
Abstract
Peritoneal fibrosis is a critical sequela that limits the application of peritoneal dialysis (PD). This study explored the role and mechanism of bone marrow mesenchymal stem cell-derived exosomes (BMSC-Exos) in preventing PD-associated peritoneal injury. C57BL/6 mice were randomized into three groups: a control (saline), peritoneal injury [2.5% glucose peritoneal dialysate + lipopolysaccharide (LPS)], and peritoneal injury + exosome group. After 6 weeks, mice were dissected, and the parietal peritoneum was collected. The level of peritoneal structural and functional damage was assessed. Additionally, transcriptome analysis of the peritoneum and miRNA sequencing on BMSC-Exos were performed. The parietal peritoneum had significantly thickened, and peritoneal function was impaired in the peritoneal injury group. Peritoneal structural and functional damage was significantly reduced after exosome treatment, while peritoneal inflammation, fibrosis, angiogenesis, and mesothelial damage significantly increased. Transcriptomic analysis showed that the BMSC-Exos affected the cell cycle process, cell differentiation, and inflammatory response regulation. Significant pathways in the exosome group were enriched by inflammation, immune response, and cell differentiation, which constitute a molecular network that regulates the peritoneal protective mechanism. Additionally, inflammatory factors (TNF-α, IL-1β), fibrosis markers (α-SMA, collagen-III, fibronectin), profibrotic cytokines (TGF-β1), and angiogenesis-related factor (VEGF) were downregulated at the mRNA and protein levels through BMSC-Exos treatment. BMSC-Exos treatment can prevent peritoneal injury by inhibiting peritoneal fibrosis, inflammation, and angiogenesis, showing a multitarget regulatory effect. Therefore, BMSC-Exos therapy might be a new therapeutic strategy for treating peritoneal injury.
Introduction
Currently, approximately two million people worldwide have end-stage renal disease, and the increasing number of cases each year represents an important health concern [1]. Peritoneal dialysis (PD), an essential therapy for kidney failure, accounts for ∼11% of all modalities of renal replacement treatment worldwide [2]. Recent studies have proven that PD is safe and as efficient as hemodialysis [3,4]. However, peritoneal fibrosis, a serious sequela that limits the application of PD techniques, results from persistent exposure of PD fluids and recurrent episodes of peritonitis. PD induces lasting exposure of the peritoneum to bioincompatible hypertonic dialysis solutions, leading to chronic peritoneal inflammation. Moreover, PD patients are at high risk for infectious peritonitis, which promotes chronic inflammation, peritoneal mesothelial injury, neoangiogenesis, and peritoneal fibrosis. Long-term PD causes the development of structural and functional alterations in the peritoneum, especially peritoneal fibrosis, which is the major contributor to technical failure in PD patients [5 –7].
Methods for clinical prevention of PD-associated peritoneal fibrosis include more biocompatible PD solutions, peritoneal short resting, tamoxifen, heparin, and RAS blockers [5], but their clinical effects are unsatisfactory. Therefore, effective treatment measures for peritoneal fibrosis are lacking. Novel strategies to effectively inhibit peritoneal injury and peritoneal fibrosis are necessary to improve the prognosis of dialysis patients.
Recently, new biological therapies have garnered attention. Mesenchymal stem cell (MSC) therapy alleviates peritoneal injury in animal models by inhibiting TGF-β1 signaling and suppressing inflammation, and may be a potential strategy to prevent peritoneal fibrosis [8,9]. Meanwhile, more and more studies have observed that MSC possess therapeutic capabilities primarily based on their paracrine mechanism [10,11]. The paracrine effects act through the transfer of extracellular vesicles (EVs) (including exosomes and microvesicles) released by MSCs [12]. MSC-derived exosomes (MSC-Exos) are typically 30–150 nm in diameter and cup shaped or round [13]. They are responsible for the horizontal transfer of mRNA, microRNA, and proteins and are integral components of intercellular communication [14]. Exosomes are ideal therapeutic agents owing to their small size, low immunogenicity, long-circulating half-life, good penetration, and good biocompatibility [15].
MSC-Exos could inhibit chronic tissue inflammation and suppress organ fibrosis. Bone marrow mesenchymal stem cell-derived exosomes (BMSC-Exos) were potent at reducing the chronic damage and fibrosis of kidneys in a rat model of chronic kidney disease [16]. BMSC-Exos attenuate rat myocardial fibrosis both in vivo and in vitro through autophagy activation [17]. Human MSC-derived exosomes inhibited liver fibrosis in a mouse model [18,19]. Delivery of BMSC-Exos could relieve atrial fibrillation-induced rat myocardial fibrosis, apoptosis, and inflammation [20]. To the best of our knowledge, no study has focused on the use of MSC-derived exosomes to reduce peritoneal fibrosis progression during PD. We aimed to elucidate the therapeutic effect of MSC-Exos on peritoneal injury.
Materials and Methods
Induction of peritoneal injury and exosome treatment
Eight-week-old male C57BL/6 mice (Siberfo Biotechnology, Beijing, China) were randomly divided into three groups (n = 6 per group): control, peritoneal injury, and exosome. In the control group, mice were subjected to intraperitoneal (i.p.) saline injection with 0.1 mL/g body weight, daily for 6 weeks. In the peritoneal injury group, mice received i.p. injection with 2.5% PDF (0.1 mL/g, daily, Huaren Pharmaceutical, H20113091) and LPS (10 mg/kg, once/5 days; Sigma, L4130) for 6 weeks [21]. In the exosome group, mice received i.p. injection with 2.5% PD fluid and LPS for 6 weeks, and treatment with BMSC-Exos (200 μg/kg/dose) at the end of the fourth (day 28) and fifth (day 35) weeks. After 6 weeks, all mice were sacrificed through cervical dislocation. Subsequently, samples of parietal peritoneum, serum, and PD effluent were collected.
In the experiments described above, the dosage (200 μg/kg) and timing (day 28 and 35) of BMSC-EXos treatment were selected based on the results of previous experiments (Supplementary Fig. S1). Animal experiments were approved and conducted in accordance with the guidelines of the Animal Ethics Committee of Army Medical University (AMUWEC20191679).5
Isolation and identification of BMSC-Exos
BMSCs from normal mice were isolated and cultured using the whole bone marrow culture method. BMSCs were identified through flow cytometry analysis of BMSC markers and the method of induced differentiation into adipocytes and osteocytes. According to the manufacturer's instructions, exosomes from the BMSC supernatant were isolated and purified utilizing the Exosome Concentration Kit (Rengen Biosciences; EXOCCon10–10) and the Exosome Purification Kit (Rengen Biosciences; EXOSEC0.5–5). BMSC-Exos were identified through western blotting analysis of exosome biomarker proteins and transmission electron microscopy Fig. 1A–D.

Identification and in vivo tracking of BMSC-derived exosomes.
Transcriptome sequencing and analysis
RNA-seq analysis was conducted on transcriptome mRNA sequencing (RNA-seq) RNA isolated from mouse parietal peritoneum. According to the manufacturer's protocol, RNA isolation and purification of total samples were performed utilizing TRIzol reagent (Invitrogen). Paired-end (2 × 150 bp) sequencing reads of libraries were conducted on Illumina NovaSeq™ 6000 sequencing platforms. Differentially expressed genes (DEGs) defined as P < 0.05 and fold change (FC) >2 or <0.5 were screened using the DESeq2 (version 1.36.0) [22] and analyzed through Gene Ontology (GO) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses.
miRNA sequencing
RNA samples from the BMSC-Exos were isolated according to the manufacturer's protocol. miRNA profiling was determined using the QIAseq miRNA Library Kit. Three databases: miRbase, piRNAbank, and Rfam, were compared and quantified. The normalization for each gene unique molecular index count was determined by counts per million, which was calculated using the edgeR package [23].
In vivo tracking of BMSC-Exos
For in vivo tracking, BMSC-Exos were stained with DiI dye (Invitrogen; C7001) according to the manufacturer's protocol. PBS or DiI dye-labeled exosomes (DiI-Exos) were administered through i.p. injection in mice with peritoneal injury. After 48 h, the mice were dissected, and the parietal peritoneal tissue was taken for immunofluorescence (IF) staining. The distribution of fluorescence images of DiI-Exos in the mice's peritoneum were acquired at 553 and 570 nm excitation and emission wavelengths under confocal microscopy (SP8; Leica).
Histology, immunohistochemistry, and IF
Paraffin sections of mouse peritoneum were processed for Periodic Acid Schiff (PAS) and Masson staining. Alpha-smooth muscle actin (α-SMA), fibronectin (FN), collagen-III, myeloperoxidase (MPO), F4/80, CD68, CA125, Griffonia simplicifolia lectin 1 (GS1-lectin), and vascular endothelial growth factor (VEGF) were detected through immunohistochemical staining of paraffin sections. E-Cadherin and Ki-67 were detected through IF staining of frozen sections. The positive staining areas (α-SMA, FN, collagen-III, GS1-lectin, and VEGF) and the percentage of positive cells (MPO, F4/80, and CD68) in the submesothelial area, were determined and measured using lumina v.2.20 (Mitani) and ImageJ v.1.53e (NIH) at eight random fields ( × 200). Antibody information is provided in Supplementary Table S1.
Real-time qPCR
The expression of mRNA was assessed by real-time quantitative PCR (RT-qPCR). RNA isolation from parietal peritoneum samples was performed using TRIzol reagent. Reverse transcription of RNA to cDNA was performed using the RT-PCR Kit (TaKaRa). We used the GoTaq® qPCR Kit (Promega) to perform RT-qPCR and CFX ManagerTM v.3.1 (Bio-Rad) to analyze the mRNA levels using the 2−ΔΔCT method. The reference gene GAPDH was used to normalize the PCR results. The sequences of primers are presented in Supplementary Table S2.
Peritoneal equilibration test
The peritoneal permeability and ultrafiltration of mice were measured through a 2-h modified peritoneal equilibration test (PET) on the last day before they were dissected [24]. Two hours after i.p. injection with 3 mL of 7% glucose PD fluid, peritoneal effluents and serum were harvested. The net ultrafiltration volume was calculated as the volume of the peritoneal effluent after 2 h minus the injection volume. We calculated the D/D0 glucose ratio (ratio of dialysate glucose at 2 and 0 h) and the D/P blood urea nitrogen (BUN) ratio (ratio of dialysate BUN to plasma BUN at 2 h) to assess the functional status of peritoneal solute transport.
Statistical analysis
Data are presented as mean ± standard deviations (SD). One-way ANOVA was used for multiple comparisons of parametric variables. The Kruskal–Wallis test was used for multiple comparisons of nonparametric variables. Statistical analysis was conducted by SPSS v.22.0 (IBM).
Results
BMSC-Exos alleviate mouse peritoneal fibrosis induced by 2.5%PDF+LPS
Chronic peritoneal injury is characterized by peritoneal fibrosis, inflammation, and neoangiogenesis [25]. In the process of constructing peritoneal injury mouse model, we found that the peritoneum began to thicken on day 28 with obvious differences on day 42 in the peritoneal injury group (Supplementary Fig. S1A and Fig. 2A, B). Then, we identified the exosomes isolated from BMSCs based on the classic markers (CD9, CD63, and TSG101) and TEM (Fig. 1A–D). To determine the optimal therapeutic dose of BMSC-Exos, we preliminarily proposed three doses of 100, 200, and 400 μg/kg/dose [26,27], and chose day 28 when the peritoneal thickening begins to appear as the treatment time point. The results showed that both 200 and 400 μg/kg/dose could alleviate obvious peritoneal thickening, and no significant differences were noted between the two groups (Supplementary Fig. S1B, C). Next, we aimed at optimizing the timing of BMSC-Exos treatment.

BMSC-derived exosomes alleviate peritoneal fibrosis and protect peritoneal function.
The results suggested that BMSC-Exos can reduce peritoneal thickening at day 28 and 35 (Supplementary Fig. S1D–F). It was further found that 28 days +35 days' treatment group had better effect on alleviating peritoneal thickening than the other two single treatment groups (Supplementary Fig. S1G, H). Therefore, we chose to treat the mice with 200 μg/kg/dose BMSC-Exos at day 28 and 35. We verified that, i.p. injection of BMSC-Exos could significantly inhibit peritoneal thickening in mice (Figs. 1E, F and 2A, B).
Fibrosis is distinguished by the exceeding deposition and remodeling of the extracellular matrix, of which markers include FN and collagen-III [28]. Furthermore, α-SMA is widely used as a marker for myofibroblasts, which drive fibrosis [29]. The protein and mRNA levels of fibrotic markers (FN, collagen-III, and α-SMA) in the exosome group were markedly reduced compared with those in the peritoneal injury group (Fig. 2A, C–H). Moreover, the mRNA level of profibrotic cytokine TGF-β1 decreased significantly after BMSC-Exos treatment (Fig. 2I). These results indicated that BMSC-Exos treatment alleviated PD-related peritoneal fibrosis in mice.
BMSC-Exos improve functional decline of peritoneal membrane
Peritoneal fibrosis can induce membrane functional failure, resulting in patients' discontinuation of PD treatment [30]. The ultrafiltration of the peritoneal membrane in mice was measured using modified PET. The net ultrafiltration volume in the peritoneal injury group was significantly lower compared with that in the control group, suggesting its ultrafiltration function was impaired (Fig. 2J). Additionally, the peritoneum in mice with peritoneal injury showed a high solute transport; that is, the ratio of glucose concentration (D/D0) decreased, and the ratio of urea nitrogen concentration (D/P BUN) increased significantly (Fig. 2K, L). Exosome treatment mitigated the reduced peritoneal ultrafiltration resulting from peritoneal injury and alleviated the rapid solute transport in the peritoneum (Fig. 2J–L).
BMSC-Exos relieve peritoneal inflammation and angiogenesis
Long-term PD causes peritoneal inflammation and neoangiogenesis, which are crucial pathologic mechanisms during the progression of peritoneal fibrosis [31]. Furthermore, macrophages and neutrophils are the main immune cells that invade the peritoneum during peritonitis [32]. We found that BMSC-Exos treatment suppressed macrophage (CD68+ or F4/80+) and neutrophil (MPO+) infiltration in the peritoneum (Fig. 3A–D). Similarly, the mRNA expression of inflammatory factors (TNF-α, IL-1β, and IL-6) in the exosome group decreased significantly from that of the peritoneal injury group (Fig. 3E–G). Likewise, the expression level of angiogenesis markers (GS1-lectin and VEGF) in the peritoneal tissues of mice in the exosome group was significantly lower compared with those in the peritoneal injury group (Fig. 3H–K). These results suggested that BMSC-Exos attenuated peritoneal inflammation and angiogenesis during peritoneal injury.
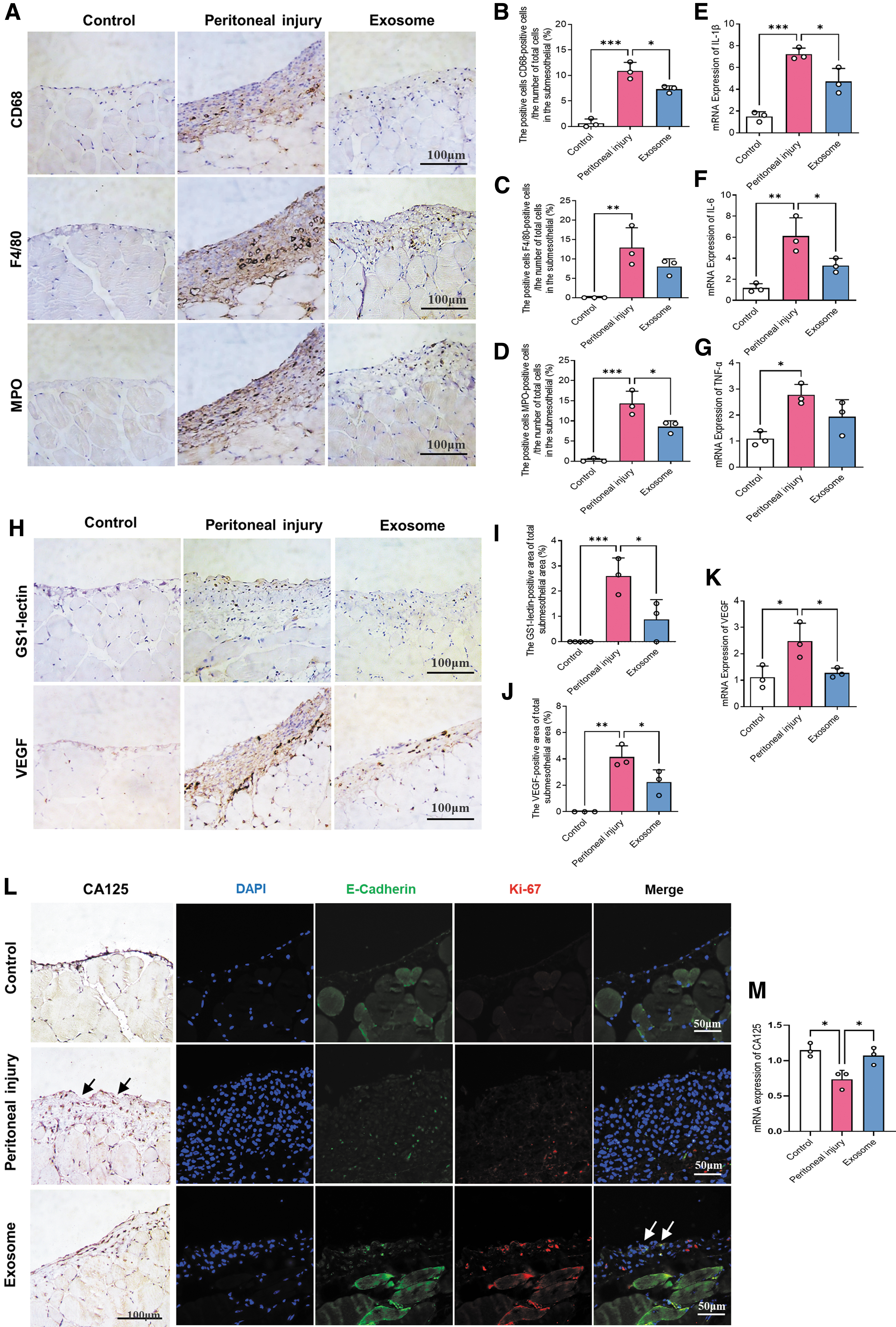
BMSC-Exos relieve peritoneal inflammation, angiogenesis, and the injury of peritoneal mesothelial cells.
BMSC-Exos alleviate peritoneal mesothelial cell injury
Peritoneal mesothelial cells (PMC) are epithelial-like cells lining the entire abdominal cavity and are the first line of defense, playing a dominant role in peritoneal function [33]. Compared with that in the peritoneal injury group, the expression of CA125, a PMC marker in the mesothelial cell monolayer, significantly increased in the exosome group (Fig. 3L, M). The IF costaining of PMC marker (E-cadherin) and cell proliferation marker (Ki-67) showed that the number of proliferative PMC in the exosome group increased significantly compared with that in the peritoneal injury group (Fig. 3L). Therefore, BMSC-Exos treatment might promote the proliferation and repair of PMC, and consequently, alleviate peritoneal mesothelial layer damage.
BMSC-Exos alleviate peritoneal injury by regulating multiple biological effects
We performed in vivo tracking of BMSC-Exos using DiI dye to determine whether BMSC-Exos can actively penetrate the peritoneal mesothelial layer and submesothelial layer to exert protective effects. The fluorescence signal of DiI-Exos was found in both the peritoneal mesothelial layer and the submesothelial layer, especially in PMC (vimentin+) and submesothelial fibroblasts (collagen
Transcriptomics was used to detect the influence of exosomes on the whole-genome expression of the peritoneum to clarify the biological role of exosomes in protecting against peritoneal injury. We performed comparisons to identify the DEGs according to the screening criteria (P < 0.05, FC ≥2 or ≤0.5). Of the 55,401 genes detected, 1,686 upregulated and 5,465 downregulated DEGs were found in the exosome group compared with the peritoneal injury group (Fig. 4C). The heat map showed that among the DEGs coregulated by the peritoneal injury effect and BMSC-Exos effect, most were upregulated in the peritoneal injury group (72.8%), whereas the BMSC-Exos treatment significantly inhibited the expression enhancement of these genes (Fig. 4F). We speculated that BMSC-Exos might restore the mRNA expression of most stimulated genes to baseline levels. Furthermore, the marker genes related to inflammation, fibrosis, cell differentiation, and angiogenesis were modulated after BMSC-Exos treatment, consistent with prior results (Fig. 4G–J).
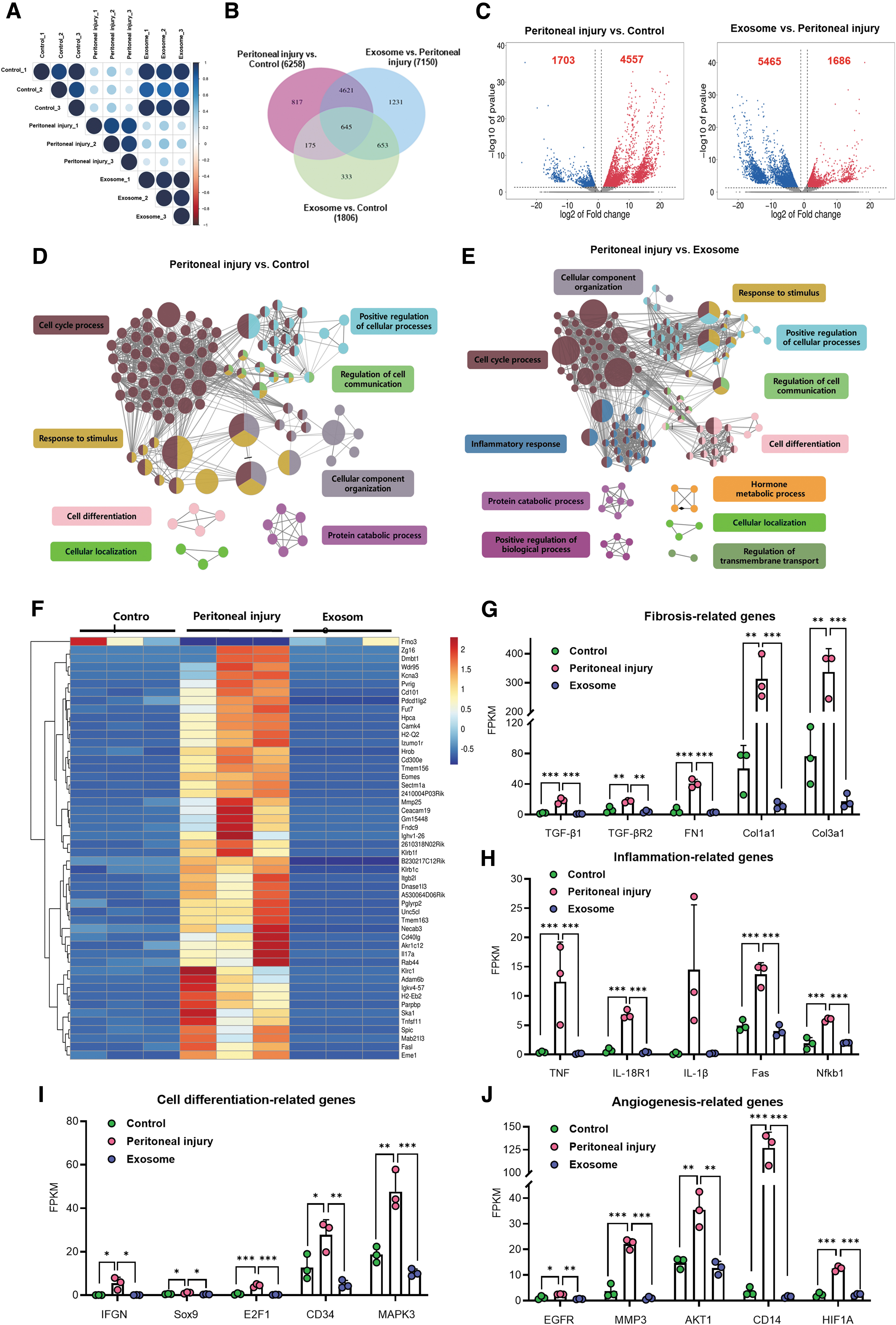
BMSC-derived exosomes alleviate peritoneal injury by regulating multiple biological effects.
GO enrichment analysis showed that the peritoneal injury effect involves numerous biological processes, mainly cell cycle, cell communication, response to stimulus, and cell differentiation (Fig. 4D). Notably, the BMSC-Exos effect could regulate all the above processes. Additionally, exosome treatment enriched different biological process clusters, such as inflammatory response, regulation of transmembrane transport, and hormone metabolic process (Fig. 4E).
KEGG pathway analysis revealed 152 significantly differential pathways between the exosome and peritoneal injury groups. These pathways were mainly enriched in inflammation (eg, NF-κB pathway, tumor necrosis factor pathway, and chemokine signaling pathway), cell differentiation (eg, Rap1, RAS, and PI3K-Akt signaling pathway), and immune response (eg, antigen processing and presentation, and Th17 cell differentiation) (Supplementary Table S3). Moreover, the exosome group's enrichment score on inflammation, immunity, and cell differentiation, among other related molecular pathways, was higher than that in the peritoneal injury group (Supplementary Fig. S2C, D). In this context, we speculated that BMSC-Exos regulated various biological functions of the peritoneum through multiple pathways.
BMSC-Exos protect the peritoneum through miRNA clusters of different functions
Numerous studies have confirmed that exosomes exert various tissue-protective effects through miRNAs [34 –37]. Through miRNA sequencing, we identified 1960 miRNAs contained in BMSC-Exos. Furthermore, these miRNAs have proved to regulate biological processes, including inflammation, fibrosis, angiogenesis, cell proliferation and differentiation, as well as oxidative stress (Table 1). To reflect the functions of these miRNAs further, a miRNA functional network was constructed based on GO analysis (Fig. 5C). The miRNA-GO network identified that miRNA clusters in BMSC-Exos were involved in regulating functions, such as inflammatory response, cell differentiation, cell proliferation, oxidative stress, and angiogenesis (Fig. 5C). Furthermore, KEGG analysis showed the most differentially expressed miRNAs associated with signaling pathways were related to cell survival (Ras, mTOR, and apoptosis), cell differentiation (Wnt and MAPK), and inflammatory response (TNF) (Fig. 5D). Therefore, BMSC-Exos could present various protective effects during peritoneal injury through a variety of miRNAs with different functions.
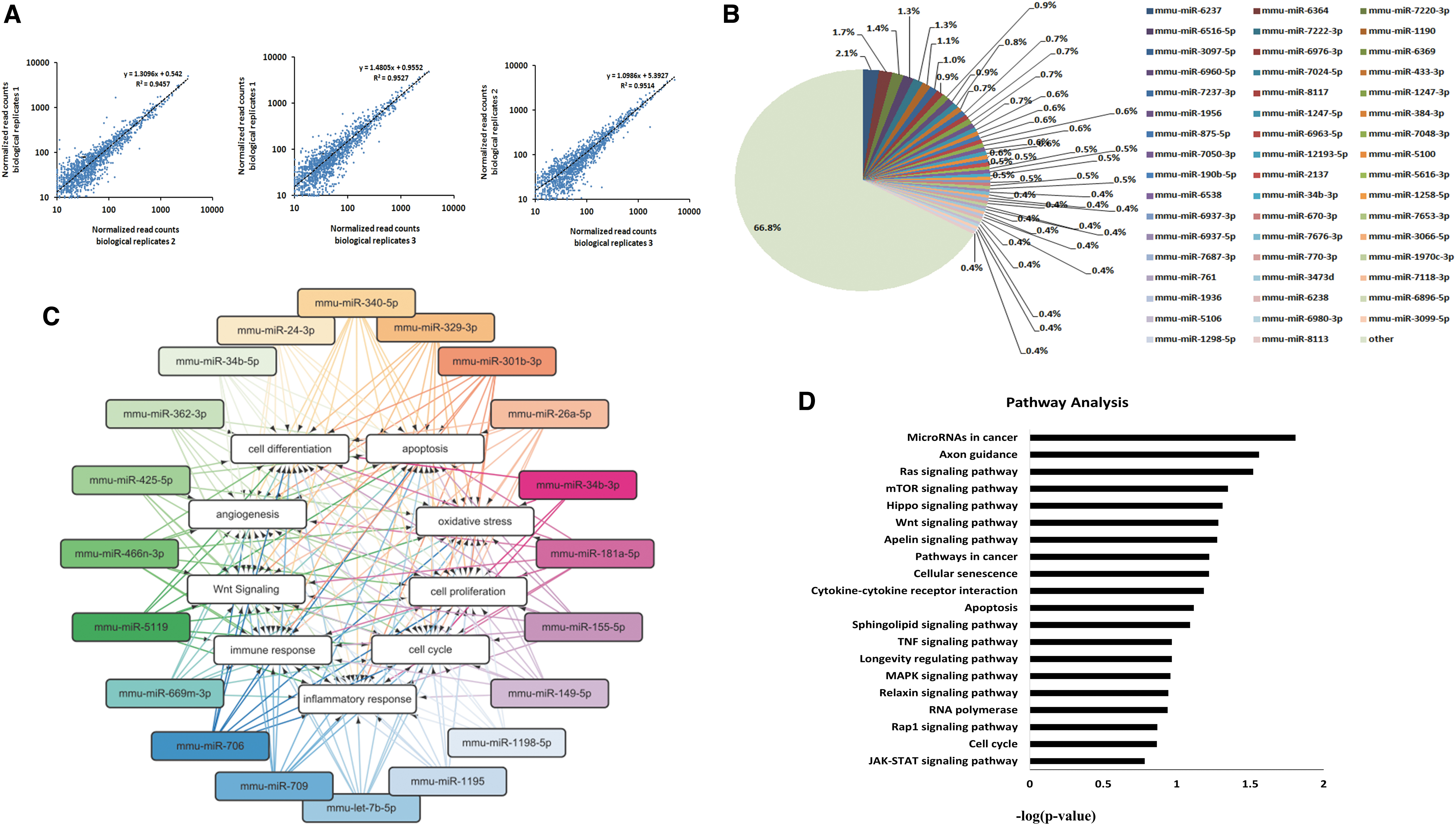
Biological processes and pathways targeted by miRNAs in BMSC-derived exosomes.
miRNAs Derived from Bone Marrow Mesenchymal Stem Cell-Derived Exosomes Involved in Important Biological Functions
BMSC-Exos, bone marrow mesenchymal stem cell-derived exosomes.
Discussion
In this study, novel results were obtained showing that PD-associated peritoneal injury in mice can be treated by i.p. injection of BMSC-Exos. BMSC-Exos also alleviated other peritoneal pathological changes, including peritoneal inflammation, PMC injury, and angiogenesis. Transcriptome results showed that the biological processes regulated by BMSC-Exos included inflammatory response, cell differentiation, cell cycle, and response to stimulus. The exosomes contained many miRNAs, which might regulate various genes that could prevent inflammation and fibrosis. Therefore, BMSC-derived exosomes might have various biological therapeutic effects on PD-associated peritoneal injury. Furthermore, i.p. injection of BMSC-Exos could be a new therapeutic strategy for treating peritoneal fibrosis.
In patients undergoing maintenance PD, repeated episodes of peritonitis represent not only a serious complication but also poor prognosis and mortality [38]. In this study, peritonitis was induced using LPS i.p. injections, which accelerated peritoneal fibrosis formation. Peritoneal inflammation and peritoneal fibrosis were observed in mice after 6 weeks. Macrophages are a primary group of resident immune cells in the peritoneum and, simultaneously, the predominant immune population found in PD effluent [39]. In vivo, MSCs relieve PD-induced peritoneal inflammation by regulating macrophage polarization [40]. In vitro, MSC-derived EVs also efficiently exerted anti-inflammatory effects through modulating macrophage phenotype switching [41]. The EVs secreted by MSCs are essentially the exosomes secreted by MSCs. Promoting the phenotypic transformation of macrophages is only one function of exosomes. We found that BMSC-Exos directly reduced the number of macrophages (F4/80+) in the peritoneum.
Moreover, MSC-treated macrophages manifested as alternatively activated macrophages possess anti-inflammatory properties [42]. KEGG analysis indicated that exosome treatment involved many inflammatory pathways and immune-related pathways. Therefore, BMSC-Exos might regulate various proinflammatory signaling pathways, inhibit the activation of immune cells such as macrophages, and alleviate peritoneal inflammation.
Myofibroblasts also emerged to be the principal effector population accounting for peritoneal fibrosis [43]. In a rat model of peritoneal fibrosis, MSCs inhibited myofibroblast accumulation in the peritoneum [9]. Similarly, BMSC-derived exosomes significantly inhibited α-SMA expression, a myofibroblast-specific marker, in the peritoneum. Myofibroblasts play crucial roles in fibrosis progression, so elucidating their origin has scientific and practical implications. Lineage tracing revealed fibroblasts as major precursor cells of myofibroblasts in peritoneal fibrosis, playing a major role in promoting peritoneal thickening and fibrosis [44]. BMSC-derived exosomes penetrated deeply into the submesothelial fibroblasts (collagen-I+ vimentin+) in the peritoneum (Fig. 1G), suggesting that exosomes might regulate some biological processes of submesothelial fibroblasts to alleviate peritoneal fibrosis. MSC-Exos could regulate cell differentiation (Figs. 4D, E, 5C and Supplementary Table S3), which might involve fibroblast-to-myofibroblast transdifferentiation. However, the specific mechanism of exosomes regulation remains unclear.
Exosomes, which deliver multiple cargoes (eg, proteins, lipids, miRNAs, and mRNAs) from donor to recipient cells, can mediate cell processes such as inflammatory response, cell differentiation, cell proliferation, and cell metabolism [45]. However, the therapeutic effect of exosomes depends on the miRNAs in exosomes. For instance, MSC-Exos-mediated miR-181-5p transfer activated autophagy and inhibited liver fibrosis [46]. MSC-Exos-derived miR-199a-3p inhibited heart fibrosis by initiating autophagy [17]. Exosomes from miR-133a-modified BMSCs ameliorated fibrosis [47]. Comprehensively exploring miRNA expression in exosomes from different MSCs is necessary to offer more strategies and potential therapeutic targets when managing fibrotic disorders. Therefore, by classifying the signaling pathways and biological roles that miRNA may affect, we found that the peritoneal protection provided by BMSC-Exos may be completed by a series of miRNAs of different functions. However, further research is needed.
Conclusions
Our study provides the first comprehensive insight into the therapeutic effects of exosomes on PD-associated peritoneal fibrosis. BMSC-Exos treatment exerts a widespread protective effect mainly on peritoneal inflammation and fibrosis through a series of miRNAs. Therefore, BMSC-Exos therapy might be a promising cell-free treatment modality for PD-associated peritoneal fibrosis. The safety and efficacy of BMSC-Exos treatment for clinical translation need to be further examined.
Data Availability
All data are available from the corresponding authors on reasonable request.
Footnotes
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This work was supported by the grants from the National Natural Science Foundation of China (82200838, 82270768) and Chongqing Technology Innovation Project (2022YSZX-JCX0007CSTB, 2023QNXM007, and SKLKF202202).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.