
Abstract
Due to their immunosuppressive potential and ability to differentiate into multiple musculoskeletal cell lineages, mesenchymal stromal cells (MSCs) became popular in clinical trials for the treatment of musculoskeletal disorders. The aim of this study was to isolate and characterize native populations of MSCs from human cortical and cancellous bone from the posterior elements of the lumbar spine and determine what source of MSCs yields better quality and quantity of cells to be potentially used for spinal fusion repair. We were able to show that MSCs from trabecular and cortical spine had the typical MSC morphology and expression markers; the ability to differentiate in adipocyte, chondrocyte, or osteoblast but they did not have a consistent pattern in the expression of the specific differentiation lineage genes. Moreover, MSCs from both sites demonstrated an immune suppression profile suggesting that these cells may have a more promising success in applications related to immunomodulation more than exploring their ability to drive osteogenesis to prevent nonunion in spine fusion procedures.
Introduction
Autologous bone grafts have been widely used in orthopedics and are considered the gold standard for spine fusion surgeries to achieve spinal fusion. However, autologous bone grafts are associated with significant disadvantages and complications such as rejection, increased operative time, insufficient availability, infection, and chronic pain at the donor site [1,2]. Due to limitations of current treatment options, over the past several decades there has been a dramatic increase in the use of cell-based therapies in orthopedics [3 –5]. Various bone substitutes have been developed to promote spinal fusion; adult mesenchymal stromal cells (MSCs) have become more popular in cell-based therapy for the treatment of orthopedic pathologies and for the repair and regeneration of bones [6 –10].
MSCs represent a heterogeneous cell population capable of differentiating into multiple musculoskeletal cell types, releasing a plethora of growth factors and cytokines, in addition to having the ability to reduce the immune response. Because of these properties, MSCs are considered a suitable tool for regenerative medicine and have already been introduced in a number of clinical trials for tissue repair [11 –15]. Moreover, these properties make MSCs particularly desirable for clinical applications in the field of orthopedics, including in spine fusion procedures [16 –20].
Due to ability of MSCs to facilitate tissue regeneration, there has been considerable clinical interest in the use of MSCs obtained from bone marrow for osseous graft supplementation therapy [16 –18,21,22]. Optimization of extraction, culture, and differentiation methods have allowed MSCs to be suitable to clinical applications [23,24]. In addition, several groups have been investigating the potential efficacy of MSCs as a cell-based therapy for orthopedic tissue applications. However, despite pre-clinical trials being conducted, commercially available products for MSC-based cell therapy have yet to be approved and used in the clinic [11 –15,25 –28].
The lag of MSCs being used in the clinic is partially due to the fact that MSCs derived from different tissues share similar epitope profiles, but show significant differences in the differentiation, proliferation, and migration potential. Multiple groups have demonstrated that properties of MSCs are significantly affected by the tissue of origination because MSCs receive various signals that directly affect the differentiation potential and immunosuppressive properties [29 –31]. For example, adipose tissue is considered a reliable source of MSCs [32,33], that can be obtained through a minimally invasive method, lipoaspiration. In addition, lipoaspiration yields a greater quantity of tissue, and thus a greater number of cells, compared to other methods [30,31,34,35]. However, most articles have described the inferior osteogenic potential of MSCs derived from adipose tissue compared to MSCs derived from bone marrow [36].
Several groups have analyzed the cellular and molecular characteristics of human adult MSCs derived from different sites within the human skeletal including bone marrow from iliac crest (Ic-MSCs), sternum (St-MSCs), vertebrae (vMSCs), colon (Co-MSCs), dental pulp (DPSCs). Sites throughout the bone are reservoirs of multipotent cells [37,38], but the trabecular [39 –41] and cortical [42,43] components are the most commonly used reservoirs for MSC extraction. Other investigations have shown that vertebral body (VB) bone marrow contains stem cells, and MSCs from the VB bone marrow are able to initiate osteogenesis better than MSCs isolated from the iliac crest (IC bone marrow) [44,45].
Since several studied have demonstrated that vMSCs have potentially superior osteogenic properties compared to MSCs from other sites [44 –46], the purpose of this study was to isolate and characterize native populations of MSCs from the human cortical and cancellous bone from the posterior elements of the lumbar spine. Characteristics investigated included (1) comparing the quantity of MSCs from cortical and cancellous bone, (2) characterizing MSCs stem cell marker expression, and (3) determining the differentiation potential and immunomodulatory potency of MSCs from the two different reservoirs. By evaluating these properties, we aimed to determine whether cortical or cancellous bone within the posterior lumbar spine yielded a superior quality and quantity of MSCs to potentially use MSCs in spinal fusion repair.
Materials and Methods
Sample acquisition
IRB approval for the collection of patient data, lumbar spine patient samples and human bone marrow samples were obtained at Houston Methodist Hospital (IRB Pro00015718) and from the orthopedic biorepository at Houston Methodist Hospital Orthopedics and Sports Medicine Department (IRB CR00006624) respectively. All patients were consented before sample collection and sample processing. The samples were deidentified and individually and anonymously bio-banked with specific identification numbers.
Cell isolation protocol
Lumbar spine MSC populations were extracted by processing the posterior elements of four patients undergoing laminectomy following a previously described protocol [47] with some minor modifications due to the use of human samples. The extraction protocol was approved by the Institutional Review Board of the Houston Methodist Hospital (Houston, TX, USA).
Bone fragments were obtained following the routine dissection and extraction of bone as part of standard of care for laminectomy surgery. No modification to the original surgery was made in the removal of bone fragments. Bone fragments used would have otherwise been discarded and no excess tissue was obtained. Bones were cleaned of any fat, connective tissue, or muscle by sharp dissection. Samples were washed with phosphate-buffered saline (PBS) (Thermo Fisher Scientific, Waltham, MA) supplemented with 1% antibiotic/antimycotic (Gibco, Grand Island, NY) until they appeared clean of all debris to remove any blood. The cancellous and trabecular portions of the bone were sharply transected and separated. The two bone fragments were crushed into chips ∼3–4 mm3 in size and transferred to 50-mL polypropylene tubes (BD Falcon, Bedford, MA).
Bone chips were suspended in alpha-modified Eagle's medium (α-MEM; Sigma-Aldrich, St. Louis, MO) containing 2% (vol/vol) defined fetal bovine serum (FBS) (Gibco) with 3 mg/mL collagenase type-I (Worthington Biochemical Corporation, Lakewood, NJ) and 4 mg/mL dispase II (Roche, Indianapolis, IN) and placed on a shaking platform at 37°C for 3 h. After digestion, bone chips were plated in new flasks for 3 days to allow cells to migrate out of bone fragments. These cells isolated were considered passage 0 (P0).
Bone marrow MSCs (BM-MSCs) were instead isolated from 10 mL human bone marrow obtained from patients. Briefly, bone marrow was washed with PBS solution and centrifugated. The pellet was resuspended in 10 mL α-MEM (Sigma-Aldrich) containing 20% (vol/vol) FBS (Gibco) supplemented with 1% penicillin (100 UI/mL)-streptomycin (100 mg/mL) at a seeding of 0.26 mL per cm2, these cells were considered passage 0 (P0).
Human lumbar spine and BM-MSCs expansion
Adherent cells were cultured in α-MEM (Sigma-Aldrich) containing 20% (vol/vol) FBS supplement and 1% penicillin (100 UI/mL)-streptomycin (100 mg/mL) (Gibco) and incubated at 37°C in low oxygen conditions. Media was changed every 48 h until the cells were at 80% confluency at which point, they were passaged. Cells were used for experiments until P5.
Cell doubling time
Proliferation rate was determined in biological triplicates from P1 to P5 as previously reported [48,49]. Cells were seeded into a 24-well tissue culture polystyrene plate. At 90% confluence, cells were trypsinized, counted, and plated again at the same density. The mean of population doublings was calculated at each passage according to the formula CD = log (Nc/No)/log2 and PD = CT/CD, where CD represents cell doubling, Nc represents the number of cells at confluence, No represents seeded cells and CT represents the culture time. Data obtained from each cell line were reported as a mean of the values.
Flow cytometry analysis
Lumbar spine MSCs were collected and characterized for the expression of their associated surface cell markers. Briefly, cells were washed with FACS buffer and stained for 30 min at 4°C with negative cocktail markers (human leukocyte antigen– DR isotype HLA-DR, CD45, CD11b, CD19, and CD34) and positive anti-human cocktail markers (cluster differentiation CD90, CD73, CD105). Conjugated primary monoclonal antibodies and isotype controls were used as recommended by the manufacturer (BD Biosciences). Cells were analyzed on a FACS Fortessa flow cytometer (BD Biosciences) and analyzed using FCS Express (Denovo Software).
Multilineage differentiation
Multilineage cell potential toward osteogenic, adipogenic, and chondrogenic differentiation was assessed in vitro at P3. Lumbar spine MSCs were seeded at the density of 5,000 cells/cm2 in 12-well plates and cultured until ∼80–90% confluence. Then the media was replaced according to the differentiation assay using StemPro Chondrogenesis, Osteogenesis, or Adipogenesis Differentiation Kit (all from Gibco) for Chondrogenic, Osteogenic, or Adipogenic induction, respectively.
After 21 days, the multilineage potential was evaluated using different staining protocols. For chondrogenic differentiation conventional Alcian Blue staining (Sigma) was performed to highlight proteoglycan deposition. Osteogenic induction was confirmed by evaluating mineral deposition using von Kossa staining (Von Kossa Stain Kit American MasterTech) and confirmed by evaluating Ca2+ deposits using Alizarin Red staining (Sigma Aldrich). For adipogenic induction, intracellular lipid droplets accumulation was visualized by Oil red O staining (Oil Red O Stain Kit American MasterTech). hBM-MSCs cultured in differentiation medium and hPLS MSC cultured in normal medium for the same time were used as positive and negative control respectively.
Immunosuppressive potential stimulation
For in vitro cell stimulation, cells were seeded at a density of 5,000 cells/cm2 in 24-well plates in standard supplemented medium, α-MEM supplemented with 20% FBS and 1% penicillin (100 UI/mL) streptomycin (100 mg/mL). After 24 h, cells were treated with 100 ng/mL of soluble recombinant human TNFα and IFNγ (Preprotech) and at different time points (6, 12, 24, 48, and 72 h) cells were harvested for flow cytometry phenotyping and RNA analysis.
Gene expression analysis
Total RNA was isolated using 0.5 mL of TRIzol reagent (Life Technologies, ThermoFisher Scientific). For each sample, RNA concentration and purity were measured using a NanoDrop spectrophotometer (ND1000; NanoDrop, ThermoFisher Scientific). cDNA was synthesized from 1 μg of total RNA using a TaqMan Reverse Transcription reagent kit (Applied Biosystems, ThermoFisher Scientific). Amplifications were set on plates in a final volume of 10 μL and carried out using TaqMan Fast Advanced MasterMix (Applied Biosystems, ThermoFisher Scientific). The housekeeping marker included in the study was eukaryotic 18S rRNA (18S; Hs03003631_g1).
Specific lineage associated markers
For chondrogenesis gene expression the following markers were included: aggrecan (ACAN; Hs00153936_m1, SRY related high-mobility group box transcription factor (SOX9; Hs01001343_g1), type 1 collagen (COL1a; Hs00164004_m1), and type 2 collagen (COL2A1; Hs00264051_m1). For osteogenesis gene expression: osteocalcin (BGLAP; Hs01587814_g1), osteonectin (SPARC; Hs00234160_m1), alkaline phosphatase (ALP; Hs01029144_m1), osteopontin (SSP1; Hs00959010_m1), and Runt-related transcription factor 2 (RUNX2: Hs00231692_m1). For adipogenesis expression: leptin (LEP; Hs00174877_m1), lipoprotein lipase (LPL; Hs00173425_m1), adiponectin (ADIPOQ; Hs00605917_m1), and peroxisome proliferator-activated receptor gamma (PPARγ; Hs0115513_m1).
Immunosuppression associated markers
Prostaglandin E synthase (PTGES; Hs01115610_m1), prostaglandin-endoperoxide synthase 2 (PTGS2; Hs00153133_m1), and indoleamine 2,3-dioxygenase 1 (IDO1; Hs00984148_m1).
Statistical analysis
Statistical analysis was performed using GraphPad for Windows (GraphPad Software). Three replicates for each experiment were performed, and the results are reported as mean ± SD, with P ≤ 0.05 used as a threshold for significance. Statistical analysis by unpaired t-test with Welch's correction was used to compare differences between groups.
Results
MSC isolation and characterization
Human cortical and trabecular MSCs were isolated from human posterior lumbar spine (hPLS) specimens collected at Houston Methodist Hospital in the Department of Orthopedics and Sports Medicine (IRB Pro00015718). Three patients were female with the average age of 67 years (range 61–76, SD 6.6) and one patient was a 40-year-old male (Table 1). Both cortical and trabecular MSCs isolated from hPLS had the fibroblast-like shape resembling MSC morphology after cell isolation and expansion (Fig. 1A). No significant differences were observed in the doubling times between passage 1 and 5 (Fig. 1B).
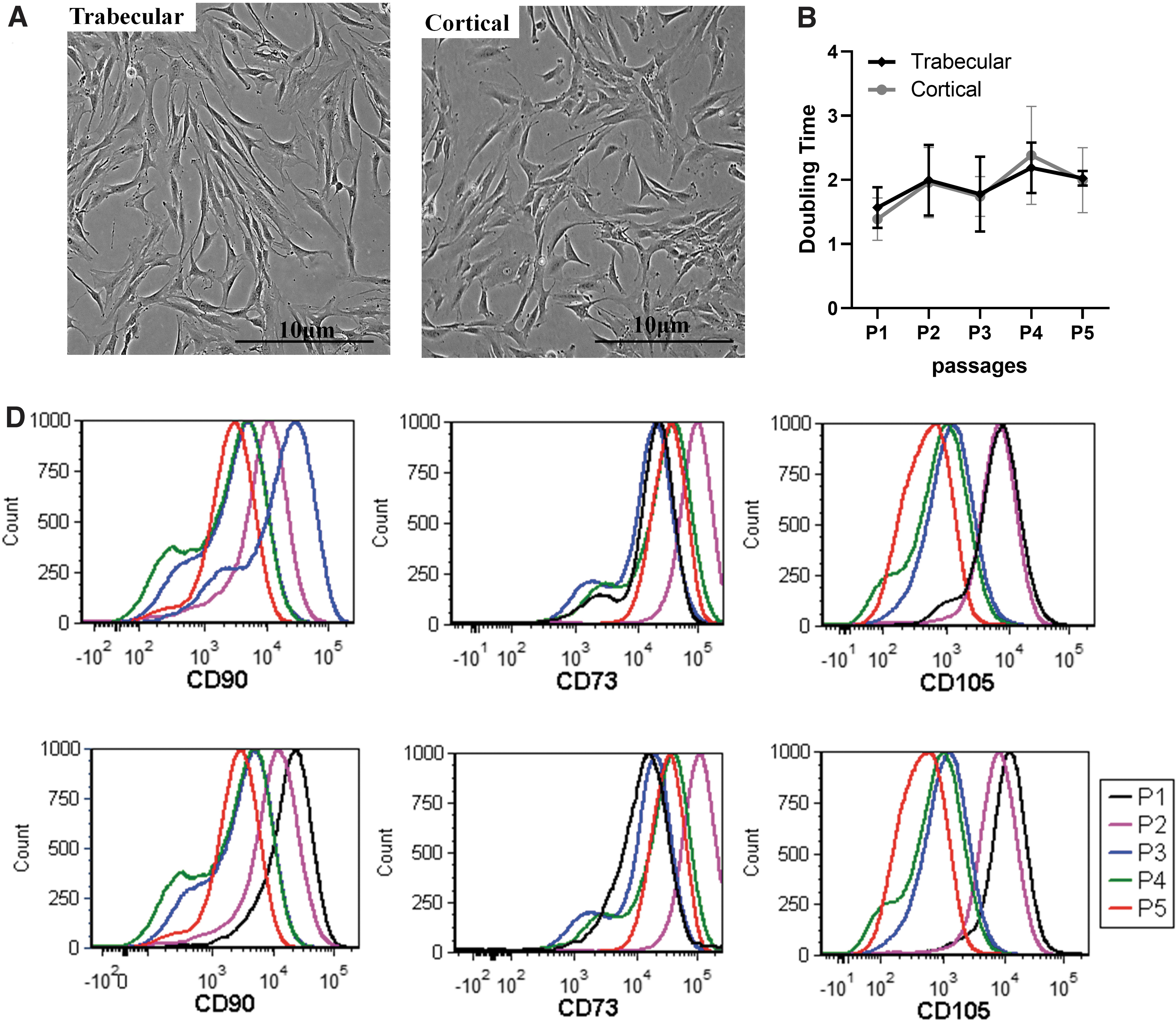
hPLS Cancellous Trabecular and Cortical MSCs isolation and stem cell characterization.
Summary Data of Patient Information
CKD, chronic kidney disease; HTN, hypertension; HLD, hypersensitivity lung disease; GERD, gastroesophageal reflux disease; HCC, hierarchical condition categories.
The immune phenotype of MSCs was characterized by evaluating the expression of the stem cell markers CD90, CD73, and CD105 and the absence of hematopoietic stem cell markers CD34, CD45, CD11b, CD19, and HLA-DR. We observed consistent and similar antigen expression profiles between the cortical and trabecular MSCs analyzed by flow cytometry (Fig. 1C, D) over the first 5 in vitro passages. An increased of antigen expression profiles was observed over time with higher percentages of triple positive cells at passage 4/5 in the trabecular versus cortical (Table 2) and no expression of HSC negative markers (Fig. 1C, D).
Percentages of Human Posterior Lumbar Spine Trabecular and Cortical Mesenchymal Stromal Cells
Percentages of negative anti-hematopoietic lineage cocktail (CD45, CD34, CD14, CD19, and HLA-DR) and triple positive markers (CD90, CD105, and CD73) of hPLS MSCs trabecular versus cortical at different passages (from P1 to P5) by Flow cytometry analysis.
In particular, we observed an average of 62% ± 8% versus 65% ± 5% triple positive cells in trabecular samples versus cortical ones respectively, reaching the highest % at passage 4 for trabecular MSCs while for cortical samples the percentage of triple positive cells was higher at earlier passages and decreased over time (70% ± % to 64% ± 4%). Similar phenotype and no significant changes in morphology was observed in different passages (P1–P5) as shown in the representative scatter plot (Supplementary Fig. S1).
Multilineage differentiation and molecular characterization
The multi-differentiation potential of hPLS cortical and trabecular samples was assessed. Osteogenic, chondrogenic, and adipogenic phenotypes were induced for 21 days. As shown by van Koss, Alizarin Red, Alcian Blue, and Oil red staining in Fig. 2A, in comparison to untreated cells (negative control), both cortical and trabecular MSCs demonstrated the ability to differentiate in osteoblasts, chondrocytes, and adipocytes similar to MSCs isolated from bone marrow (hBM-MSCs, positive control). However, these results were not completely supported by the mRNA analyses. To evaluate differentiation, associated genes expression levels RUNX2, ALP, SPARC, BGLAP, and SPP1 for osteogenesis; SOX9, ACAN, COL1A1, and COL2A1 for chondrogenesis; PPAR gamma, ADIPOQ, LEP, and LPL for adipogenesis were determined after 21 days.
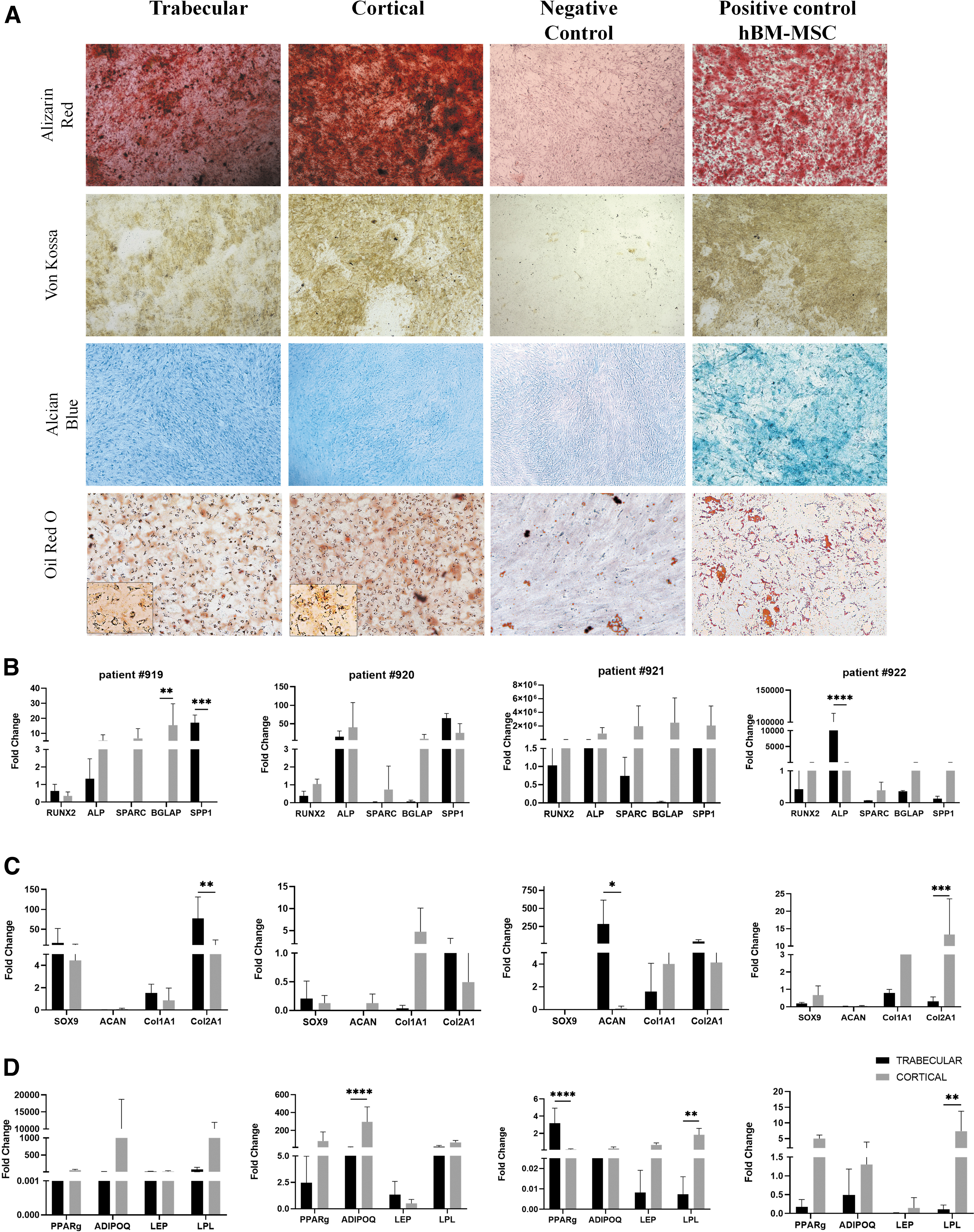
hPLS Samples Cancellous Trabecular and Cortical and their potential differentiation.
Because MSCs typically take 21 days to differentiate, we expected to observe an upregulation of genes expressed at the latest stage of each differentiation: SPP1 for osteogenesis; COL1A1 and COL2A1 for chondrogenesis; and LPL, LEP, and ADIPOQ for adipogenesis (Fig. 2B–D).
Overall, cortical MSCs showed higher expression of osteogenic gene markers. However, in sample #919, SPP1 was increased 146-fold (P < 0.0001) in cortical MSCs versus trabecular while in sample #922, ALP was higher (P < 0.0001) in trabecular MSCs versus cortical MSCs. This suggests that the two MSCs lineage might be in different stages of the differentiation process (Fig. 2B). Conversely, we did not observe a specific pattern in chondrogenesis gene expression between samples from the same group or a consistent difference between the trabecular and cortical specimens (Fig. 2C). Adipogenesis genes were most consistently upregulated in all four samples, in particular cortical MSCs differentiated better than adipocytes compared to trabecular MSCs (sample #920: ADIPOQ was increased 31.64-fold (P < 0.0001); sample #921: LPL was increased 245-fold (P < 0.001) and sample #922: LPL was increased 58.38-fold (P < 0.005) (Fig. 2D).
Immunosuppressive potential
To assess the immune tuning capability of MSCs, mRNA expression levels of IDO1, PTGES, and PTGS2 was evaluated in the two cell subtypes after 6-12-24-48-72 h of stimulation with TNFα and INFγ in vitro.
We first determined the optimal treatment dose for TNFα and INFγ. A dose titration experiment was performed in hBM-MSCs at 72 h and combination treatment of 100 ng/mL TNFα plus INFγ was identified as the optimal condition (Supplementary Fig. S2A), because the doses were not associated with any significant cytotoxicity or changes in MSCs marker expression at 24, 48, and 72 h (Supplementary Fig. S2B, C).
When trabecular and cortical MSCs were analyzed, we observed different gene expression patterns for IDO1, PTGES, and PTGS2 expression, which are key players in the suppression of chronic inflammation. After 12 h of treatment, IDO1 expression was significantly upregulated, and continued to increase until 48 h (Fig. 3A).
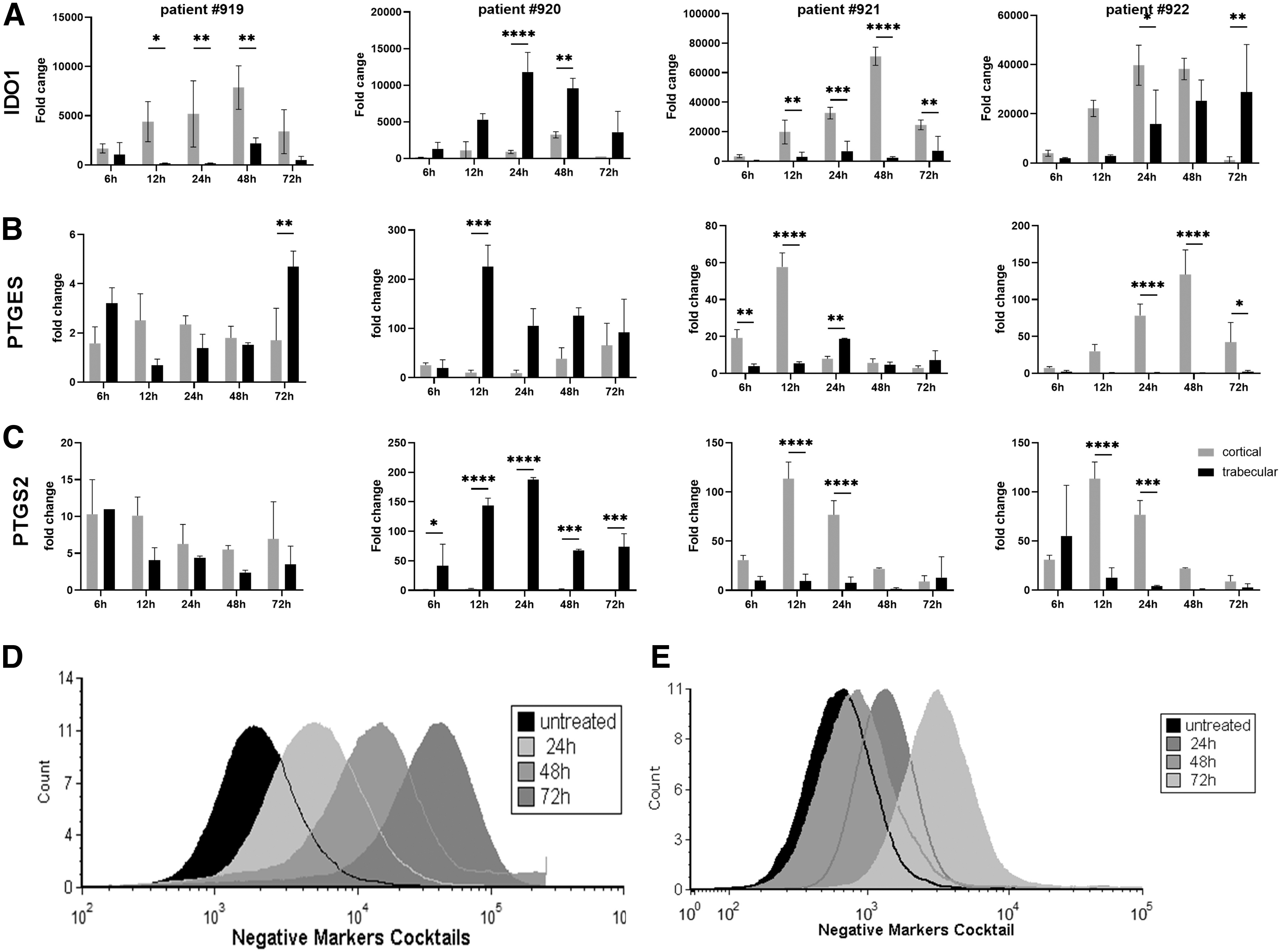
Immunomodulatory analyses of hPLS-MSCs.
In particular, in samples #919, #921, and #922, cortical MSCs showed significantly increased expression over time compared to trabecular samples, with a peak of expression from 24 and 48 h [sample #919: cortical MSCs vs. trabecular were increased 31.6-fold (P < 0.05), 37.28-fold (P < 0.001), and 3.6-fold (P < 0.005) at 12, 24, and 48 h respectively; sample #920: cortical MSCs vs. trabecular were increased by 6.8-fold (P < 0.05), 4.8-fold (P < 0.0001), 32-fold (P < 0.0001), and 3.4-fold (P < 0.05) at 12, 24, 48, and 72 h respectively; sample #922: cortical MSCs vs. trabecular were increased by 7.5-fold, 2.5-fold (P < 0.05), and 1.5-fold at 12, 24, and 48 respectively].
A completely different pattern was observed in sample #920; there was a higher level of IDO1 gene expression in trabecular MSCs compared to cortical MSCs, which was increased after 12 h of treatment, reaching its peak at 24 h [increased by five-fold (P < 0.005), eight-fold (P < 0.0001), and four-fold (P < 0.005) at 12, 24, and 48 h respectively].
For PTGES and PTGS2 (Fig. 3B, C), in sample #919, we did not observe consistent differences in gene upregulation between trabecular and cortical MSCs. Conversely, sample #920 showed higher gene expression in trabecular MSCs compared to cortical MSCs, as previously observed in IDO1. In particular PTGES showed a peak of upregulation at 12 h and PTGS2 at 24 h that were maintained until 72 h [PTGES increased by 22-fold (P < 0.0001), 11.2-fold (P < 0.05), 3.2-fold (P < 0.05), and 3.4-fold (P < 0.05) at 12, 24, 48, and 72 h, instead PTGS2 increased by 41.6 (P < 0.05), 57-fold (P < 0.0001), 215-fold (P < 0.0001), 55-fold (P < 0.0005), and 160-fold (P < 0.0005) at 6, 12, 24, 48, and 72 h respectively] (Fig. 3B, C). In sample #921 and #922, we observed similar pattern with the cortical MSCs showing higher gene expressions compared to trabecular MSCs.
For patient #921, PTGES upregulation started after only 6 h of treatment reaching a peak at 12 h, while the peak of PTGS2 was at 12 h and was maintained until 24 h [increased by 4.9-fold (P < 0.001), 10.7-fold (P < 0.0001), and 0.4-fold (P < 0.05) at 6, 12 and 24 h, and increased by 11.8-fold (P < 0.001) and by 9.8-fold (P < 0.0001) 12 and 24 h respectively for PTGES and PTGS2]. Patient #922 instead, showed upregulation of PTGES at 12 h maintained till 72 h, reaching the peak at 48 h [increased by 60-fold, 137-fold (P < 0.0001), 236-fold (P < 0.0001) and 22.7 (P < 0.05) at 12, 24, 48, and 72 h]. For PTGS2 gene, we observed similar upregulation pattern than patient #921, with a peak at 12 h sustained for 24 h [increased by 8.85-fold (P < 0.0001), 18.6-fold (P < 0.005) and 21.76-fold at 12, 24, and 48 h (Fig. 3B, C)].
We also evaluated the stem cell surface markers expression after treating with TNFα and INFγ. Flow cytometry analysis revealed that treatment altered the morphology and the expression of MSC surface cell markers in both cell subsets in a time-dependent manner (Fig. 3D, E and Supplementary Fig. S3). In particular, the percentage of trabecular MSCs negative for human leukocyte antigen–DR isotype HLA-DR, CD45, CD11b, CD19, and CD34 decreased dramatically after 24 h of IFNγ/TNFα treatment (Fig. 3D and Supplementary Fig. S3A), in cortical MSCs we observed this decrease only after 72 h of treatment (Fig. 3E and Supplementary Fig. S3B).
Discussion
The treatment of bone fractures and trauma-related or overuse joint injuries are a large focus within orthopedic surgery. Due to adequate evidence-based research, the successful treatment of these pathologies has greatly advanced over the past several decades. Moreover, improvements in technology have facilitated the decrease in treatment-related complications. However, certain complications such as non-union and delayed union continue to be common in the field [50], despite advances in treatment.
Spine fusion is a common procedure within orthopedics, but the success of the surgery relies on the ability of the patient to have adequate bone formation and fusion. The procedure is performed when patients present with traumatic, degenerative, or oncological spine disease. This includes disorders such as spondylolisthesis, deformity, trauma, neoplasm, and infection [51].
Over the past several decades various surgical techniques and approaches have been developed and implemented to increase fusion rates and decrease surgery related complications. For example, autologous bone grafts are considered the gold standard for spinal fusion procedures due to their osteogenic, osteoinductive, and osteoconductive ability [52]. However, there are a multitude of complications associated with obtaining iliac crest autografts, and obtaining an autograft can cause donor site pain, increased operative time, nonunion post-lumbar fusion, and often times adequate graft sites are limited [1,53 –55].
Due to these disadvantages, allograft bone is commonly used instead of autologous bone grafts to avoid donor site morbidity. However, allografts are associated with increased risks of infection and rejection and also have been demonstrated to have poorer osteoinductive properties [8].
As a result of the limitations in current treatment, research has focused on identifying various bone substitutes [9,10], along with novel and effective methods for treatment. The use of MSCs has drawn a significant amount of interest [3,4] because of their ability to differentiate to many different types of tissues, including muscles, bones, fat, and cartilage. In addition, MSCs are able to be harvested from many sources (ie, bone, tendon, skin, adipose tissue, umbilical cord, blood, and amnion) [56 –61]. Due to these characteristics, MSCs provide a promising area of research within the medical field [62].
Historically, many sources of stem cells have been studied for orthopedic procedures. Though adipo-derived MSCs (AD-MSCs) and BM-MSCs have been most preferred sources of MSCs, there is still controversy about the most effective source of MSCs for orthopedic procedures [63].
Fernandez-Moure et al. [64] compared human cortical bone fraction (CBF-MSCs), BM-MSCs, and AD-MSCs, and demonstrated that CBF-MSCs had superior osteogenic potential compared to BM-MSCs and AD-MSCs grown in both normoxia and hypoxia conditions. Similarly, Veriter et al. [65] used AD-MSCs in patients who had bone nonunion and nonhealing chronic wounds and was able to achieve successful outcomes in both clinical scenarios with no serious adverse or side effects.
Moreover, preclinical and clinical results have also had positive results in the application of MSCs. Fernandez-Bances et al. demonstrated that when MSCs are directly applied to sites where bone loss occurred, MSCs are able to stimulate bone regeneration. [66] In the study, seven patients with pseudoarthrosis of a long bone were treated with BM-MSCs isolated from the iliac crest, and positive results were achieved in all cases [67].
However, many studies have demonstrated the beneficial effects of MSCs in the preclinical and clinical setting, extraction methods for MSCs have yet to be optimized before utilizing them in orthopedic procedures such as spinal fusion. VB-BM is a well-known source for MSCs transplantation [68,69], but the use of VB-MSCs in spinal fusion is not yet a routine, despite the observed benefit of combining MSCs with scaffolds that enhance bone formation [70,71].
There are limited results and studies examining the use of MSCs in spinal fusion. Studies evaluating the effect of a combination of IC-BM aspirates and/or local bone autografts were able to demonstrate that MSCs from IC-BM aspirates helped improve outcomes in preclinical spinal fusion models, and spinal fusion in the clinic [72 –74]. However, the outcomes of the clinical trials were widely dependent on the quality of the BM [75,76]. In a donor-matched study, Fragkakis et al. analyzed the effect of BM processing methods on the quantity of VB-BM and IC-BM MSCs. They were able to conclude that red blood cell (RBC) lysis was a more effective method to process VB-BM compared to gradient centrifugation. Moreover, RBC lysis had minimal MSC loss compared to other methods [77].
There is controversy about the best site and method to isolate MSCs used in spinal procedures and the efficacy of MSCs obtained from vertebral spinal processes is still being delineated. Some studies have demonstrated that vertebral MSCs can grow and differentiate with greater efficiency compared to MSCs derived from iliac crest in the in vitro setting [16]. Moreover, others have demonstrated that MSCs derived from the cancellous bone in vertebral spinal processes are superior compared to MSCs isolated from the vertebral lamina or iliac crest [78]. However, even if it has been shown that MSCs from vertebral bodies have higher efficacy in inducing osteogenesis, our data suggest that there is a large heterogeneity in the MSC cell population derived from both the cortical and trabecular bone in vertebral spinous processes.
We demonstrated that MSCs from trabecular or cortical spine had MSC morphology, fibroblastic-like shape, expression of stem cell markers (CD90, CD73, and CD105), and the typical multilineage differentiation ability of stem cells, but we were unable to determine a consistent pattern in the gene expression of the specific markers in the adipocyte, chondrocyte, or osteoblast lineage. While we observed no differences between the trabecular and cortical specimens for chondrogenesis gene expression and mostly higher adipogenesis gene expression in cortical samples, the differences observed in osteogenic gene markers expression suggested that the two cortical and trabecular MSCs lineage might be in different stages of the differentiation. Overall, our data suggest that MSCs from vertebral bodies may not be an adequate and reliable source for MSC-based therapeutic strategies for regeneration and repair of spine fusion due to variable efficacy.
Our results were not confirmed by prior studies where instead they demonstrated the osteogenic potential of MSCs from cortical bone. [64] showed the trilineage differentiation capability of MSCs derived from cortical fraction of the bone was superior in comparison to MSCs harvested from bone marrow or adipose. This effect was even more pronounced in hypoxic conditions. This study suggested that MSCs derived from cortical bone could potentially be suitable candidates for orthopedic tissue engineering and regenerative medicine applications. The discordance in prior studies and our study could be due to multiple factors: differences in harvesting and processing techniques, inability and difficulty in completely separating cortical and trabecular bone (causing an impure cortical or trabecular MSC line), and limited sample number.
The limited sample number was apparent due to the variability in the relative gene expression of cortical versus trabecular was widely different between patient samples. Though there were mixed results in gene expression that would support the terminal differentiation of MSCs, there were positive results in the ability of MSCs from either site to be immunosuppressive. However, there was not a consistent or substantial difference between MSCs obtained from cortical or cancellous bone; MSCs from both sites demonstrated an immune suppression marker profile. This suggests that MSCs obtained from hPLS may have more success in applications related to immunomodulatory treatment rather than as a tool to support and drive osteogenesis to prevent nonunion in spine fusion procedures.
Conclusion
In conclusion, our study was unable to demonstrate a consistent difference between MSCs obtained from cortical and trabecular bone in hPLS. Though we were able to isolate MSCs from all samples, results were inconsistent between and among samples. Due to the large variability in the properties of MSCs obtained from the site, additional research is needed to further determine the potential use as a source for MSCs and their clinical application in spinal fusion procedures. Though it would be beneficial and more simplistic to be able to harvest MSCs from the same surgical incision the spinal fusion was performed, to decrease operation times and patient morbidity, it may not be an efficacious alternative to obtain MSCs from more traditional sites such as adipose tissue or iliac crest BM.
Footnotes
Author Disclosure Statement
The authors declare that this research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
Research reported in this publication was supported by internal seeding grant from the Orthopedic and Sports Medicine Department (OPPI) at Houston Methodist Hospital.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.