
Abstract
Cartilage is derived from the chondrogenic differentiation of stem cells, for which the regulatory mechanism has not been fully elucidated. N6-methyladenosine (m6A) messenger RNA (mRNA) methylation is the most common posttranscriptional modification in eukaryotic mRNAs and is mediated by m6A regulators. However, whether m6A regulators play roles in chondrogenic differentiation is unknown. Herein, we aim to determine the role of a main m6A reader protein, YTH N6-methyladenosine RNA binding protein 1 (YTHDF1), in chondrogenic differentiation regulation. Western blotting (WB) assays found that the expression of YTHDF1 increased during chondrogenic differentiation of human bone marrow mesenchymal stem cells (hBMSCs). The results of quantitative polymerase chain reaction, WB, immunohistochemistry, and Alcian blue staining revealed that overexpression of YTHDF1 increased cartilage matrix synthesis and the expression of chondrogenic markers when hBMSCs, ATDC5 cells, or C3H10T1/2 cells were induced to undergo chondrogenesis. Conversely, chondrogenesis was clearly inhibited when YTHDF1 was knocked down in hBMSCs, ATDC5 cells, or C3H10T1/2 cells. Further RNA sequencing and molecular biology experiments found that YTHDF1 activated the Wnt/β-catenin signaling pathway during chondrogenic differentiation. Finally, the effects of overexpression and knockdown of YTHDF1 on chondrogenic differentiation were reversed by inhibiting or activating β-catenin activity. Therefore, we demonstrated that YTDHF1 promoted chondrogenic differentiation through activation of the Wnt/β-catenin signaling pathway.
Introduction
Cartilage, which is rich in extracellular matrix, plays an important role in normal bone development and the maintenance of normal joint function [1,2]. The extracellular matrix of cartilage is mainly composed of collagen type II and aggrecan, both of which are synthesized by chondrocytes [3]. Chondrocytes are the only cell type present in cartilage tissue, and SRY-box transcription factor 9 (SOX9) is the most important transcription factor that regulates the synthesis of the cartilage matrix [4]. In mammals, cartilage tissue is mainly divided into the growth plate and articular cartilage [5]. Bones are mainly formed through endochondral bone formation, which relies on the coordinated development of the growth plate [6]. During endochondral bone formation, within embryonic limb buds, mesenchymal stem cells (MSCs) first undergo chondrogenic differentiation to form the growth plate, the calcified part of which is gradually replaced by bone tissue [6].
Previous studies have shown that abnormal growth plate formation not only impairs the length of bones but also affects bone density, ultimately leading to various bone deformities [3,7]. Located on the surface of the joint, articular cartilage has elasticity and toughness, and can distribute force, expand the load-bearing surface, and protect the joint from damage [8]. Cartilage defects are responsible for progressive joint damage in many acute and chronic osteoarthropathies, such as osteoarthrosis (OA) [9]. Methods of regenerating cartilage tissue have always been a difficult and hot topic in the field of OA treatment [10]. Due to the limited repairability of resident articular chondrocytes, stem cell implantation for chondrogenic differentiation is considered to be a more effective method for cartilage repair [10 –12]. Therefore, studying chondrogenic differentiation is not only of great significance for understanding endochondral bone formation but can also broaden the horizons for strategies of cartilage regeneration in OA disease.
N6-methyladenosine (m6A) messenger RNA (mRNA) methylation, the most prevalent posttranscriptional mRNA modification, has emerged as an important player in many biological processes [13 –15]. By affecting the splicing, stability, and translational efficiency of mRNA, it plays an important role in the posttranscriptional regulation of gene expression [15]. The modification process is dynamic and reversible, and is mediated by several m6A regulators that can be divided into writer, eraser, and reader proteins [16]. The writers include methyltransferase-like 3 (METTL3), Wilms tumor 1-associated protein (WTAP), and METTL14, which is also called the methyltransferase complex [17]. The methyltransferase complex catalyzes m6A methylation of mRNA [17]. Fat mass and obesity-associated protein (FTO) and alkB homolog H5 RNA demethylase (ALKBH5) were found to mediate the reversible removal of this methylation and are therefore referred to as eraser proteins [17].
In addition, the corresponding reader proteins include YTH N6-methyladenosine RNA binding proteins (YTHDF1, YTHDF2, and YTHDF3) and YTH domain-containing proteins (YTHDC1 and YTHDC2), which act as the main m6A binding proteins to regulate RNA metabolism, including mRNA splicing, degradation, and translation [18].
Recently, many studies have identified that m6A mRNA methylation is a crucial modulator of the regulation of stem cell pluripotency [19]. Some researchers have found that Pth1r mRNA methylation mediated by METTL3 promoted the osteogenic differentiation of MSCs [15]. Unexpectedly, ablation of METTL3 protein in mice promoted the development of osteoporosis through downregulation of RUNX2 protein expression [20]. In addition, FTO, an eraser protein, has been reported to act as a suppressor in the function of osteoblasts [21]. This evidence suggests the positive regulation of some m6A regulators in the osteogenic differentiation of stem cells. However, compared with osteogenic differentiation, the role of m6A regulators in chondrogenic differentiation has not been well-studied. Recently, two studies reported that YTHDF1, one of the reader proteins mediating the m6A methylation modification, is associated with apoptosis, autophagy, and hypertrophy of mature chondrocytes, suggesting that YTHDF1 may play an important role in chondrogenic differentiation [22,23].
In this study, we used three cell models, including human bone marrow mesenchymal stem cells (hBMSCs), ATDC5 cells, and C3H10T1/2 cells, to explore the role of YTHDF1 in chondrogenic differentiation. Our results found that YTHDF1 enhanced chondrogenic differentiation by activating the Wnt/β-catenin signaling pathway, and this is the first time that the role of m6A regulators in chondrogenic differentiation has been revealed. Our results indicated that YTHDF1 is potentially a good target for the treatment of chondrodysplasia and a strategic choice for cartilage regeneration in OA.
Materials and Methods
Cell culture and mouse breeding
The study was approved by the Ethics Committee of Sun Yat-sen Memorial Hospital of Sun Yat-sen University (approval No. SYSEC-KY-KS-2021-065, April 1, 2021), and written informed consent was obtained from all subjects included in the study. All the mice in this study were bred in the Laboratory Animal Center of Sun Yat-sen University. hBMSCs were isolated from bone marrow obtained from healthy volunteer donors as previously described [24]. Briefly, the bone marrow samples were diluted with phosphate-buffered saline (PBS). Cells were then fractionated using a Lymphoprep density gradient by centrifugation at 500g for 20 min. Interfacial mononuclear cells were collected, resuspended in low-glucose Dulbecco's modified Eagle's medium (DMEM; Gibco, Grand Island, NY), supplemented with 10% fetal bovine serum (FBS), seeded, and incubated at 37°C and 5% CO2.
After 48 h, nonadherent cells were removed by changing the medium. Thereafter, the medium was changed every 3 days. When the cells reached 80%–90% confluence, they were trypsinized, counted, and plated again. Cells from passages 3–6 were used for experiments.
The mouse chondrogenic cell line ATDC5 was purchased from the RIKEN Cell Bank (Tsukuba, Japan) and cultured in DMEM/F-12 (Gibco) supplemented with 10% FBS (HyClone). The murine MSC line C3H10T1/2 was purchased from ATCC and cultured in minimum essential medium α (αMEM; Gibco) containing 10% FBS and 100 U/mL of penicillin–streptomycin (HyClone).
Chondrogenic differentiation
As we have previously reported, the high-density micromass culture system was used for chondrogenic differentiation of hBMSCs and C3H10T1/2 cells [25]. Briefly, hBMSCs or C3H10T1/2 cells were trypsinized, washed with PBS, and then resuspended at 2 × 107 cells/mL in the OriCell™ Human MSC Chondrogenic Differentiation Medium (Cyagen Biosciences, Inc., Guangzhou, China), which consists of high-glucose DMEM supplemented with 10 ng/mL of recombinant human transforming growth factor-b3 (TGF-b3), 100 nM dexamethasone, 50 μg/mL of ascorbic acid 2-phosphate, 1 mM sodium pyruvate, 40 μg/mL of proline, and 1% ITS. Droplets (12.5 μL) were carefully placed in each interior well of a 24-well plate to form a sphere.
For chondrogenic differentiation of ATDC5 cells, cells were seeded in a six-well plate at a density of 60,000 cells per well, and the differentiation medium (DMEM/F-12 containing 5% FBS and 1% ITS) was replaced after 24 h. The medium was changed every 3 days, and cells were harvested on day 7.
Lentivirus and cell transduction
Full-length, wild-type human YTHDF1 or mouse Ythdf1 complementary DNA (cDNA) was cloned by polymerase chain reaction (PCR) into the GV358 vector plasmid (Genechem Co., Shanghai, China), and short hairpin RNA (shRNA) targeting YTHDF1 or Ythdf1 mRNA was cloned by PCR into the GV248 vector plasmid (Genechem Co.). Lentivirus packaging was performed by Genechem Co. For cell transduction, cells were seeded at a density of 106 cells per 100 mm dish in growth culture medium, and the next morning, they were incubated with viral supernatant and 5 μg/mL of polybrene (Sigma-Aldrich) for 16 h, and then, the normal culture medium was replaced. Seventy-two hours after infection, the cells were treated with 1 μg/mL puromycin (Sigma-Aldrich) to select stably transduced cells. The sequence information for shRNA cloning is provided in Supplementary Table S1.
Reverse transcription and real-time PCR analysis
Total RNA was extracted from micromasses or adherent cells using the TRIzol reagent (Sigma). Then, cDNA was synthesized by mixing 500 ng of total RNA and Prime-Script RT reagent kit with genomic DNA (gDNA) eraser (Takara) following the manufacturer's instructions. Quantitative reverse transcription PCR (qRT–PCR) was performed with a LightCycler 480 instrument (Roche) using SYBR Green I Master Mix (Roche) according to the manufacturer's instructions. The 2−ΔΔCt method was used to calculate relative expression levels. GAPDH or Gapdh served as the internal control for normalization. The specific primers used in this article are listed in Supplementary Table S2.
Measurement of mRNA half-life
After undergoing induction of chondrogenic differentiation for 7 days, the adherent ATDC5 cells (with or without YTHDF1 overexpression) were treated with 10 μg/mL of actinomycin D (MCE HY-17559) for 0, 0.5, 1, 2, 4, or 6 h. Then, the RNA was extracted from the samples and quantitative polymerase chain reaction (qPCR) was used to determine the levels of β-catenin mRNA. The half-life was calculated using the formula T1/2 = 0.3 t/log(D1/D2) [26].
Western blot analysis
The proteins of micromasses, adherent cells, or mouse embryonic limb buds were extracted with RIPA buffer (Beyotime, Shanghai, China) containing 1% phosphatase and protease inhibitors (Bimake, Shanghai, China). Protein concentrations were measured using a BCA protein assay (Beyotime). After denaturation by boiling, the protein samples (20 μg/sample) were subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to PVDF membranes. After blocking with 5% skim milk for 1 h at room temperature, the membranes were probed with primary antibodies at 4°C overnight. After washing three times with TBST, the membranes were incubated with the horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at room temperature, and the antigen-antibody reaction was visualized using an ECL chemiluminescence detection kit (Beyotime).
Finally, images were captured using a gel imager system (ImageQuant Las4000 mini; GE). GAPDH served as the internal control. The specific antibodies used in this article are listed in Supplementary Table S3.
Histology and immunohistochemistry
Micromasses, adherent cells, or mouse limbs were harvested and fixed with 4% paraformaldehyde for 24 h at room temperature. After dehydration with gradient alcohol, micromasses or mouse limbs were embedded in paraffin. Then, the embedded samples were processed for paraffin sectioning at a 4 μm thickness. For Alcian blue staining, the hydrated slices or fixed cells were stained with Alcian blue solutions (pH = 2.5) for 30 min. After washing with PBS for 15 min, photographs were captured by fluorescence microscopy (BX63; Olympus). For quantitative analysis, we added 500 μL of dimethyl sulfoxide (DMSO) to elute the Alcian blue-stained adherent cells, and the absorbance at 650 nm was measured with a microplate reader. For immunohistochemistry, heat-mediated antigen retrieval was performed with citrate buffer, and the hydrated slices were incubated with hydrogen peroxide solution for 20 min to remove the endogenous peroxidase activity.
Subsequently, sections were blocked with 5% bovine serum albumin (BSA) in PBS at room temperature for 30 min, followed by incubation with a primary antibody overnight at 4°C. After washing three times with PBS, sections were incubated with species-matched secondary antibodies for 1 h. Detection was conducted with a DAB Kit (ZSGB-BIO) according to the manufacturer's protocols. For immunofluorescence staining, sections were incubated with species-matched Alexa Fluor 555 for 1 h, and the nuclei were counterstained with DAPI (Cell Signaling Technology, Danvers, MA) for 5 min. The specific antibodies used in this article are listed in Supplementary Table S3.
RNA sequencing
Total RNA was isolated from control or YTHDF1-overexpressing ATDC5 cells after 7 days of chondrogenesis as described above. RNA quality was assessed by the Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA) and verified using RNase-free agarose gel electrophoresis. After total RNA was extracted, eukaryotic mRNA was enriched by Oligo(dT) beads. Then, the enriched mRNA was fragmented into short fragments using the fragmentation buffer and reverse transcribed into cDNA using the NEBNext Ultra RNA Library Prep Kit for Illumina (NEB#7530; New England Biolabs, Ipswich, MA). The purified double-stranded cDNA fragments were end-repaired, subjected to base addition, and ligated to Illumina sequencing adapters. The ligation reaction was purified with AMPure XP Beads (1.0 × ). Ligated fragments were subjected to size selection by agarose gel electrophoresis and PCR amplification. The resulting cDNA library was sequenced using an Illumina Novaseq6000 by Gene Denovo Biotechnology Co. (Guangzhou, China).
Genes with a P value <0.05 and absolute fold change ≥1.8 were considered differentially expressed genes (DEGs), and bioinformatic analysis was performed by the Gene Denovo Biotechnology Co.
Statistical analysis
All quantitative data shown were obtained from independent experiments that were performed at least in triplicate. Histogram data are presented as the mean ± standard deviation and were calculated using SPSS software (version 20.0), where P < 0.05 was considered to indicate statistical significance. Comparisons between two different experimental groups were made using a two-tailed independent Student's t-test. Data from more than two groups were analyzed by one-way analysis of variance followed by Tukey's post hoc test.
Results
The expression of YTHDF1 protein correlates with chondrogenesis in vitro and in vivo
We first explored whether the YTHDF1 protein was expressed in cells producing cartilage matrix. Human chondrosarcoma cell lines SW1353 and Hs819.T, have been reported to produce cartilage matrix [27]. Our results revealed that the YTHDF1 protein was expressed in both Hs819. T and SW1353 cells (Fig. 1a). YTHDF1 protein was also detected in mouse growth plate chondrocytes and the mouse chondrogenic cell line ATDC5 (Fig. 1b). These results revealed that YTHDF1 protein was expressed in some cells producing cartilage matrix. Then, we observed the expression of YTHDF1 protein during the chondrogenic differentiation of hBMSCs. Our results demonstrated that the expression of YTHDF1 protein gradually increased in hBMSCs undergoing chondrogenic differentiation (Fig. 1c). These results suggest that the YTHDF1 protein correlates with chondrogenesis in vitro.
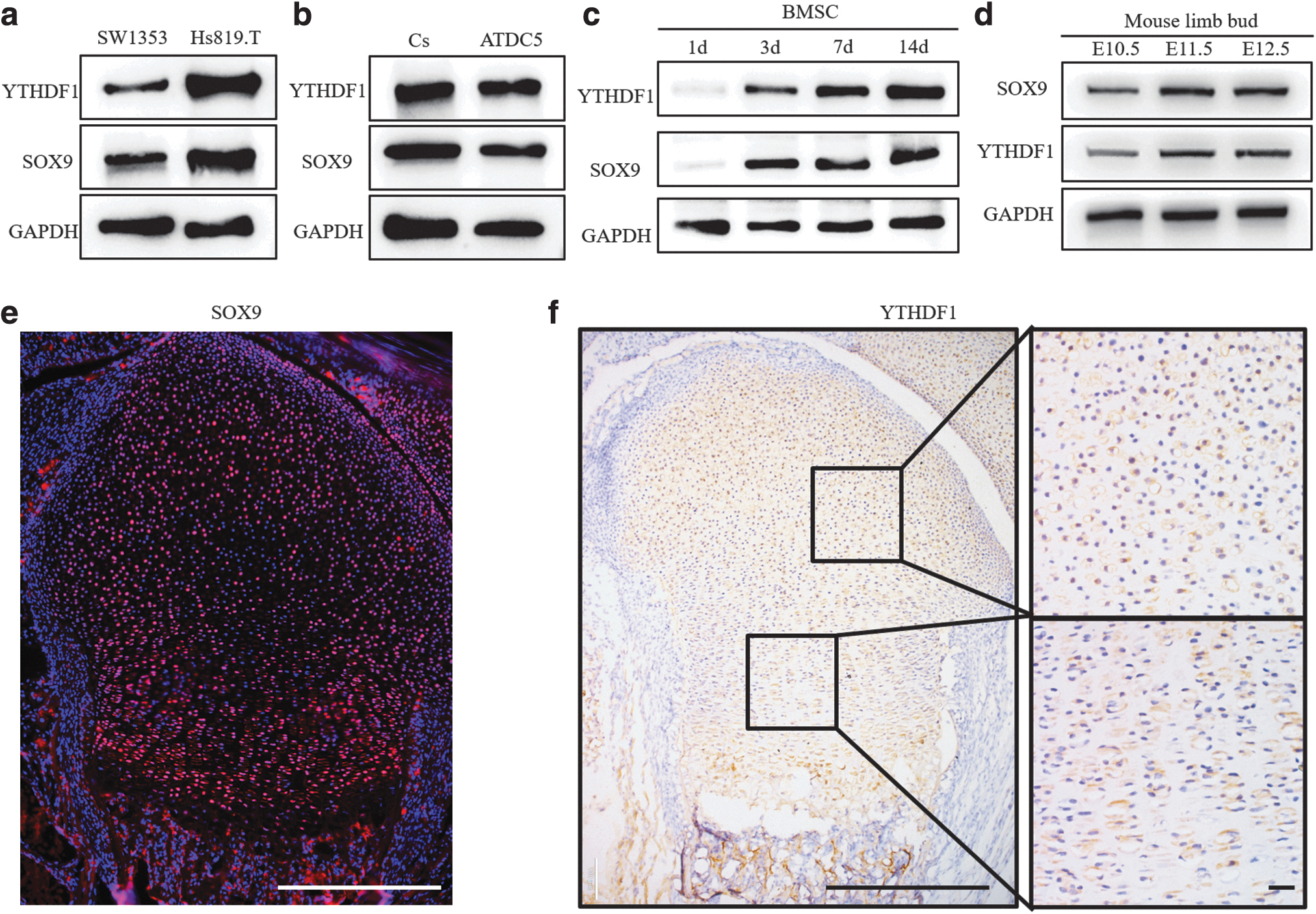
The expression of YTHDF1 protein correlates with chondrogenesis in vitro and in vivo.
To further examine whether the expression of YTHDF1 protein correlated with chondrogenesis in vivo, we examined the YTHDF1 protein level in mouse embryonic limb buds and growth plate cartilage of newborn mice. From embryonic day (E) 10.5 to 12.5, mouse limb bud mesenchymal cells are undergoing chondrogenesis to form cartilage primordia [6]. From the results of western blotting (WB), compared with E10.5 limb buds, we found that both the YTHDF1 and SOX9 proteins were increased in E11.5 and E12.5 limb buds (Fig. 1d), which indicated that YTHDF1 expression correlated with the expression of SOX9 in the early embryonic limb buds. As shown in Figure 1e and f, YTHDF1 was widely expressed in the resting zone and proliferative zone of growth plate cartilage coincident with the expression of SOX9 (Fig. 1e, f). Our data suggested that YTHDF1 expression correlates with chondrogenesis in the early embryonic limb buds and perinatal growth plate.
YTDHF1 promotes the chondrogenic differentiation of hBMSCs
To investigate the role of YTHDF1 in the chondrogenic differentiation of hBMSCs, we stably overexpressed and knocked down YTHDF1 protein in hBMSCs using lentivirus. After 7 days of chondrogenic differentiation, the diameter of the cartilage pellet was measured. Compared with the empty control group, the induced cartilage tissues were larger, and the diameter of the spheres was significantly increased when YTHDF1 was overexpressed (Fig. 2a, b). Subsequently, we tested the expression of genes related to chondrogenic differentiation. From the qPCR results, we found that the expression levels of SOX9, COL2A1, and ACAN mRNA were significantly upregulated in the YTHDF1 overexpression group (Fig. 2c). Consistently, the protein levels of SOX9, COL2A1, and ACAN were increased when YTHDF1 was overexpressed (Fig. 2d), and the immunohistochemical staining results of COL2A1 and ACAN showed a consistent trend (Fig. 2e, f).

YTHDF1 promotes the chondrogenic differentiation after hBMSCs were induced for 7 days. The cartilage pellet
Therefore, these results indicated that overexpression of YTHDF1 promoted the chondrogenic differentiation of hBMSCs.
To further investigate whether chondrogenic differentiation of hBMSCs was mediated by the YTHDF1 protein, we utilized shRNA against YTHDF1 to inhibit the expression of the YTHDF1 gene. As shown in Figure 2g and h, after silencing YTHDF1 protein expression, the induced cartilage tissues were smaller, and the diameter of the cartilage pellet was significantly reduced. We used qPCR and WB to evaluate the effects of YTHDF1 protein on the regulation of cartilage gene expression. Our results demonstrated that transduction of hBMSCs with shRNA against YTHDF1 significantly downregulated the mRNA and protein levels of COL2A1, SOX9, and ACAN (Fig. 2i, j). In addition, decreased accumulation of collagen type II and aggrecan, as determined by immunohistochemical staining, was observed in cartilage tissue with YTHDF1 knockdown compared with the control group (Fig. 2k, l). Therefore, our results indicated that YTDHF1 could promote the chondrogenic differentiation of hBMSCs.
YTDHF1 promotes the chondrogenic differentiation of murine ATDC5 and C3H10T1/2 cell lines
Subsequently, we further explored the role of YTHDF1 in chondrogenic differentiation in two murine cell lines, ATDC5 and C3H10T1/2, both of which are common choices for studying chondrogenic differentiation in vitro. ATDC5 and C3H10T1/2 cells were transduced with a lentivirus to stably overexpress and knock down the murine YTHDF1 protein.
Alcian blue staining showed more abundant cartilage matrix in the YTHDF1 protein overexpression group after adherent ATDC5 cells underwent induction of chondrogenic differentiation for 7 days (Fig. 3a, b). Then, qPCR and WB were used to evaluate the role of YTHDF1 overexpression on the regulation of cartilage gene expression. As shown in Figure 3c, lentivirus-transduced ATDC5 cells overexpressing YTHDF1 lentivirus significantly upregulated the mRNA levels of Col2a1, Sox9, and Acan. Correspondingly, the protein level of SOX9 was increased when YTHDF1 was overexpressed (Fig. 3d).
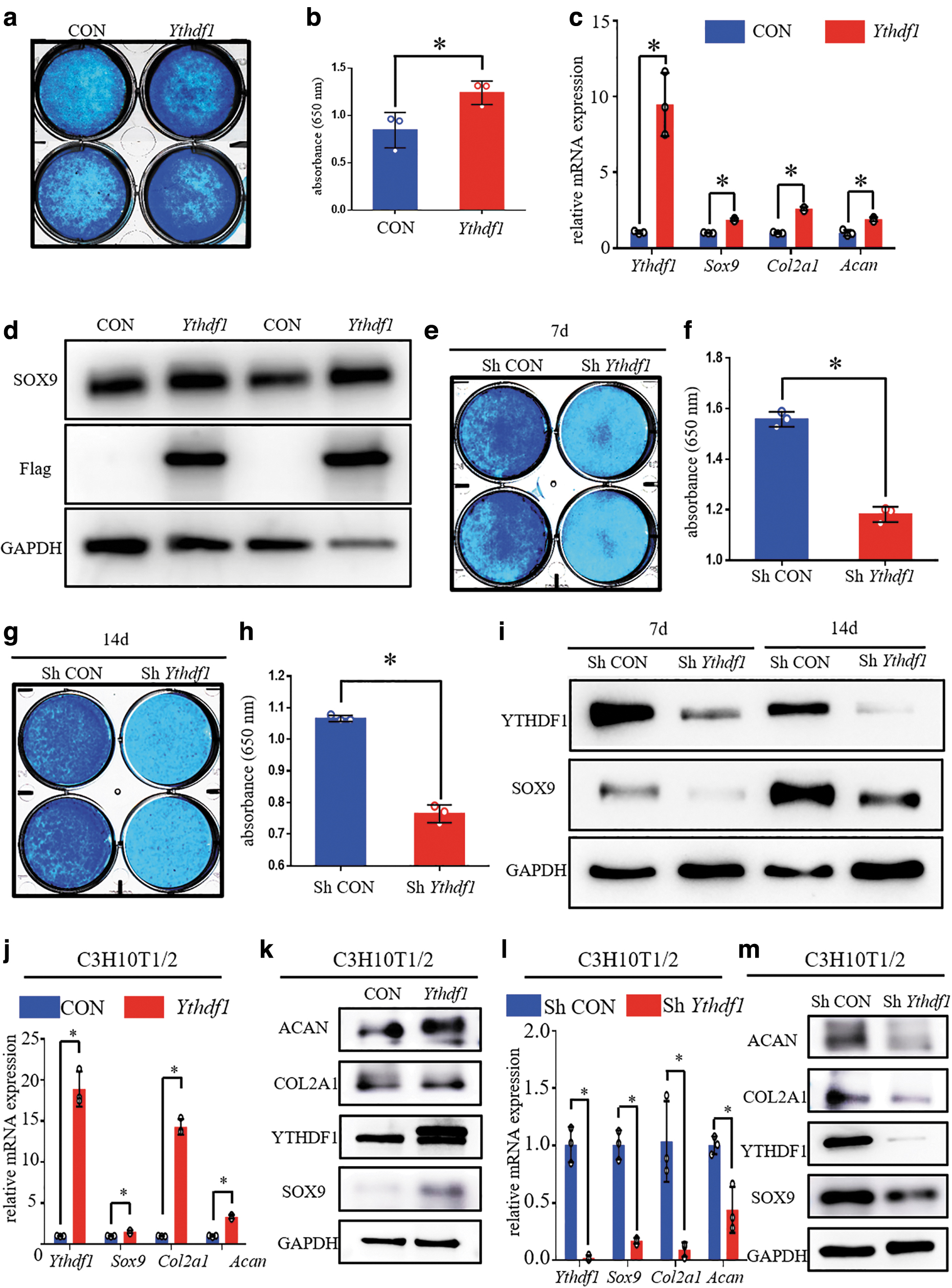
YTDHF1 promotes the chondrogenic differentiation of murine ATDC5 and C3H10T1/2 cell lines. After induction of chondrogenic differentiation for 7 days,
In addition, we also found that the transduction of ATDC5 cells with shRNA against Ythdf1 dramatically disturbed the synthesis of cartilage matrix after adherent ATDC5 cells underwent induction of chondrogenic differentiation for 7 and 14 days (Fig. 3e–h). After chondrogenic differentiation was induced for 7 and 14 days, the expression levels of Col2a1, Sox9, and Acan mRNAs were decreased when YTHDF1 was knocked down in ATDC5 cells (Supplementary Fig. S1a, b). Consistently, transduction of ATDC5 cells with shRNA against Ythdf1 dramatically reduced the protein level of SOX9 regardless of whether chondrogenic differentiation was induced for 7 or 14 days (Fig. 3i).
Subsequently, we also investigated the role of YTHDF1 in the chondrogenic differentiation of the C3H10T1/2 cell line. As shown in Figure 3j and k, we observed that YTHDF1 overexpression increased the mRNA and protein levels of SOX9, COL2A1, and ACAN (Fig. 3j, k). Correspondingly, both the mRNA and protein levels of SOX9, COL2A1, and ACAN were significantly reduced in the knockdown group after C3H10T1/2 cells underwent induction of chondrogenic differentiation for 7 days (Fig. 3l, m).
RNA sequencing revealed that YTHDF1 regulates the catabolism and anabolism of the cartilage matrix
To identify the key factors in chondrogenesis that were regulated by YTHDF1, after 14 days of chondrogenic differentiation induction, the total RNA was obtained from ATDC5 cells transfected with control or overexpression of Ythdf1 lentivirus for RNA sequencing. Three biological replicates were obtained from each group, and the correlation coefficient within the same group was significantly higher than the correlation coefficient between the two groups (Fig. 4a). A total of 914 protein-encoding DEGs were identified (Fig. 4b). Among these DEGs, 316 DEGs were downregulated and 598 DEGs were upregulated in the YTHDF1 overexpression group (Fig. 4b). Subsequently, the up- and downregulated genes were subjected to Gene Ontology (GO) enrichment analysis. We found that changes in increased gene expression were enriched for the collagen extracellular matrix and cell differentiation in the GO analysis (Fig. 4c). However, in downregulated genes, terms related to extracellular matrix and cell differentiation were not significantly enriched (Fig. 4d).

RNA-seq revealed that YTHDF1 regulated the catabolism and anabolism of the cartilage matrix.
From the RNA-seq results, we found that YTHDF1 overexpression upregulated the expression of Sox6, Sox8, and Sox9, which are crucial transcription factors that are known to participate in cartilage matrix synthesis [28,29] (Fig. 4e). Indeed, including Col2a1 and Acan, many other extracellular collagens were also upregulated in the YTHDF1 overexpression group (Fig. 4e). However, no difference was observed in the expression of Col10a1, a marker for hypertrophic chondrocytes, between the two groups (Fig. 4e). In addition, we found that some catabolic enzymes were downregulated when YTHDF1 was overexpressed (Fig. 4f). Therefore, our RNA sequencing revealed that YTHDF1 promoted anabolism and inhibited the catabolism of the cartilage matrix.
YTHDF1 activates the Wnt/β-catenin signaling pathway during chondrogenic differentiation
To identify the signaling pathways in chondrogenesis that were regulated by YTHDF1, the up- and downregulated genes were subjected to the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. We found that changes in increased genes were enriched for the Wnt/β-catenin signaling pathway in KEGG pathway mapping (Fig. 5a). However, for the reduced genes, KEGG pathway analysis did not identify the Wnt/β-catenin signaling pathway (Fig. 5b). Because the Wnt/β-catenin signaling is of great importance for chondrogenic differentiation, we then focused on studying Wnt/β-catenin signaling. According to the results of RNA sequencing, we found that key genes related to Wnt/β-catenin signaling were almost all upregulated, except for Smad3, Nfadc2, and Ccnd1 (Fig. 5c). Then, qPCR was performed to verify the changes in the expression of several key β-catenin pathway downstream genes.
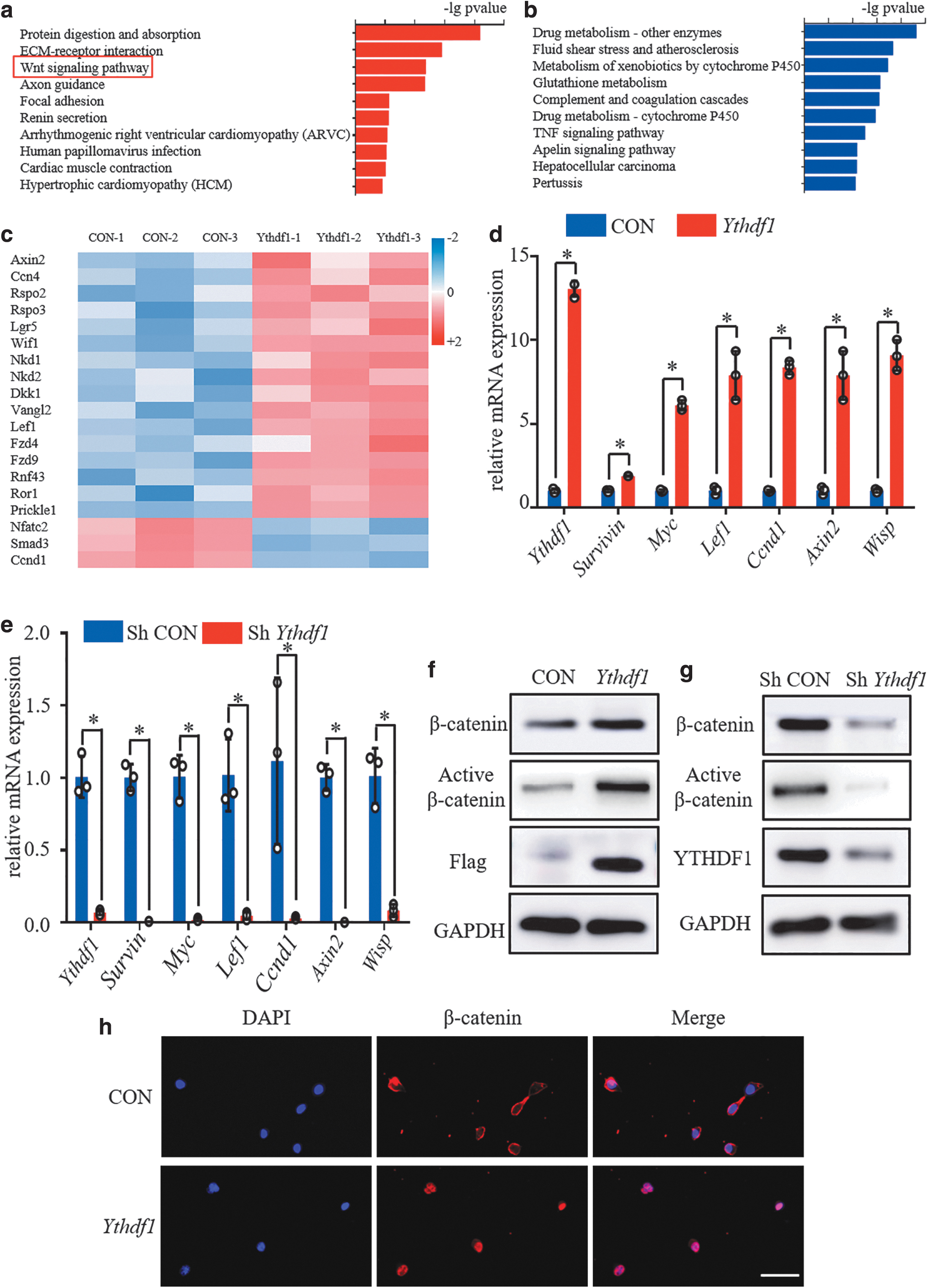
During the chondrogenic differentiation of ATDC5 cells, YTHDF1 activates the Wnt/β-catenin signaling pathway.
The mRNA expression levels of Survivin, Myc, Lef1, Ccnd1, Axin2, and Wisp were all increased in ATDC5 cells overexpressing YTHDF1 (Fig. 5d). Furthermore, in ATDC5 cells with low YTHDF1 expression, these key target genes of Wnt/β-catenin signaling were all dramatically downregulated (Fig. 5e). We then asked whether YTHDF1 regulated Wnt/β-catenin signaling by influencing the expression of the β-catenin protein. By performing WB assays on ATDC5 cells undergoing chondrogenesis, we observed that YTHDF1 overexpression effectively increased the levels of total and nonphosphorylated active β-catenin proteins, while silencing YTHDF1 protein expression reduced the levels of total and nonphosphorylated active β-catenin protein (Fig. 5f, g). Moreover, more nuclear accumulation of β-catenin protein was noted in YTHDF1-overexpressing ATDC5 cells undergoing chondrogenesis (Fig. 5h).
Next, we determined whether YTHDF1 regulated β-catenin signaling during chondrogenic differentiation of hBMSCs and C3H10T1/2 cells. As shown in Supplementary Fig. S2a and c, C3H10T1/2 cells overexpressing YTHDF1 expressed higher levels of β-catenin signaling downstream genes and accumulated more total and active β-catenin proteins after induction of chondrogenic differentiation for 7 days (Supplementary Fig. S2a, c). Meanwhile, silencing the expression of YTHDF1 in C3H10T1/2 cells inhibited β-catenin pathway downstream gene expression and resulted in fewer total and active β-catenin protein residues (Supplementary Fig. S2b, d). Consistently, in cartilage micromasses derived from hBMSCs, we found that YTHDF1 positively regulated the expression of β-catenin pathway downstream genes (Supplementary Fig. S2e, f). Furthermore, WB and immunohistochemical staining assays both showed a positive correlation between the expression of YTHDF1 and the accumulation of total β-catenin protein in induced cartilage tissue (Supplementary Fig. S2g–j).
The level of nonphosphorylated active β-catenin was also found to be increased upon YTHDF1 overexpression and decreased upon YTHDF1 knockdown (Supplementary Fig. S2g, h). Our results revealed that YTHDF1 could activate the Wnt/β-catenin signaling pathway during chondrogenic differentiation.
YTHDF1 enhances β-catenin protein stability
Next, we set out to explore the molecular mechanism by which YTHDF1 overexpression activated the Wnt/β-catenin signaling pathway during chondrogenic differentiation. We first examined the effect of YTHDF1 on β-catenin mRNA expression and did not observe any increase in β-catenin mRNA levels in ATDC5 cells after YTHDF1 overexpression (Fig. 6a). To test whether YTHDF1 expression affected the β-catenin mRNA stability, the ATDC5 cells were treated with actinomycin D to determine the RNA half-life, and YTHDF1 overexpression did not affect the half-life of β-catenin mRNA (Fig. 6b).

YTHDF1 enhances β-catenin protein stability.
These results of β-catenin mRNA suggest that YTHDF1 may play a role in the posttranscriptional regulation of β-catenin. To test the possibility that YTHDF1 overexpression affected the β-catenin protein stability, we treated ATDC5 cells (with or without YTHDF1 overexpression) with 10 μg/mL of cycloheximide to inhibit protein synthesis and we found that the β-catenin protein degradation rate was decreased after YTHDF1 overexpression (Fig. 6c). In addition, control and YTHDF1 overexpression cells were treated with 5 μg/mL of MG132 for 3 h to inhibit proteasome activity. MG132 treatment was able to distinctly attenuate the YTHDF1 overexpression-mediated increase in the β-catenin protein level (Fig. 6d). Therefore, our results revealed that YTHDF1 enhances β-catenin protein stability.
YTHDF1 promotes chondrogenic differentiation by activating the Wnt/β-catenin signaling pathway
We first observed the effects of the β-catenin signaling inhibitor (ICG-1) and β-catenin signaling activator (BML-284 HCL) on chondrogenic differentiation. The expression of β-catenin downstream genes was reduced by ICG-1 treatment and increased by BML-284 HCL treatment (Supplementary Fig. S3a, c). After wild-type ATDC5 and C3H10T1/2 cells underwent induction of chondrogenic differentiation for 7 days, the mRNA expression levels of Sox9, Col2a1, and Acan were all decreased in cells with ICG-1 treatment (Supplementary Fig. S3a, c). In addition, ICG-1 treatment downregulated the expressions of SOX9 and COL2A1 proteins (Supplementary Fig. S3b, d). Consistently, BML-284 HCL significantly upregulated the mRNA levels of Sox9, Col2a1, and Acan, and increased the protein levels of SOX9 and COL2A1 (Supplementary Fig. S3e–h).
To further clarify whether YTHDF1-regulated chondrogenic differentiation depended on the Wnt/β-catenin pathway, we added ICG-1 or BML-284 HCL when YTHDF1 was overexpressed or knocked down in ATDC5 and C3H10T1/2 cells. As expected, ICG1 attenuated the YTHDF1 overexpression-mediated increase in the expression of β-catenin downstream genes, and BML-284 HCL rescued the inhibitory effect of YTHDF1 knockdown on the β-catenin signaling pathway (Supplementary Fig. S4a–d).
For ATDC5 cells undergoing chondrogenesis for 7 days, ICG-1 treatment was able to obviously attenuate the YTHDF1 overexpression-mediated increase in the synthesis of cartilage matrix (Fig. 7a, b). In addition, BML-284 HCL rescued the inhibitory effect of YTHDF1 knockdown on cartilage matrix synthesis in ATDC5 cells (Fig. 7c, d). For qPCR and WB analyses of chondrogenesis biomarker expression in ATDC5 cells that underwent 7 days of chondrogenic differentiation, increased mRNA and protein levels of Sox9, Col2a1, and Acan mediated by YTHDF1 overexpression were reversed by ICG-1 (Fig. 7e, f). Meanwhile, YTHDF1 knockdown downregulated the mRNA expression of Sox9, Col2a1, and Acan, whereas after the addition of BML-284 HCL, upregulation of these mRNAs was observed (Fig. 7g). For WB assays, YTHDF1 knockdown decreased the protein levels of total β-catenin, active β-catenin, SOX9, and COL2A1, whereas BML-284 HCL treatment reversed this effect (Fig. 7h).
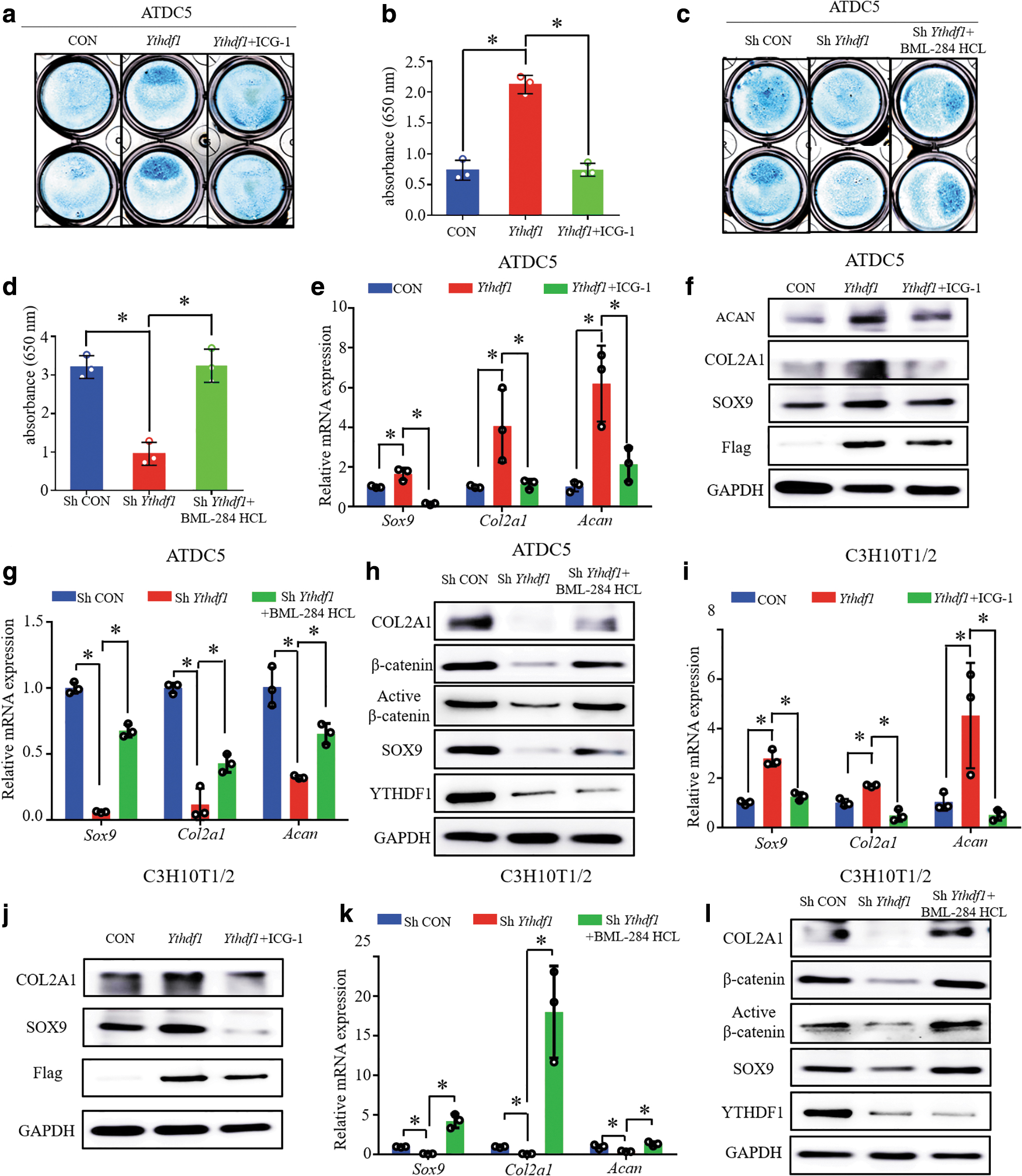
YTHDF1 regulates chondrogenic differentiation through the Wnt/β-catenin signaling pathway. After ATDC5 cells were induced to chondrogenic differentiation for 7 days,
We further determined the effect of the Wnt/β-catenin pathway on YTHDF1-mediated chondrogenic differentiation of C3H10T1/2 cells. From the results of qPCR assays, we observed that ICG1 attenuated the YTHDF1 overexpression-mediated increase in the mRNA levels of Sox9, Col2a1, and Acan in C3H10T1/2 cells that underwent 7 days of chondrogenic differentiation (Fig. 7i). As shown by the WB assays, YTHDF1 overexpression increased the protein levels of SOX9 and COL2A1, whereas ICG-1 treatment reversed this effect (Fig. 7j). Consistently, BML-284 HCL rescued the YTHDF1 deficiency-impaired β-catenin activity and chondrogenesis of C3H10T1/2 cells (Fig. 7k, l).
Discussion
In the present study, we first investigated the effects of YTHDF1 on chondrogenic differentiation. In previous studies, a variety of cell models from different species possessed potential for chondrogenic differentiation. Of the human-derived cells, hBMSCs are an effective cell model in which chondrogenic differentiation induces the formation of cartilage pellets, which have also been extensively studied by us [24,25]. Of the mouse-derived cells, ATDC5 is the most common chondrogenic cell line and a good choice for researching cartilage development [30,31]. In addition, C3H10T1/2, the murine immortalized MSC line, has been confirmed to have multilineage potential [32].
Many studies have revealed that C3H10T1/2 cells were induced to chondrogenesis in micromass cultures, and thus became a well-known in vitro model of chondrogenesis [33,34]. Therefore, in this study, we used these three cell models from humans and mice to fully illustrate the role of YTHDF1 in the regulation of chondrogenic differentiation.
m6A mRNA methylation has been reported to be a crucial modulator in regulating the pluripotency of stem cells. Consistent with our study, many reports have also demonstrated that m6A can promote stem cell differentiation [35 –37]. However, other studies have shown that m6A modifications maintain the pluripotency of stem cells [38,39]. Therefore, the impact of m6A modification in stem cell biology is complex, and m6A may be involved in regulating multiple steps of stem cell differentiation, which requires further research in the future.
Reduced synthesis and increased degradation of cartilage matrix are thought to play a key role in the pathogenesis and progression of OA [40]. Some studies have explored the role of writer and eraser proteins of m6A in the progression of OA. Using the ATDC5 cell model, one study found that the expression of METTL3 was increased in IL-1β-treated cells and that METTL3 promoted the degradation of the cartilage matrix under inflammatory stimulation [41]. However, another study found that the expression of METTL3 was reduced in IL-1β-treated SW1353 cells [42]. The m6A eraser protein FTO was reported to be significantly downregulated in osteoarthritic cartilage and may inhibit the progression of OA [43]. However, unlike the writer and eraser proteins of m6A, the role of reader proteins in osteoarthritis remains unstudied.
In our study, we found that YTHDF1 promoted anabolism and inhibited the catabolism of the cartilage matrix, which indicated that YTHDF1 may play a protective role in the onset or progression of OA. Therefore, in the pathogenesis and progression of OA, the role of molecules that regulate m6A mRNA methylation modification (especially reader proteins) requires further exploration.
Some researchers have found that inhibiting Wnt/β-catenin signaling impaired the proliferation and differentiation of chondrocytes and skeletal formation in mice, suggesting that this pathway plays an important role in chondrogenic differentiation [44]. In the current study, we found that chondrogenesis of wild-type ATDC5 and C3H10T1/2 cells was reduced upon β-catenin inhibitor treatment and increased upon β-catenin activator treatment. Moreover, inhibition of β-catenin activity in ATDC5 and C3H10T1/2 cells reversed the promoting effect of YTHDF1 on chondrogenic differentiation. Our results revealed that the Wnt/β-catenin signaling pathway positively regulated chondrogenic differentiation. A study carried out in ATDC5 and C3H10T1/2 cells has also reported that Wnt/β-catenin signaling activation promoted chondrogenic differentiation [45], which is consistent with our findings. However, another study on the chondrogenesis of hBMSCs derived from OA patients demonstrated that activation of the Wnt/β-catenin signaling pathway inhibited chondrogenic differentiation [46].
These inconsistencies suggest that the Wnt/β-catenin signaling pathway may be able to regulate chondrogenesis through different molecular mechanisms; this speculation requires further exploration. Several in vitro studies carried out in other types of cells, such as primary chondrocytes, human synovial fluid MSCs, and rat BMSCs, showed that chondrogenic differentiation was significantly attenuated when β-catenin was inhibited, which was also in agreement with our findings [47,48]. Another group found that knockout (KO) of Ptch1 in limb bud MSCs promoted chondrogenic differentiation, which contributed to enhanced β-catenin activation [49]. Maruyama et al. constructed an Axin2 KO mouse model to activate the Wnt/β-catenin pathway in vivo and found that MSCs in the skull of Axin2 KO mice underwent chondrogenic differentiation to form chondrocytes, but did not directly differentiate into osteoblasts [50]. These in vivo experiments suggested that activating Wnt/β-catenin signaling could promote chondrogenic differentiation of MSCs during skeletal development, which was consistent with our results.
In this study, we used a TGFβ3-containing chondrogenic induction medium. However, there are other known growth factors, such as BMP2, that can be used for chondrogenic differentiation induction [51]. We investigated whether YTHDF1 affected chondrogenic differentiation when BMP2 instead of TGFβ3 was added to the chondrogenic differentiation medium. After chondrogenic differentiation for 7 days, the mRNA levels of Sox9, Col2A1, and Acan were significantly reduced in C3H10T1/2 cells with YTHDF1 knockdown (Supplementary Fig. S5a). Moreover, YTHDF1 knockdown decreased the protein levels of SOX9 and COL2A1 in differentiated C3H10T1/2 cells (Supplementary Fig. S5b). Our results suggest that YTHDF1 may also play a catalytic role in BMP2-induced chondrogenic differentiation.
Our study has some limitations that we hope to address. First, we only demonstrated that YTHDF1 promoted chondrogenic differentiation in vitro. Whether YTHDF1 regulates chondrogenic differentiation of limb bud MSCs during skeletal development is still unknown. We will construct transgenic mice with generalized or limb bud conditional KO of the Ythdf1 gene to elucidate its role in chondrogenic differentiation in vivo. Second, the molecular mechanisms underlying the regulation of Wnt/β-catenin signaling by YTHDF1 during chondrogenic differentiation remain unknown. Two other studies have demonstrated that the YTHDF1 protein activated the Wnt/β-catenin pathway by promoting the translation of the Wnt signaling effector TCF4 and the Wnt receptor FZD7 [19,52]. Eventually, we speculate that YTHDF1 may activate the β-catenin pathway by promoting the translation of few or even many m6A-modified mRNAs during chondrogenic differentiation, and further methylated RNA immunoprecipitation and sequencing assays may help to comprehensively elucidate which m6A-modified mRNAs are involved in this mechanism.
Conclusions
In the current study, for the first time, we elucidated the role of the m6A reader protein YTHDF1 in the regulation of chondrogenic differentiation. The expression of YTHDF1 was significantly increased during chondrogenic differentiation of hBMSCs. Furthermore, RNA-seq and molecular biology experiments found that YTHDF1 activated the Wnt/β-catenin signaling pathway during chondrogenic differentiation of hBMSCs, ATDC5 cells, and C3H10T1/2 cells. Finally, inhibiting or activating β-catenin activity can reverse the effect of overexpressing or knocking down YTHDF1 on chondrogenic differentiation. Therefore, our data indicate that YTHDF1 enhances chondrogenic differentiation by activating the Wnt/β-catenin signaling pathway.
Footnotes
Data Availability
The primary data presented in this study are available on request from the corresponding author.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 82202652); the Fundamental Research Funds for the Central Universities (No. 2042022kf1097); the Guangdong Natural Science Foundation (No. 2021A1515012619); the Sun Yat-sen Scholarship for Young Scientist for W.G.; and the Science and Technology Program of Guangzhou, China (No. 202102010159).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.