian organ that can fully regenerate annually. Based on the prior evidence, ASC-conditioned medium could improve regenerative cutaneous healing in rats. The purpose of the study was to evaluate the therapeutic effects of RMC-conditioned medium (RMC-CM) on reducing the destruction in the mice periodontitis (PD) model and the underlying mechanisms. The lipopolysaccharide (LPS)-stimulated RAW264.7 cells were used in vitro to verify the effects of RMC-CM. The results revealed that RMC-CM could significantly reduce bone resorption and osteoclast activation, upregulate anti-inflammatory macrophages (M2) related interleukin (IL)-10 and CD206, and downregulate pro-inflammatory macrophages (M1) related tumor necrosis factor alpha (TNF-α) and inducible nitric oxide synthase in vivo. In vitro, RMC-CM could significantly promote LPS-stimulated RAW264.7 cells migration, reduce osteoclast differentiation, downregulate the expression of TNF-α, IL-6, and IL-1β, and upregulate the expression of IL-10 and arginase 1. According to the results, we concluded that RMC-CM could significantly reduce alveolar bone resorption and inhibit inflammation in gingival tissue by decreasing the activation of osteoclasts and inducing macrophage polarization toward the M2 phenotype. This study may serve as the experimental foundation for RMC-CM in the treatment of PD.
Introduction
Periodontitis (PD) is a chronic inflammatory disease that occurs in response to bacterial infection within the oral biofilm, which leads to the destruction of tooth-supporting tissues and is the main cause of tooth loss in adults [1]. The tissue destruction is mainly caused by the host immune response [2]; the infiltration of immune cells and activation of osteoclastic cells eventually lead to the periodontal destruction of both soft and hard tissues [3,4].
Macrophages are a type of immune cells that play a critical role in defense against periodontal pathogens [5]. Macrophages show plasticity [6,7]; M1 macrophages can induce alveolar bone loss and promote osteoclast differentiation in periodontal tissue [8]. Therefore, the factors that can transform macrophages into M1 or M2 are of great significance in the occurrence and development of periodontal disease.
The aim of periodontal regeneration is to restore the structure and function of the periodontal tissues. Stem cells are considered effective approaches for achieving true regeneration, compared with the available treatment options [9 –11]. Mesenchymal stem cells (MSCs) are a heterogeneous subset of stromal stem cells that can be isolated from various adult tissues and differentiate into many lineages [12 –15]. The crosstalk between MSCs and macrophages plays a key role in bone regeneration [16].
Reserve mesenchyme cells (RMCs) are a type of antler stem cells (ASCs), which contribute to the rapid growth of deer antlers owing to their remarkable proliferation potential. Deer antlers are the extraordinary and only known mammalian organs that can fully regenerate annually [17]. The RMCs express classic MSCs surface markers and have some properties of embryonic stem cells [18]. The RMCs are reported to have faster proliferation and increased osteogenic differentiation capacity relative to human MSCs under identical culture conditions [19].
However, transplanted MSCs fail to persist in vivo [20]. The “paracrine hypothesis” is believed to play a key role in their functions [21,22]. The secreted factors are broadly defined as conditioned medium (CM) [23]. Compared with stem cell-based therapies, CM have many advantages and confer similar therapeutic effects to MSCs in periodontal tissue regeneration [24 –27]. Further, RMC-CM has been reported to promote regenerative cutaneous healing in rats [28]. Therefore, this study was designed to investigate the ability of RMC-CM in reducing the destruction of periodontal tissues and the underlying mechanisms by using an experimental PD mouse model to provide a strategy for PD treatment.
Materials and Methods
Cell culture
The RMCs were kindly provided by Dr. Qianqian Guo based on a previous study [18]. Human bone marrow mesenchymal stem cells (hBMSCs) were purchased from Guangzhou Cellbank Biotech CO., Ltd. (Guangzhou, China). The RMCs and hBMSCs were cultured in α-minimum essential medium (α-MEM) (Gibco, NY, USA) containing 10% fetal bovine serum (FBS; BI, Israel) and 1% penicillin/streptomycin (P/S; Hyclone, USA). Cells at passages 4–6 were used for all experiments.
Mycoplasma detections of RMCs and hBMSCs were carried out using a mycoplasma detection kit (Solarbio, Beijing, China) according to the manufacturer's instructions. Briefly, cells were seeded at a density of 2 × 104 cells/well into 6-well plates; RMCs and hBMSCs cultured in α-MEM with 10% FBS and 1% P/S were used as control groups, whereas those cultured in α-MEM with 10% FBS and without P/S were used as experimental groups.
After culturing for 5 days, the supernatants were discarded, and the cells were fixed and incubated in Hoechst 33258 solution away from light. A fluorescence microscope (AMG Evos; Thermo Scientific, USA) was used for observation (Supplementary Fig. S1). Murine macrophage RAW 264.7 cell line was purchased from CCTCC (Wuhan, China) and cultured in DMEM (Gibco) containing 10% FBS and 1% P/S.
Preparation of CMs
The two types of CMs were prepared as previously described [28]. Briefly, when RMCs and hBMSCs reached 90% confluency, the normal medium was discarded and the cells were washed with phosphate-buffered saline (PBS; Gibco). After incubation in α-MEM (without FBS and P/S) for 48 h, the supernatants were harvested, centrifuged (1,000 rpm, 10 min at 4°C), filtered through 0.22 μm pore size filters (Millipore Corp., Billerica, MA, USA), and concentrated using ultrafiltration with a cutoff of 3 kDa (5,000 rpm, 40 min at 4°C; Millipore Corp.) for CM preparation.
The total protein concentrations in CMs were measured using the Enhanced BCA Protein Assay Kit (Beyotime, Shanghai, China), adjusted to 1 mg/mL, and stored at −80°C. The blank control (PBS) and negative control (α-MEM) were concentrated as described earlier.
Animal experiments
The study was approved by the Institutional Animal Care and Use Committee of Chinese Academy of Agricultural Sciences (CAAS) (Protocol No. 2021-001D). Specific pathogen-free male C57BL/6J mice (10–11-week-old) were purchased from Liaoning Changsheng Biotechnology Co., Ltd. (Liaoning, China). The sample sizes for the experimental groups were established based on a previous study (n = 32) [29]. The mice were housed in plastic cages (4–5 mice/cage), under a 12-h light-dark cycle with maintenance of a temperature of 24°C–26°C and a humidity of 40%–70% for 1 week.
During the experiments, efforts were made to reduce the pain and other unpleasant feelings of the animals; laboratory mice were kept in comfortable facilities and allowed to drink water and food. After mice acclimatized to the pathogen-free conditions, an experimental PD model was established according to a previous study [30]. The procedures of ligature placement, local injection, and euthanasia were conducted under isoflurane inhalation anesthesia (HFQ, China) supplied by the animal anesthesia system (R620-S1-IECS, RWD, China).
Briefly, mice were weighed and placed in a sealed container with 4% (v/v) isoflurane flow. After full anaesthetization, dual-knotted 5-0 silk ligatures were placed between the first and second right maxillary molars to initiate PD. Experimental PD mice were randomly divided into four groups (control, α-MEM, hBMSC-CM, and RMC-CM). The RMC-CM and hBMSC-CM were 10-fold concentrated and locally injected at a volume of 5 μL every 2 days [31], so as PBS and α-MEM. Mice in each group were sacrificed with euthanasia at 2 weeks. The collected gingival tissues around all three molars were stored at −80°C, and the maxillae were fixed in 4% paraformaldehyde (PFA). The entire experimental procedure of the present study is depicted (Fig. 1).

Schematic of the entire experimental procedure. RMCs, reserve mesenchyme cells; RMC-CM, RMC-conditioned medium
Micro-CT scanning
Maxillae were scanned using high-resolution Micro-CT (Scano Medical AG, Bassersdorf, Switzerland). Key parameters were set at 70 kV, 150 mA, and 10-μm increments, with an integration time of 300-ms. The scanned files were reconstructed and analyzed using Mimics Innovation Suite software (Materialise, Kingdom of Belgium). The distance between the cementoenamel junction and the alveolar bone crest of the distobuccal root of first molars and the mesiobuccal root of second molars in 3D images was measured as previously described [29,30]. The measurements were repeated twice by two independent pathologists who were blinded to the group allocation using ImageJ software (National Institutes of Health, USA), and the mean distances were obtained in millimeters.
Histology and immunohistochemistry staining
The specimens were decalcified in 10% Ethylene Diamine Tetraacetic Acid solution (Beijing Chemical Works, Beijing, China) for 2 weeks at room temperature. After being embedded and sliced, the sections were stained with hematoxylin-eosin (H&E) (Servicebio, China), tartrate-resistant acid phosphatase (TRAP) (Jiancheng Technology, Nanjing, China), and immunohistochemistry (IHC). TRAP-positive multinucleated (>3 nuclei) cells were considered osteoclasts [32].
An antibody against CD68 (Proteintech, Wuhan, China) was used. The sections were scanned using an Olympus BX53 microscope (Olympus Corporation, Japan) and browsed using CaseViewer software (3DHISTECH Ltd., Budapest, Hungary). The number of TRAP and CD68 positive cells was counted by two independent pathologists who were blinded to the group allocation using ImageJ software.
Cell Counting Kit-8 assay
The viability of RAW264.7 was evaluated using the Cell Counting Kit-8 (CCK-8) assay (Solarbio), according to the manufacturer's instructions. RAW 264.7 cells were seeded at 3,000 cells/well into a 96-well plate. Except for the control wells, all cells were treated with Porphyromonas gingivalis-Lipopolysaccharide (LPS) (1 μg/mL; InvivoGen, San Diego, CA, USA) for 12 h; then, α-MEM (without FBS and P/S) and a double dilution series of RMC-CM were added to the control wells and other wells for 8 h of further incubation.
Measurement was performed at 450 nm using a microplate reader (Synergy HT; Biotek, USA), and the results are expressed as the ratio of the wells compared with control wells.
Enzyme-linked immunosorbent assay
RAW 264.7 cells were seeded at 3 × 105 cells/well in a 6-well plate. After treating with P. gingivalis-LPS (1 μg/mL) overnight, RMC-CM (0.25 mg/mL), hBMSC-CM (0.25 mg/mL), and α-MEM were added to the wells separately for 24 h. Supernatants were collected to detect cytokine levels of tumor necrosis factor alpha (TNF-α) and interleukin (IL)-10 using enzyme-linked immunosorbent assay (ELISA) kits (Cloud-Clone Corp., Wuhan, China) according to the instructions. RAW 264.7 cells were used for RNA extraction.
Flow cytometry
The ratio of M2/M1 macrophage was determined using flow cytometry [33]. Briefly, RAW 264.7 cells were seeded at 5 × 106 cells/well in a 6-well plate. After stimulation with P. gingivalis-LPS (1 μg/mL) for overnight, the cultured medium was changed to DMEM separately with RMC-CM, hBMSC-CM, and α-MEM for 24 h. The cell suspension was separately divided into 1.5 mL EP tubes and incubated with blocking antibody CD16/32 (Proteintech) at 4°C for 1 h.
After washing twice with PBS, RAW264.7 were incubated in PE anti-mouse CD206 antibody (surface marker of M2; Biolegend, San Diego, CA, USA) and APC anti-mouse CD86 (surface marker of M1; Proteintech) at 4°C for 30 min. After washing twice with PBS, cells were suspended in 300 μL PBS and detected using flow cytometry (BD FACSCalibur, Shanghai, China).
Transwell analysis
Transwell dishes with 8.0 μm pore filters (Corning Incorporated, NY, USA) were used as previously described [34]. Briefly, after treating with P. gingivalis-LPS (1 μg/mL) overnight, 2 × 105 RAW264.7 cells in 300 μL of DMEM with 0.5% FBS and 1% PS were seeded into the upper chamber. RMC-CM, hBMSC-CM, and α-MEM were separately added to the lower chambers. After further incubation for 24 h, cells on the transwell filters were fixed in 4% (v/v) PFA and stained with crystal violet solution (Beyotime).
The cells on the upper surface of the filters were removed using cotton swabs and counted at 200 × magnification. The measurements were repeated twice by two independent pathologists who were blinded to the group allocation using ImageJ software.
Osteoclastogenesis induction
RAW 264.7 cells were seeded at 6,000 cells/well in a 96-well plate [35], treated with 30 ng/mL RANKL (Novoprotein, Shanghai, China) for 4 h and P. gingivalis-LPS (10 ng/mL) overnight. After rinsing with PBS, RANKL with RMC-CM, hBMSC-CM, and α-MEM were separately added to the wells. The mediums were changed daily until osteoclast formation. Osteoclasts were stained using TRAP staining kits (Cosmo Bio, Japan) according to the manufacturer's instructions. The measurements were repeated twice by two independent pathologists who were blinded to the group allocation using ImageJ software.
Quantitative real-time PCR
Total RNA was extracted from gingival tissues and RAW264.7 cells using TRIeasyTM Total RNA Extraction Reagent (Yeasen, Shanghai, China), and cDNA was synthesized with Hifair® II 1st Strand cDNA Synthesis SuperMix for qPCR (Yeasen). qPCR was performed using Hifair qPCR SYBR Green Master Mix (Low Rox Plus; Yeasen). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) primer was purchased from Sangon Biotech (Shanghai, China). The primer sequences are listed (Table 1).
Primer Sequences
Arg-1, arginase 1; Catk, cathepsin K; Il, interleukin; inos, inducible nitric oxide synthase; Nfatc1, nuclear factor of activated T cells c1; Tnfα, tumor necrosis factor alpha; Trap, tartrate-resistant acid phosphatase.
Western blot assay
RAW264.7 were collected and lysed by radioimmunoprecipitation assay (Solarbio) on ice for 10 min. After decomposing with ultrasonic, the mixture was centrifugated at 16,000 rpm at 4°C for 30 min. Then, the supernatant was collected and the total protein concentration was determined by BCA protein quantification kit (Beyotime). After denaturation with 5 × loading buffer (Beyotime) at 100°C for 10 min, 20 μg protein samples were separated in 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (Beyotime) and then transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA).
Membranes were soaked in 5% non-fat milk for 1 h at room temperature; then, they were covered with the primary antibodies against nuclear factor of activated T cells c1 (NFATc1) (1:1,000; Affinity, Jiangsu, China), c-Fos (1:10,000; Proteintech), cathepsin K (CatK) (1:1,000; Affinity, Jiangsu, China), TRAP (1:1,000; Proteintech), and GAPDH (1:10,000; Engibody, DE, USA) overnight at 4°C and incubated with anti-mouse (1:10,000; Proteintech) or anti-rabbit secondary antibodies (1:2,000; Yeasen) for 1 h at room temperature on shaker. The protein bands were visualized with NcmECL Ultra (NCM, Suzhou, China), and the protein expression was analyzed by Image J software. The protein levels were valued using GAPDH as a loading control.
Statistical analysis
Data are expressed as mean ± standard deviation. Comparisons between groups were performed using One-way analysis of variance (ANOVA) of variance with a post hoc test. Differences were considered significant at P < 0.05. All statistical analyses were performed using the Prism software (GraphPad, CA, USA).
Results
RMC-CM inhibits alveolar bone loss
First, we verified the effect of RMC-CM on alveolar bone loss. By comparing the results of micro-CT, we found that RMC-CM significantly reduced alveolar bone loss compared with control and α-MEM groups, with no significant differences between RMC-CM and hBMSC-CM groups (Fig. 2A, C). The trends of result from sections stained with H&E were consistent with that of micro-CT (Fig. 2B, D). The results of RMC-CM biosafety in vivo showed there were no histological abnormity in the thymus, liver, spleen, and kidney (Supplementary Fig. S2), indicating that RMC-CM is safe for application in mice.

Effect of RMC-CM on the reduction of alveolar bone loss in an experimental PD model.
RMC-CM inhibits osteoclast activation
To explore the mechanism of reduction of alveolar bone resorption, we stained the sections with TRAP for osteoclast activation analysis. The results demonstrated that osteoclast number was significantly lower in RMC-CM group than control group, with no significant differences found between RMC-CM and hBMSC-CM groups (Fig. 3A, C). The IHC staining against CD68 verified the result of osteoclast activation (Fig. 3B, D). Overall, osteoclast activation displays similar trends with alveolar bone loss during the development of PD.
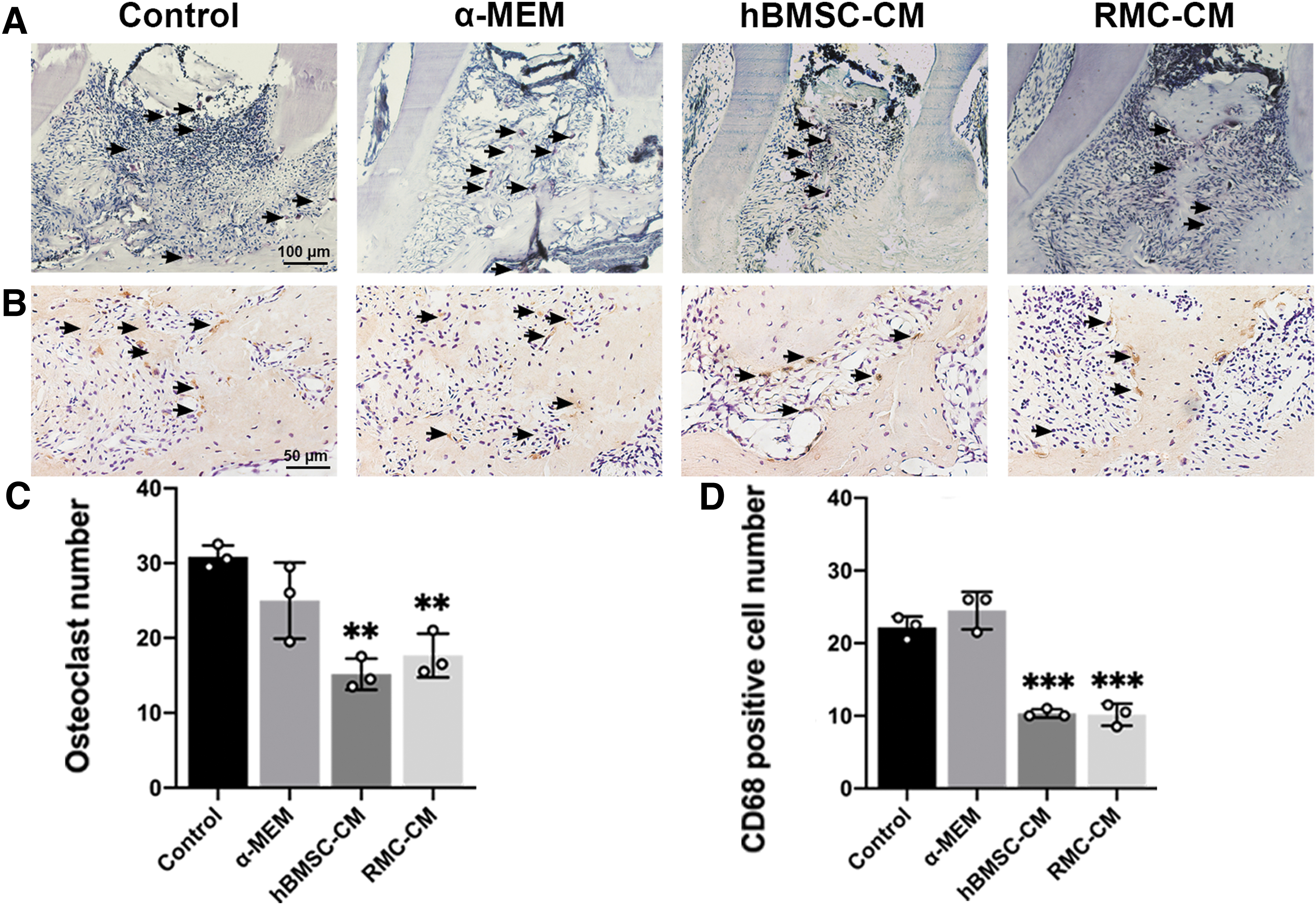
Effect of RMC-CM on osteoclast activation.
RMC-CM attenuates the level of gingival tissue inflammation
Next, we explored the effect of RMC-CM on inflammation in the gingival tissue. The results revealed that the expression of TNF-α and inducible nitric oxide synthase (iNOS) in RMC-CM group was significantly lower than the control group (Fig. 4A, C), and the expression of IL-10 and CD206 in RMC-CM group was significantly higher than the control group (Fig. 4B, D), with no significant difference between hBMSC-CM and RMC-CM groups.
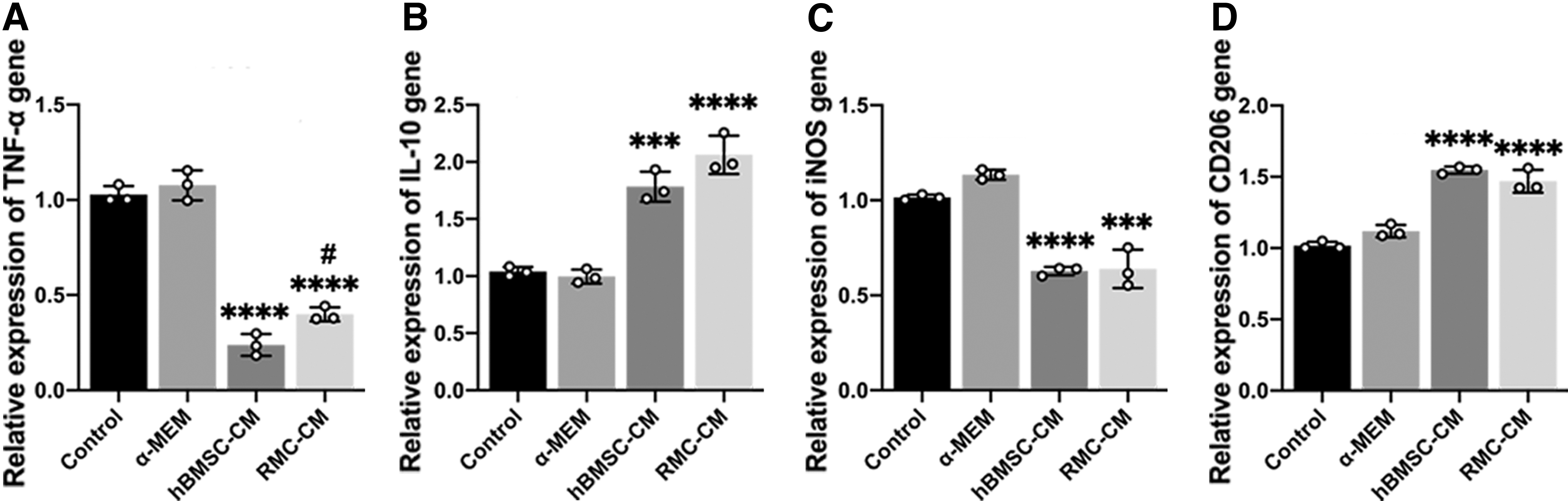
Effect of RMC-CM on inflammation-related mRNA expression in an experimental PD model.
TNF-α and iNOS are pro-inflammatory cytokines released by M1 macrophages, IL-10 is an anti-inflammatory cytokine released by M2 macrophages, and CD206 is a marker related to M2 macrophages. These results suggest that RMC-CM may attenuate the level of gingival tissue inflammation by regulating the proportion of M1 and M2.
RMC-CM downregulates pro-inflammation and upregulates the anti-inflammatory response in LPS-stimulated RAW 264.7 cells
We used LPS-stimulated RAW 264.7 in vitro to imitate inflammation. The result of cell viability showed that, at a concentration of 0.25 mg/mL, RMC-CM could reduce the negative effect of P. gingivalis-LPS on RAW264.7 (Fig. 5A). Next, we tested TNF-α and IL-10 produced from LPS-stimulated RAW264.7 cells using ELISA. The concentration of TNF-α and IL-10 was found to be significantly lower and higher in the RMC-CM group than in the α-MEM group, with no significant difference between the RMC-CM and hBMSC-CM groups (Fig. 5B, C).
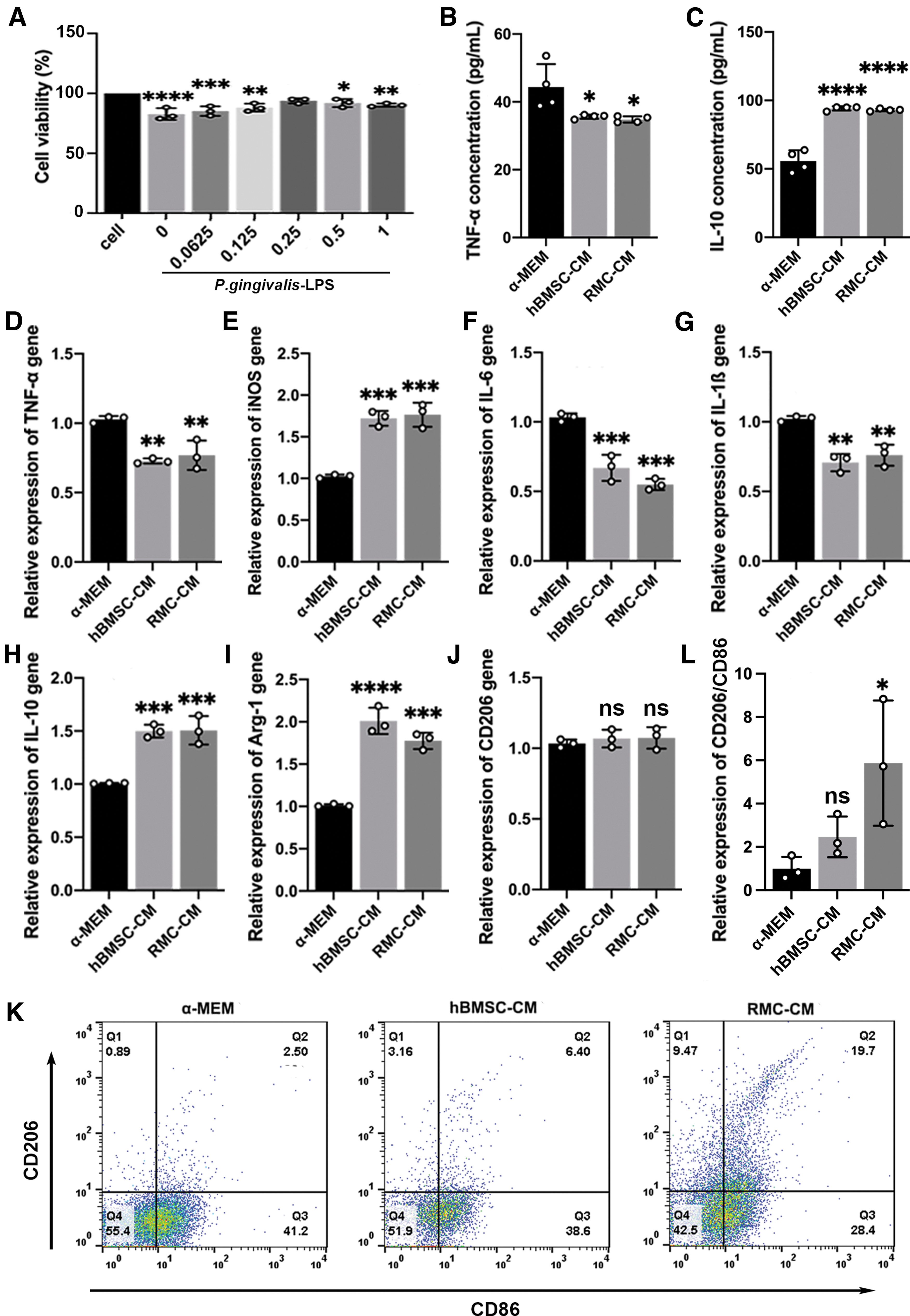
Effects of RMC-CM on LPS-stimulated RAW 264.7 cells.
The mRNA expression of TNF-α and IL-10 was consistent with the in vivo findings (Fig. 5D, H). The mRNA expression of IL-6 and IL-1β was significantly lower and the expression of arginase 1 (Arg-1) was significantly higher in RMC-CM group than α-MEM group, with no significant difference between RMC-CM and hBMSC-CM groups (Fig. 5F, G, I). However, the expression of iNOS was significantly higher in the RMC-CM group than in the α-MEM group, with no significant difference found between the RMC-CM and hBMSC-CM groups (Fig. 5E), and the expression of CD206 in RMC-CM group showed no difference among the three groups (Fig. 5J).
To further verify the effect of RMC-CM on macrophage polarization, we tested the ratio of M2/M1 using flow cytometry. CD206 and CD86 are separately the special surface phenotype marker of M2 and M1. The results showed that the ratio of CD206/CD86 in RMC-CM group was significantly higher than that in other groups (Fig. 5K, L). Because M1 macrophages are characterized by a high pro-inflammatory cytokine production (such as TNF-α, IL-1β and IL-6), and M2 macrophages are mainly characterized by high IL-10 and Arg-1 production, and low TNF-α and IL-1β production [36], the underlying mechanism of the effects of RMC-CM in LPS-stimulated RAW 264.7 cells may be related to the upregulation of the M2 polarization phenotype.
RMC-CM inhibits osteoclastogenesis in LPS-stimulated RAW 264.7 cells
To verify the effect of RMC-CM on osteoclast activation, we first investigated the effect of RMC-CM on the migration of LPS-stimulated RAW264.7 in vitro using a Transwell assay. The result showed that RMC-CM could significantly promote migration of LPS-stimulated RAW 264.7 compared with α-MEM and hBMSC-CM groups (Fig. 6A, D). After osteoclastogenesis induction, the area (Fig. 6B, E) and number (Fig. 6C, F) of TRAP-positive multinucleated cells in RMC-CM group were significantly less than those in the other two groups.

Effects of RMC-CM on osteoclast activation.
To verify whether the TRAP-positive multinucleated cells were osteoclasts, the expressions of osteoclast-related genes were determined using qPCR and were found to represent osteoclast-related markers (Fig. 6G–J). c-Fos and NFATc1 are involved in early osteoclast differentiation, whereas TRAP and CatK are involved in terminal osteoclast differentiation [37]. Western blot assay was used to further determine the protein production level of these osteoclast differentiation-related markers (Fig. 6K–O). Overall, the results indicated that RMC-CM could significantly inhibit osteoclast activation in vitro.
Discussion
The objective of the current study was to determine the potential of RMC-CM in PD treatment. First, we demonstrated that RMC-CM reduced alveolar bone resorption and osteoclast activation in an experimental PD model, and RMC-CM may attenuate the level of gingival tissue inflammation by upregulating the proportion of M2 macrophages. The effects of RMC-CM in LPS-stimulated RAW 264.7 cells in vitro also verified the results from in vivo.
The RMCs are a type of ASCs that are the initiating factors for the periodic complete regeneration, and deer antler is the only mammalian organ known to regenerate integrally every year [38]. Compared with other types of mammalian stem cells, RMCs are easier to obtain and exhibit higher proliferation and regeneration capacity [18,28,39]. Further, they play a key role through paracrine effect [40]. Therefore, the study of RMCs will help to provide new strategies for the treatment of PD.
In the present study, hBMSCs were selected as a positive control for the following reasons: (1) Both RMC-CM and hBMSC-CM are cross-species CM to mice, so the comparison results are relatively convincing; (2) hBMSC-CM has been used in human clinical studies of alveolar bone regeneration and has shown remarkable osteogenic potential in regenerative medicine [41 –43]. Although the most effect of RMC-CM was similar to hBMSC-CM, it was sufficient to prove the effectiveness and application potential of RMC-CM.
In addition, RMCs are much easier to obtain compared with hBMSCs and could overcome disadvantages of hBMSCs [44]. In our study, we have not found immunological rejection or side effects of RMC-CM injection on C57/BL6 mice. First, there is no inflammation reaction such as redness, swelling, or ulceration around the injection point. Second, there were no morphological and histological changes in the vital organs through weighing organs and performing histological staining. Third, the research of RMC-CM in other fields has been reported and no immunological rejection or side effect has been reported [28,39,45]. The earlier results prove the safety of RMC-CM.
The effect of RMC-CM on inflammation regulation was also determined in this study. Macrophages are important in innate immunity and are not homogenous [46,47]. M1 macrophages are characterized by high production of IL-1β, IL-6, IL-12, and IL-23. M2 macrophages are mainly characterized by high production of IL-10, transforming growth factor-β, and vascular endothelial growth factor, and low production of IL-12, TNF-α, and IL-1β [36]. In addition, iNOS is the marker of M1 macrophages [48]; CD206 and Arg-1 are the markers of M2 macrophages [49].
In our study, the results of qPCR and flow cytometry suggest that RMC-CM might play a key role in upregulating the M2 polarization phenotype. However, the expression of CD206 and iNOS in vitro was not consistent with that of in vivo. First, CD206 was reported to express in a great quantity on day 21 in vivo [50]. However, since our study mainly focused on the effect of RMC-CM on the early stage of inflammation, so LPS-stimulated RAW264.7 cells were treated with RMC-CM in vitro for only 24 h.
It may be the reason why there is no statistical significance between the groups. Second, the upregulated expression of iNOS in vitro was also noted. It has been reported that iNOS is not only the marker of M1 macrophages, but it also plays a key role in the mechanism of MSC-mediated immunosuppression [51]. The upregulation of iNOS in vitro might be related to the immunosuppressive activity of RMC-CM.
Our study also revealed that RMC-CM could inhibit osteoclastogenesis. It is worth noting that, though there was no significance in gene expression level among control, hBMSC-CM, and RMC-CM groups, the protein expression level of TRAP was significantly lower than that in other groups. The reason may be explained that TRAP activity is based not only on the gene expression but also on reactive oxygen species (ROS) [52].
We are currently comparing the components of RMC-CM and hBMSC-CM and have found that the specific component in RMC-CM can eliminate ROS, and this part of the results has not been published. We speculate that RMC-CM inhibits the activity of TRAP by scavenging ROS without affecting the expression of TRAP gene.
The results of the present study indicated RMC-CM as a promising alternative strategy for PD treatment in regenerative medicine, but it still faces many problems in the application of RMC-CM. To solve the problems, our current work is to analyze and purify specific components of RMC-CM; to apply RMCs more scientifically and effectively in the treatment of PD.
Conclusions
In summary, RMC-CM could significantly reduce alveolar bone resorption and gingival tissue inflammation in mice with experimental PD by decreasing the activation of osteoclasts and altering the proportion of M1 and M2. Our findings suggest the potential of RMC-CM for PD treatment.
Footnotes
Acknowledgments
The authors are grateful for helpful comments from Zhen Liu, Lijing Wang, and Janak Lal Pathak.
Author Disclosure statement
No competing financial interests exist.
Funding Information
This research was supported by grants from the Ministry of Science and Technology (No. 2021YFE0108000), Medical Support Program of the Jilin University (No. 20170311032YY), Science and Technology Project of the Jilin Provincial Department of Finance (Nos. jcsz2020304-9 and jsz2018170-12), and Traditional Chinese Medicine Bureau of Guangdong Province (No. 20222131). The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the article apart from those disclosed. No writing assistance was utilized in the production of this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.