
Abstract
Adipose-derived stromal cells (ADSCs) have well-established regenerative and immunomodulatory properties. For such reasons, ADSCs are currently under investigation for their use in the setting of both regenerative medicine and autoimmune diseases. As per dermatological disorders, mesenchymal stromal cell (MSC)-based strategies represent potential therapeutic tools not only for chronic ulcers and wound healing, but also for immune-mediated dermatoses. However, a growing body of research has been focusing on the role of MSCs in human cancers, due to the potential oncological risk of using MSC-based strategies linked to their antiapoptotic, proangiogenic, and immunosuppressive properties. In the dermatological setting, ADSCs have shown not only to promote melanoma growth and invasiveness, but also to induce drug resistance. In contrast, genetically modified ADSCs have been demonstrated to efficiently target therapies at tumor sites, due to their migratory properties and their peculiar tropism for cancer microenvironment. The present review briefly summarizes the findings published so far on the use of ADSCs in the dermato-oncological setting, with the majority of data being available for melanoma.
Adipose-derived stromal cells (ADSCs) are a peculiar subtype of mesenchymal stromal cells (MSCs) located in fat tissue [1]. MSCs were first identified in the bone marrow as multipotent spindle-shaped cells [bone marrow stromal stem cells (BMSCs)] [2 –4]. Since their first characterization in the bone marrow, many others tissues and organs have been demonstrated to host a MSC reservoir, including dental tissue, skeletal muscle, tendons, synovium, dermis, subcutaneous fat, liver, lungs, placenta, umbilical cord, amniotic fluid, and breast milk [5 –11].
In 2006, three minimal criteria for defining MSCs were identified by the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy: adherence to plastic under standard culture conditions, multipotency (ability to differentiate into chondroblasts, osteoblasts, and adipocytes in vitro), and specific surface markers, such as CD105, CD73, and CD90 [12]. By definition, MSCs lack the expression of hematopoietic markers, such as CD34, CD45, CD14, or CD19 [12]. More recent revisions of MSC nomenclature and classification also included STRO-1, CD106, and CD146 as MSC-defining markers [13,14] (see Fig. 1).
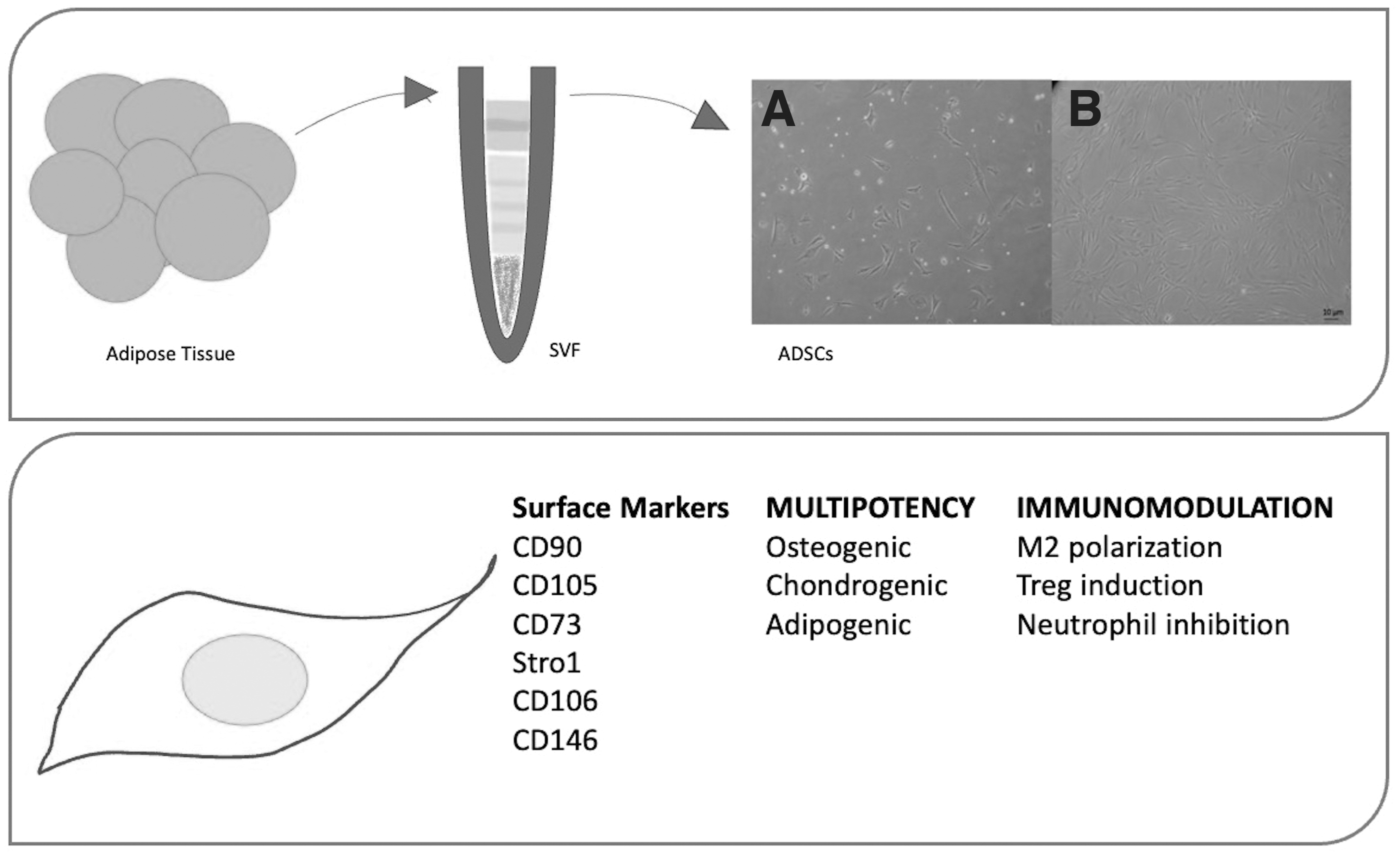
ADSC extraction and characterization. Upper panel: Fat tissue (obtained either through liposuction and surgical excision) undergoes mechanical and/or enzymatic digestion to obtain the SVF. Such cell population, naturally enriched in ADSCs, is then plated in plastic flasks/dishes. Cellular morphology is shown for SVF
MSCs display a large variety of potential clinical applications, due both to their multipotency and their peculiar immunological features. In the clinical setting, ADSCs represent a valid alternative to BMSCs due to their relative abundancy and availability in the human body [15]. First, subcutaneous fat is easily accessible and, therefore, isolation procedure is particularly safe for patients [16,17]. Second, ADSCs represent ∼2% of the viable cells in lipoaspirates, whereas only ∼0.02% of the nucleated cells in the bone marrow are BMSCs [18,19]. However, the use of lipoaspirates as effective MSC sources requires further validation in the clinical setting, due to the pro-inflammatory milieu represented by fat tissue from obese donors potentially having an impact on MSC function.
Current literature suggests that the secretion of a large variety of growth factors, cytokines, and chemokines gives reason of most of their regenerative and immunomodulating properties [5,20,21]. Several authors also hypothesized a role for MSCs in cancer genesis and dissemination [22,23]. Recently, a growing body of research has been focusing on the role of MSCs in human cancers, due to the potential risk of using MSC-based strategies linked to their antiapoptotic, proangiogenic, and immunosuppressive properties. Owing to their peculiar location and migratory capabilities, ADSCs can be considered part of tumor microenvironment in most skin and soft-tissue neoplasms.
With regard to cutaneous oncology, the role of MSCs on melanoma has widely been studied in the past few years, but current knowledge does not enable to draw similar conclusions on a possible pathogenic role in vivo [24]. Moreover, controversial data have recently been published on the effects of MSC secretome on melanoma growth, therefore leaving an open question on the topic [25 –27]. If, on the one hand, MSCs carry the risk of having a promoting role on cutaneous neoplasms, on the other hand, migratory properties at tumoral and inflammatory loci could also make them ideal vehicles for carrying therapeutic genes and/or proteins at tumor sites.
The aim of this study is to give a comprehensive overview specifically on ADSCS and skin cancers (see Fig. 2), including both melanoma and nonmelanoma skin cancers (NMSCs). Table 1 briefly summarizes the major publications in this setting.
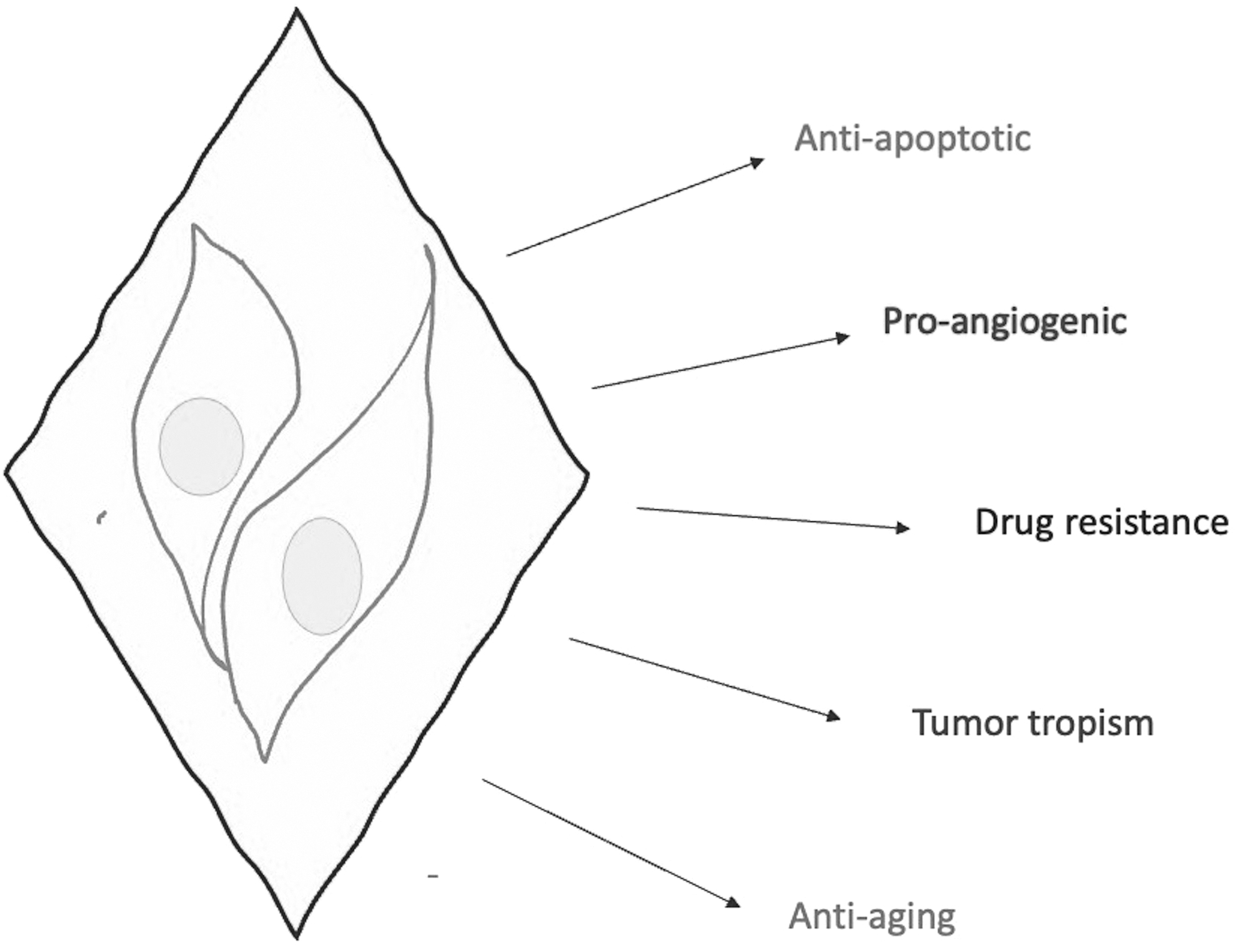
Multifaceted role of ADSCs in cutaneous neoplasms. ADSCs seem to have a tumor promoting role due to their antiapoptotic and proangiogenic properties. Moreover, drug resistance has been demonstrated to be possibly linked to interactions between ADSCs and cancer cells. However, ADSCs could also be efficiently exploited for vehiculation of anticancerous drugs at tumor sites due to their peculiar tropism for tumor microenvironment. Finally, ADSCs possess antioxidant and antiaging properties.
Main Publications Specifically Focusing on Adipose-Derived Mesenchymal Stromal Cells and Skin
ADSC, adipose-derived mesenchymal stromal cell; CM, conditioned medium; COX, cyclo-oxygenase; eNOS, endothelial nitric oxide synthetase; HMB, high mobility group; IL-2, interleukin 2; MAPK, mitogen-activated protein kinases; MDR, multidrug resistance; NF, nuclear factor; SCC, squamous cell carcinoma; TGF, transforming growth factor; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; TRP1, tyrosinase-related protein 1; TSG-6, tumor necrosis factor-stimulated gene-6; UV, ultraviolet; vWF, von Willebrand factor.
Tumor-Promoting ADSC
ADSCs in tumor microenvironment have recently been demonstrated to promote tumor growth and invasiveness [28 –30].
Preisner et al. evaluated cell-growth kinetics in different types of melanoma cell lines (G-361-MMCs, A2058-MMCs, SK-Mel-5-MMCs, and MeWo-MMCs) and demonstrated melanoma-cell proliferative and migratory activity to be significantly increased when cocultured with ADSCs [24]. The authors also identified a subset of overexpressed genes potentially responsible for such phenomenon, including CXCL12, PTGS2, IL-6, IL-8, MMP-2, and HGF.
ADSCs also seem to directly promote cancer cell growth and migration. In fact, both ADSCs and BMSCs have been described to stimulate migration of melanoma B16 cells [24,31]. Compared with BMSCs, ADSCs also promoted tumor growth, possibly by enhanced secretions of specific cytokines and growth factors, with a central role for CCL2 [32].
Not surprisingly, similar results were found in the setting of squamous cell carcinoma (SCC). In fact, migratory and invasive properties of SCC were demonstrated to significantly increase in cancer cells and ADSCs cocultures [23]. The same study also demonstrated coculture conditioned media to promote angiogenesis in vitro. Such changes have been shown to be associated with overexpression of specific cytokines and chemokines (CCL2, CCL4, IL6, and IL1beta) and, even more importantly, of several metalloproteases (MMPs) and growth factors (MMP3, MMP10, and VEGF) [23]. Notably, interleukin (IL)-1 promotes the expression of several genes, included those coding for MMPs, VEGF, IL-6, and CCl2 [33]; such molecules are mainly responsible for tumor growth, invasion and metastasization. Nevertheless, IL1-beta also induces cyclooxygenase-2 (COX-2), possibly representing the joining link between inflammation and drug resistance [34].
ADSC and Drug Resistance
A major obstacle in cancer treatment is represented by tumor cell resistance to chemotherapies. Endogenous MSCs in tumor microenvironment have been demonstrated to affect drug resistance [35]. Through a metabolomic approach, Roodhart and collaborators identified platinum-induced polyunsaturated fatty acids (PIFAs) to be associated with resistance to platinum analogs [36]. Such mechanism of resistance was shown to rely upon specific signaling between MSCs and cancer cells. Moreover, the inhibition of COX-1 and thromboxane synthase was described to revert MSC-induced resistance.
COX-2 has already been proven to promote tumor growth, angiogenesis, and metastatization [37 –39]. Several studies also demonstrated that COX-2 can mediate drug resistance and that such mechanism has a big impact on oncological patient survival [38,40,41]. Previous studies demonstrated that tumor necrosis factor alpha-stimulated gene/protein 6 (TSG-6) upregulates COX-2 expression in inflammatory conditions [42,43]. In in vitro animal models, the addition of ADSCs to melanoma cells (LMeC canine cell line) determines an increase in COX-2 expression [44]. Such significant variations are absent when siTSG-6-transfected ADSCs are cocultured with cancer cell, therefore demonstrating TSG-6-mediated COX-2 upregulation to be a possible mechanism of chemoresistance development induced by ADSCs.
ADSC-Based Therapeutic Strategies for Cutaneous Neoplasms
Owing to their selective migratory properties, ADSCs represent a powerful tool for targeting gene therapies at tumor site [45,46]. Advanced-stage melanoma poses a great therapeutic challenge for oncologists due to its quick dissemination and poor prognosis [47 –49]. The lack of tumor specificity gives reason of the frequent side effects of most of the therapies currently used in the clinical practice. Therefore, more specific therapeutic strategies exclusively targeting cancerous tissues are urgently needed. Since tumor-directed migration has already been described in several publications for MSCs, more recent studies are focusing on their potential role in vehiculating anticancerous agents [46,50].
Jing et al. in 2016 demonstrated that human ADSC cell lines transfected with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-containing plasmids cud efficiently induce apoptosis in A375 melanoma cells, using a Transwell coculture system [51]. Melanoma-cell apoptosis seemed to be associated with caspase 4 and 8 activation. Recombinant TRAIL (rTRAIL) has already been demonstrated to harbor antitumor activity, but its short half-life limits rTRAIL use in vivo [52]. On the contrary, TRAIL-ADSC mediated apoptosis was observed to be dose dependent and even more prominent after 48 h of coculture. Such data confirmed the hypothesis that ADSC could be used as a TRAIL-delivering source for melanoma [53].
An interesting study on MSC-based therapeutic strategies for melanoma focused on anticancer vaccination approach using immunoproteasome-expressing MSCs, but only BMSCs were used as MSC source [54].
ADSCs as a Tool for Understanding IL-2 Chemoresistance
IL-2 is involved in T cell survival and proliferation and is crucial for T effector cell differentiation and function [55,56]. In the past decades IL-2 has been widely used a therapy for patients with advanced melanoma since it was the only available treatment for stages II and III before the advent of BRAF/MEK inhibitors and immunotherapies [57 –59].
However, benefits from high-dose IL-2 therapy are limited to a small fraction of treated cases and, therefore, IL-2 is currently being replaced by checkpoint inhibitors and targeted therapies [60 –62].
Notably, no endogenous expression of IL-2 is generally present in MSCs, but efficient transfection can be achieved in such cells with subsequent implications for therapeutic ex vivo expansion of immune cells [63].
With regard to cutaneous oncology, an Iranian research group demonstrated that genetically modified ADSCs expressing IL2 increased viability and reduced apoptosis of B16F10 melanoma cells [64]. Such findings were also confirmed in vivo in a murine model of metastatic melanoma, where co-injection of IL2-expressing ADSCs with melanoma cells significantly enhanced melanoma growth.
Taken together, these data give new insights on the biological bases of IL-2 therapy side effects and on open a new scenario on ADSCs as a powerful tool for assessing drug safety.
ADSC Differentiation Potential
Owing to their multipotency, ADSCs can differentiate into a large variety of cellular types of mesodermal origin [12,65]. However, recent reports also describe ADSC capability to differentiate into other cellular types deriving from other embryonic epidermal sheets.
Marino and coauthors, for example, demonstrated ADSCs differentiation into endothelial cells when grown on polycaprolactone [66]. Other groups achieved similar results with MSCs and single-walled carbon nanotubes [67].
A Brazilian study demonstrated umbilical-cord derived MSCs to differentiate into keratinocytes in the presence of a defined keratinocyte serum-free medium [68]. The researchers postulated such mechanism to occur in vivo in the human epidermis, but no studies are currently available for ADSCs on this topic.
Under specific differentiation conditions, ADSCs were also described to develop melanosome, express melanocyte-specific proteins, and display human melanocyte-like properties in vivo when injected into the dorsal skin of immunocompromised mice [69].
Such findings pave the way to ADSC use both in the setting of wound healing and vitiligo treatment. However, whether such proliferative and differentiative potential could be responsible for some cases of melanoma, NMSC, or vascular neoplasms remains unknown. Therefore, future studies aimed at assessing a possible direct pathogenic role for ADSCs (and MSCs in general) in cutaneous carcinogenesis are urgently needed.
A Protective Role Against Photodamage
Notably, chronic ultraviolet (UV) exposure determines an impairment in collagen deposition and distribution [70,71]. UV-A radiation seems to be mostly responsible for inducing wrinkling pigmentary changes and loss of elasticity, due to its penetration deep in the dermis [72,73]. Oxidative stress plays a pivotal role in driving UV-induced cutaneous changes. Several studies have been published so far on the efficacy of MSC-based strategies in the setting of photoaged skin [73,74].
Antioxidant effects of ADSC secretome are described to occur through the production of enzyme scavenging reactive oxygen species, such as catalase, glutathione peroxidase, and/or superoxide dismutase [73,75].
ADSCs and ADSC-conditioned medium (ADSC-CM) injection has also been demonstrated to increase dermal thickness and collagen content in animal models, therefore having a potential rejuvenating and antiwrinkle effect [76]. Other groups demonstrated an improvement in skin wrinkling with ADSCs in murine models of photoaging [77]. Not only ADSCs possess well-established collagen secreting properties that could partially explain the aforementioned results [65], but also Wnt/β-catenin pathway seems to contribute to the ADSC-mediated activation of dermal fibroblasts [78].
Pigmentary changes typically found in sun-damaged skin [79 –81] are commonly treated with antioxidant and depigmenting agents that are able to revert the clinical signs of chronic photoaging and to restore normal cutaneous pigmentation [82,83]. As other depigmenting agents, ADSCs also seem to have a whitening effect. Reduced melanogenesis has been demonstrated in melanoma B16 cells after treatment with ADSC-CM [84]. Tyrosinase and TRP1 downregulation was mediated by transforming growth factor beta 1 (TGF-β1) in such in vitro model. Inhibition of melanogenesis was also confirmed by Fontana–Masson and HMB-45 staining on pathological samples from in vivo studies on C57BL/6 mice injected intradermally with ADSCs and subsequently irradiated with UVB (150 mJ/cm2) [85].
Discussion
ADSCs and other types of MSCs are certainly of interest because of their regenerative properties and, therefore, are currently considered one of the most promising tools for a wide variety of medical settings (e.g., cardiovascular disorders and neurodegenerative diseases) [86 –89].
In particular, the dermatological setting represents a perfect stage for MSCs due to their collagen-secreting and pro-epithelizing role in the wound healing process [65,90 –92].
Anti-inflammatory and/or immunoregulatory properties also represent fundamental attributes for ADSCs next to their regenerative potential [93,94]. An increasing number of clinical trials are currently using MSCs for the treatment of severe degenerative and immune-mediated diseases with the aim of avoiding side effects of conventional immunosuppressant therapies [95 –98]. In recent years, MSCs have also demonstrated to be considered as a cell-based therapy for the treatment of COVID-19-related clinical manifestations [99,100].
Immunoregulatory properties of MSCs have been described to depend on both direct cell-to-cell interaction and the secretion of soluble mediators [101].
In the context of wound healing, ADSCs were recently demonstrated to interact with different cell types, including fibroblasts, macrophages, keratinocytes, melanocytes, and endothelial cells, mainly through the production of secretory molecules. The coordinated action of all these different cell types brings to the correct progression across the sequential phases of wound healing (inflammation, proliferation, and remodeling) and seems mainly to depend upon the regulated secretion of key growth factors and cytokines (e.g., TGF-β, GDF-11, SDF-1, and PDGF) by ADSCs [102].
Notably, MSC secretome contains different types of extracellular vesicles: exosomes, microvesicles, and apoptotic bodies [103]. Exosomes, characterized by the presence of specific surface markers (CD9, CD63, and CD81), originate from recycling of endosomes and are specifically considered to be crucial for most of MSC properties [104]. MSC secretome not only has been shown to account for most of the effects of MSCs in tissue regeneration but also has been demonstrated to be involved in MSC-mediated immunomodulation.
For this reason, the use of MSC secretome, mostly in the form of MSC-CM and MSC-derived exosomes, is currently under investigation for the treatment of immune-mediated diseases (such as lupus, dermatomyositis, inflammatory bowel diseases, multiple sclerosis, psoriasis, systemic sclerosis, and many others) to avoid the limitations associated with cell therapy [105 –107].
MSCs have been demonstrated to possess outstanding migratory properties and to be efficiently recruited at tumoral sites, therefore paving the way to MSC-mediated cancer therapy [108].
Several authors are currently studying new strategies for optimizing MSC chemotactic response to chemokines released from lesions, mainly using genetically modified MSCs overexpressing specific chemokine receptors [109].
As for anticancer MSC mediated therapy, TRAIL-engineered MSCs demonstrated promising results not only in melanoma treatment, as previously discussed, but also in hepatocellular carcinoma cells and both in vitro and in vivo models of brainstem glioma [45,50]. The use of engineered MSCs significantly lowers the risk of insertion mutagenesis in host DNA when compared with other types of genetically modified therapeutic vectors such as DNA-based plasmids [110,111].
The in vivo use of MSCs is currently limited by their potential tumor-promoting role (see Fig. 3). In fact, ADSCs and the other subtypes of MSCs exert an antiapoptotic, pro-proliferative, immunomodulating, and proangiogenic action. Such characteristics give reason of the potential role of ADSCs located adjacent to skin and/or soft-tissue neoplasms in promoting tumor growth and metastatization.

Main findings on ADSCs in the dermato-oncological setting. ADSCs are proven to differentiate into EC and melanocyte precursors (Pre-Mel) under specific differentiation conditions. ADSCs also contrast the effects of skin photoaging, which is often considered as a risk factor for skin cancer development. Not only ADSCs revert pigmentary changes through inhibition of Tyr and TRP-1, but also reduce sun-induced wrinkling by promoting fibroblast-mediated Col I secretion. However, ADSCs are demonstrated to promote tumor growth, invasion and distant spread both in melanoma and in squamous cell carcinoma; such tumor promoting-role is thought to occur through the production of several MMP, growth factors, and cytokines. Finally, genetically modified ADSCs have been used to explore possible mechanisms of MDR development and to assess their efficacy in vehiculating therapeutic targets at tumor sites. Col I, collagen I; COX, cyclo-oxygenase; EC, endothelial cells; MDR, multidrug resistance; MMP, metalloproteinases; TGF, transforming growth factor; Wnt: IL, interleukin; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; TRP-1, tyrosinase-related protein 1; TSG, tumor necrosis factor-stimulated gene; Tyr, tyrosinase.
Only few studies have deeply explored ADSCs interactions with immune cells, and little is known about the exact mechanisms underlying this peculiar interplay between MSCs and immune cells.
A major limit in better understanding of MSC immune interactions resides in the extreme heterogeneity of isolation techniques and sources of such cells [17]. For this reason, it is not always possible to draw conclusions generalizing findings validated for a specific MSC subtype to all other MSCs. Moreover, most of the available studies on MSC immunomodulatory functions are based on by in vitro tests of inhibition of T lymphocyte proliferation [112]. More specific functional assays focusing on the interplay between MSCs and immune cells are needed.
Also aging seems to have a deep impact on MSC function. A reduction in both MSC immunomodulatory and regenerative functions has recently been described to occur with aging [113,114]. Aged MSCs display reduced proliferative potential, and decreased secretome content, with cytokine and chemokine production being particularly affected [115]. Significant changes in cell expression profiles seem to bring to upregulation of HLA class II antigen expression in senescent MSCs, turning into a pro-inflammatory phenotype, with reduced immunosuppressive potential [116].
Production techniques, cell sources, and aging impact on cells as well as on secretome, giving reason of the wide range of MSC-derived acellular products described in literature, with variable characteristics in terms of anti-inflammatory action [117]. Moreover, cytokine content and immunoregulatory effects of secretory vesicles can also present significant variations depending upon the considered disease and the clinical or experimental setting.
Notably, tumor microenvironment and inflammation are ambivalently related to each other: if chronic inflammation is a well-established promoting factor in carcinogenesis, suppression of the local antitumoral inflammatory response remains one of the most explored mechanisms of cancer immune escape [118 –120]. Although pro-inflammatory environments seem to negatively impact on MSC regenerative function, inflammation seem to trigger MSC-mediated immunosuppression, and this could probably represent a cornerstone for better understanding the subtle relationship between MSCs and cancer.
MSCs reduce innate immune response, which is crucial for cancer surveillance [117]. As for the molecular and cellular mechanisms, MSCs have been described to inhibit neutrophil apoptosis in vitro, despite not having an impact on their phagocytic and chemotactic activity [121,122]. MSCs also induce macrophage M2 polarization, which eventually contributes to Treg induction [123,124]. Moreover, MSCs also directly interact with lymphocytes, dendritic cells, and natural killer cells, therefore giving reason of the complexity in delineating the exact MSC-mediated immune alterations in cancer.
When using ADSCs in vivo, there is always a risk of malignant or premalignant cells being located close to the transplanted ADSCs, and whether such risk could be lowered by the use of acellular ADSC-derived products, such as ADSC-CM as well as exosomes, still has to be clarified.
Very little is known on a potential promoting role for ADSCs in cutaneous carcinogenesis, with most of the available data being focused on melanoma (Fig. 3).
The exact mechanisms underlying ADSC-mediated immunosuppression remain unclear and the role of such immune interactions in skin cancer still has to be determined. In addition, it is not clear whether MSC functions could be altered under local and/or systemic inflammatory conditions, which often characterize advanced stages of neoplastic progression. This point is crucial to understanding whether MSC-based strategies could represent a double-edged sword in the setting of cutaneous oncology [125]. Despite the promising results achieved in ADSC-based therapies in the field of wound healing and for the treatment of immune-mediated dermatoses, safety concerns on the potential tumor-promoting action of these cells need to be methodically addressed before ADSCs could be routinely used in the clinical practice.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.