
Abstract
Corneal integrity, transparency, and visual acuity are maintained by corneal epithelial cells (CECs), which are continuously renewed by limbal epithelial stem cells (LESCs). The limbal stem cell deficiency is associated with ocular diseases. This study aimed to develop a novel method to differentiate bone marrow mesenchymal stem cells (BM-MSCs) into LESC-like cells using a culture medium and paired box 6 (Pax6) transfection. The LESC-like cells were confirmed using the LESC markers CK14 and p63 and CEC marker CK12. Pax6 induces BM-MSCs to differentiate into LESC-like cells in vitro. Mouse models of chemical corneal burn were obtained and treated with the LESC-like cells. The transplantation experiment indicated that Pax6-reprogrammed BM-MSCs attached to and replenished the damaged cornea through the formation of stratified corneal epithelium. The proliferation and colony formation abilities of Pax6-overexpressing BM-MSCs were significantly enhanced. These findings provide evidence that BM-MSCs might serve as an excellent candidate for generating bioengineered corneal epithelium and provide a new strategy for the treatment of clinical corneal damage.
Introduction
Corneal damage is a defect in the exposed anterior portion of the ocular epithelium, which is often the result of trauma [1 –3]. It is the most common cause of eye-related emergency department visits [4]. The main causes of corneal damage are direct mechanical trauma, contact lens-related injury, foreign bodies (corneal or intraocular), and motor vehicle accidents [1,2,4]. Corneal transplantation remains the most effective treatment for corneal damage, but there are several limitations in the source of limbal tissues [5].
Limbal epithelial stem cells (LESCs) play a very important role in corneal epithelial cell (CEC) proliferation and differentiation [6,7]. The LESCs are located at the junction of the sclera and cornea [6,7]. Severe corneal damage caused by chemical or mechanical factors and immune or hereditary diseases can lead to the destruction of the LESCs, causing limbal stem cell deficiency (LSCD), which ultimately leads to blindness [8 –10]. If corneal transplantation is used, there is a risk of developing LSCD in the donor eye, which adds controversy to the transplantation of autologous limbal epithelium [11]. The development of stem cell transplantation therapy represents a significant breakthrough in the treatment of LSCD, thus providing a more rational, less invasive, and better physiological treatment option in regenerative medicine for the ocular surface [12 –14], which indicates that we can use this technology for corneal damage treatment. Unfortunately, the number of LESCs is small and would require harvesting corneal tissues, causing trauma to the donor eye. Previous research revealed that human embryonic stem cells (ESCs) are able to differentiate into CEC-like cells [15]; however, the use of embryonic tissues will raise ethical issues.
The application of mesenchymal stem cells (MSCs) as bioengineered corneal epithelia has been attempted in several experimental studies [16 –18]. Previous studies showed that MSCs could differentiate into CECs [16 –18], but CECs are terminal cells with a limited ability to proliferate and differentiate. Therefore, it would be more desirable to differentiate MSCs into LESCs, which have more physiological functions and differentiation potentials. Hence, a way to circumvent this limitation would be to induce the differentiation of MSCs into LESCs. Bone marrow mesenchymal stem cells (BM-MSCs) possess several advantages, including easy accessibility, low immunogenicity, potent pluripotency, a lack of ethical controversies, and a reduced risk of contamination [19 –22], suggesting that BM-MSCs might be a good candidate for differentiation into LESCs. However, the growth and differentiation of cells are affected by the internal and external environment. The differentiation of BM-MSCs into LESCs continues to pose a challenge because the growth factors and three-dimensional signals that control BM-MSC differentiation have remained elusive. Most previously published studies relied on the use of undefined factors such as cell culture medium (containing insulin, hydrocortisone, triiodothyronine, fibrin substrates, collagen substrates), feeder cells, Bowman's membrane, or amniotic membrane [23 –25].
Paired box 6 (Pax6) is a member of the Pax gene transcription factor family. Pax6 is required for embryonic and postnatal development and the maintenance of the adult cornea [24 –26]. Previous reports have shown that Pax6-transfected skin epithelial stem cells (SESCs) can differentiate into LESC-like cells in vitro [23,27]. Thus, we speculated that the differentiation of BM-MSCs into LESC-like cells could also be induced by Pax6 transfection.
Therefore, this study aimed to develop a novel method to differentiate BM-MSCs into LESC-like cells using a special cell culture medium [27] and Pax6 transfection. This method has substantial potential to provide an infinite source of LESCs, with similar phenotypic and functional characterization to LESCs, for basic biological research and LSCD treatment.
Materials and Methods
Isolation, culture, and identification of mice BM-MSCs
BM-MSCs were isolated from the bone marrow of healthy BALB/c mice (6–8 weeks of age) purchased from the Experimental Animal Center of Guizhou Medical University. All animal experiments were performed in accordance with the guidelines of the Chinese Council on Animal Care. The present study was approved by the Guizhou Medical University the Animal Care Welfare Committee (permit. no. 1900021). Bone marrow was washed extensively with Dulbecco's modified eagle medium/nutrient mixture F-12 (DMEM/F12) (C11330500BT; GIBCO, Invitrogen Inc., Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS) (Biological Industries, Kibbutz Beit Haemek, Israel) followed by centrifugation at 500 g for 15 min. The supernatant was discarded. The cell pellets were resuspended in DMEM/F12 supplemented with 10% FBS and incubated at 37°C in a 5% CO2 incubator. After 24–48 h, nonadherent cells were discarded, and the medium was changed every 2–3 days. The cells were maintained in culture until they reached 80% confluence and were examined daily using an inverted fluorescence microscope. The BM-MSCs at the third passage were trypsinized, washed with phosphate buffered saline (PBS), and incubated with antibodies against CD34, CD45, CD90.2, and CD29 (BioLegend, San Diego, CA) for 30 min at 4°C. Flow cytometry was performed using a BD Celesta flow cytometer (BD Biosciences, Franklin Lake, NJ). The osteogenic and adipogenic differentiation potential of P3 generation cells was analyzed using the corresponding kits (Cyagen Bioscience, Santa Clara, CA). Matrix mineralization was confirmed by alizarin red staining after 30 days of induction. Adipogenesis was confirmed by oil red O staining after 21 days of induction.
Stable transfection with Pax6
To overexpress Pax6, BM-MSCs were infected with lentiviruses (Lv)-Pax6-green fluorescent protein (rLV-EF1α-PAX6-ACGFP-CMV-Neo-WPRE; BrainVTA, China) at a multiplicity of infection (MOI) of 50 at 37°C for 24 h and then cultured with fresh medium for 48 h at 37°C in a 5% CO2 incubator before the subsequent experiments. To silence the expression of Pax6, BM-MSCs were infected with Lv-short hairpin RNA Pax6-GFP [rLV-U6-shRNA2(PAX6)-CMV-ACGFP-T2A-Neo-WPRE; BrainVTA] at a MOI of 50 at 37°C for 24 h and then cultured with fresh medium for 48 h at 37°C in a 5% CO2 incubator before the subsequent experiments. The cells infected with Lv-ACGFP (BrainVTA; MOI, 50) were negative controls. GFP expression was analyzed using fluorescence microscopy following transfection. The three group cells were exposed to 200 mg/mL of G418 (Clontech Laboratories, Inc., Mountain View, CA). After 14 days, the stable G418-resistant clones with good growth were isolated and subcultured. The resultant clusters were observed, and images were captured under a fluorescence microscope. Subsequently, the expression of marker proteins was determined in the stably transfected cells at the third passage using immunofluorescence staining and Western blot analysis.
Western blot analysis
The total proteins were extracted from BM-MSCs using the RIPA Lysis Buffer (Beyotime Institute of Biotechnology, Haimen, China). The protein concentration was measured using a BCA Protein Assay Kit (Beyotime Institute of Biotechnology). The proteins (30 μg/lane) were separated using 10% sodium dodecyl sulfate-polyacrylamide gels and transferred to polyvinylidene fluoride membranes (Millipore Corp., Billerica, MA). The membranes were incubated with 5% bovine serum albumin at 24°C for 2 h. The primary antibodies were incubated at 4°C for 16 h. The secondary HRP-conjugated IgG antibody (1:3,500; STAR208P; Neobioscience Technology Co., Shenzhen, China) was incubated at 24°C for 2 h. The primary antibodies included glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (1:2,000, GTX100118; GeneTex, Inc., Irvine, CA) and Pax6 (1:1,000, GTX55739; GeneTex). The protein expression levels were detected using the enhanced chemiluminescence kit assay system (CLINX-5600; Clinx Science Instruments Co., Ltd., Shanghai, China). The band densities were analyzed using Image Lab 4.1 (Media Cybernetics, Inc., Rockville, MD) and standardized according to the expression of GAPDH.
Reverse transcription PCR
Total RNA was extracted from BM-MSCs using the TRNzol Universal Total RNA Extraction Kit (DP424; Tiangen Biotech Co., Ltd., Beijing, China), and complementary DNA was synthesized using the FastKing-RT SuperMix (KR118; Tiangen Biotech) according to the manufacturer's protocol. Reverse transcription PCR (RT-PCR) was performed using the 2 × SanTaq PCR Master Mix (with Blue Dye) (B532061; Sangon Biotech [Shanghai] Co., Ltd., Shanghai, China) on an ABI thermocycler (Applied Biosystems, Foster City, CA). The PCR conditions were (1) 30 s at 95°C and (2) 35 cycles of 5 s at 95°C, 30 s at 60°C, and 7 min at 72°C. GAPDH was used as the internal control. The following primer sequences were used: Mouse GAPDH Endogenous Reference Gene Primers (B662304; Sangon Biotech [Shanghai] Co., Ltd.), Pax6 (mouse) S-5′-ATC AGA GTC GAC ATG CAG AAC AGT CAC AGC GGA G-3′ and A-5′-ATC TGA GGA TCC TTA CTG TAA TCG AGG CCA GTA CTG-3′.
Colony formation assay
The cells from each group were seeded onto 6-well plates at 50 cells/well in triplicates. Fresh basic growth medium was added to each well following incubation for 10 days under 5% CO2 at 37°C. The culture medium was aspirated; each well was washed with PBS and added with 1 mL 0.5% crystal violet solution (C8470; Solarbio Science & Technology Co., Ltd., Beijing, China), followed by incubation for 10 min at room temperature. Each well was washed with a gentle stream of water under running tap water. The number of “darkly” stained colonies was counted under a phase-contrast microscope at low magnification. Values were expressed as colony-forming efficiency calculated as the percentage of the ratio of the number of colonies counted versus the number of cells initially seeded.
Cell proliferation analysis
Cell number counting was performed to assess cell proliferation, as previously described. The cells of the GFP/BM-MSCs, Pax6/BM-MSCs, and shPax6/BM-MSCs groups were released from the culture and dispersed by gently pipetting up and down several times. The cells were detached from the culture dishes using 0.05% trypsin, and phycoerythrin anti-mouse Ki67 antibody (1:100; 652404; BioLegend) was added to the cell suspension. The samples were analyzed on fluorescence-activated cell sorting (FACS).
Cell apoptosis
The cells from each group were digested with trypsin and centrifuged. The supernatant was discarded, and the cells of each group were collected. The cells were rinsed with precooled PBS twice. The resuspended cells were gently resuspended with an appropriate amount of buffer. The cells were adjusted to 1 × 109/L. An Annexin V-FITC Kit (556547; BD Biosciences) was used. The cell suspension (100 μL) was mixed with 5 μL of Annexin V-FITC, incubated at room temperature hidden from light for 15 min, and 5 μL of propidium iodide staining was added.
Induction of mouse BM-MSCs into LESCs
The cells of the stable G418-resistant clones in the GFP/BM-MSCs, Pax6/BM-MSCs, and shPax6/BM-MSC groups with good growth morphology were differentiated using 3 × 105 cells per plate (six wells). The cell culture medium was composed of the following components: DMEM/F12, 1/100 penicillin–streptomycin (15140122; GIBCO), 20% fetal bovine serum (10099141C; GIBCO), 5 μg/mL insulin (I6634; Sigma, St. Louis, MO), 0.4 μg/mL hydrocortisone (G8450; Solarbio Science & Technology Co., Ltd.), 10–10 M cholera toxin (C8052; Sigma), 2 × 10−10 M 3,3’,5-triiodo-L-thyronine (T2877; Sigma), 50 ng/mL epidermal growth factor (315-09; PeproTech, Rocky Hill, NJ), 10 ng/mL transforming growth factor (100-21; PeproTech), and 5 ng/mL nerve growth factor (11050-HNAC; Sino Biological Inc., Beijing, China). The medium was replaced every 2 days. LESC-like cells were formed after 10 days in the cell culture medium, followed by immunofluorescence and flow cytometry to detect the expression of the LESC markers CK14 and p63 and the CEC marker CK12.
Immunofluorescence
The cells were fixed with 4% paraformaldehyde (BL539A; Biosharp, Shanghai, China) for 45 min and incubated in 0.2% Triton X-100 (T8200; Solarbio Science & Technology Co., Ltd.) for 20 min and 5% goat serum (ZLI-9022; ZSGB-BIO, Beijing, China) for 40 min. The primary antibodies included p63 (1:200, GTX102425; GeneTex, Inc.), anti-cytokeratin 14 (1:200; ab7800; Abcam, Cambridge, MA), and anti-cytokeratin12 (1:100; ab185627; Abcam). Cells were incubated with the primary antibodies overnight at 4°C. The secondary antibodies, Cy3-labeled Goat Anti-rabbit (H+L) (A0516; Beyotime Institute of Biotechnology) or Cy3-labeled Goat Anti-mouse (H+L) (A0521; Beyotime Institute of Biotechnology), were used at a dilution of 1:300 for 2 h. 4′,6-diamidino-2-phenylindole (1:50; BB4401; BestBio, Shanghai, China) was used for 15 min. Images were obtained using laser scanning confocal microscopy (model 1000; Olympus Corporation, Tokyo, Japan).
Flow cytometry analysis
The single-cell suspensions were stained with fluorochrome-conjugated antibodies, as described. For intracellular staining, the cells were permeabilized with a BD Cytofix/Cytoperm solution for 30 min at 4°C. The primary antibodies were the same as before. The secondary antibodies, donkey anti-rabbit IgG H&L (AlexaFluor-647) (1:200; Ab150075; Abcam, Cambridge, United Kingdom) or goat anti-mouse IgG H&L (Phycoerythrin) (1:200; Ab97024; Abcam), were used at 1:200 for 30 min. The samples were analyzed on a FACS.
Mouse model and grouping
Female BALB/c mice (6 weeks of age) were used for transplantation recipients. The right eye of each mouse was injured, and the left eyes were used as a control without treatment. Modeling was performed as previously described [28]. Lidocaine was used for topical anesthesia of the right eye. Atropine was used to dilate the pupil for 1 min. A 2-cm diameter round paper soaked in 1 mol/L sodium hydroxide was placed on the right cornea for 30 s, followed by rinsing with 0.9% saline for 1 min. Local treatment with 0.3% levofloxacin drops and tobramycin thrice a day for 2 weeks was given to prevent secondary infection.
Corneal haziness was graded based on the criteria used by Jiang et al. [29]. Corneal opacity, neovascular conditions, and epithelial defects were observed. The corneal score was scored according to these conditions 1 week after modeling. After alkali injury, the mice with corneal opacity grade ≥2 and cornea neovascularization grade ≥3 were included in the study.
Thirty-six mice with successful modeling were randomly divided into three groups: GFP/bone marrow mesenchymal stem cell (BMMSC)-LESCs (Group 1, n = 12), Pax6/BMMSC-LESCs group (Group 2, n = 12), and shPax6/BMMSC-LESCs group (Group 3, n = 12). After modeling, the injured eye immediately received a single subconjunctival injection of stem cell suspension (1 × 105 cells/0.1 mL), while the uninjured eyes received 0.1 mL of normal saline in a similar manner. The mice were euthanized on day 42 by CO2 asphyxiation, and the corneas were removed.
Histological analysis
The isolated corneas in paraffin sections were prepared after fixation, dehydration, and implantation with thickness of 6 μm and routine hematoxylin-eosin (HE) staining. The morphological changes of corneas were observed under the microscope. Subsequently, the evaluation of corneal opacity, neovascularization grades was determined by a graded scale. For immunofluorescence analyses, deparaffinized sections were treated with a primary antibody anticytokeratin 3 (1:50; Ab68260; Abcam) at 4°C overnight and subsequently incubated with secondary antibodies conjugated with cy3 in the dark for 1 h at room temperature. The secondary antibodies were the same as those used for immunocytochemistry. Fluorescence was observed under a fluorescence microscope.
Cytokine measurement in serum
Blood samples were collected and centrifuged at 1200 g for 10 min. The serum levels of cytokines of vascular endothelial growth factor (VEGF) were measured by enzyme linked immunosorbent assay (ELISA) Kits (EMC103.96; NeoBioscience) according to the manufacturer's instructions.
Statistical analysis
Data were analyzed using GraphPad Prism v5.01 (GraphPad Software Inc., San Diego, CA) and presented as the mean ± standard error of the mean of ≥3 independent experiments. The differences between two groups were analyzed using the unpaired Student's t-test. The differences among multiple groups were determined by one-way analysis of variance with Tukey's post hoc test. P < 0.05 was considered statistically significant.
Results
Characterization of mouse BM-MSCs
A small number of scattered adherent cells were observed after 24 h of culture of the bone marrow cells. Most were spindle shaped and circular. The cells grew rapidly from days 5 to 10. Many colonies were gradually formed and then aggregated into bulks. At this moment, the cells were triangular and fusiform in shape, and a large number of round cells were scattered on days 10 to 14. The cell density reached 80%–90%. After passaging, the cells were floating evenly in the medium instead of colony-like growth. At the third generation, the cells were fusiform in shape, obviously different from the primary cells (Fig. 1A). Many orange-red calcium nodules were found after 30 days of cell culture after osteogenic differentiation (Fig. 1B-1). At 21 days after induction of adipogenic differentiation, oil red O staining showed red lipid droplets (Fig. 1B-2). The results showed that the cultured cells had the potential to differentiate into osteoblasts and adipocytes. Flow cytometry showed that the third-generation BM-MSCs expressed the MSC surface markers CD29 and CD90.2, but not the hematopoietic stem cell surface markers CD34 and CD45 (Fig. 1C).
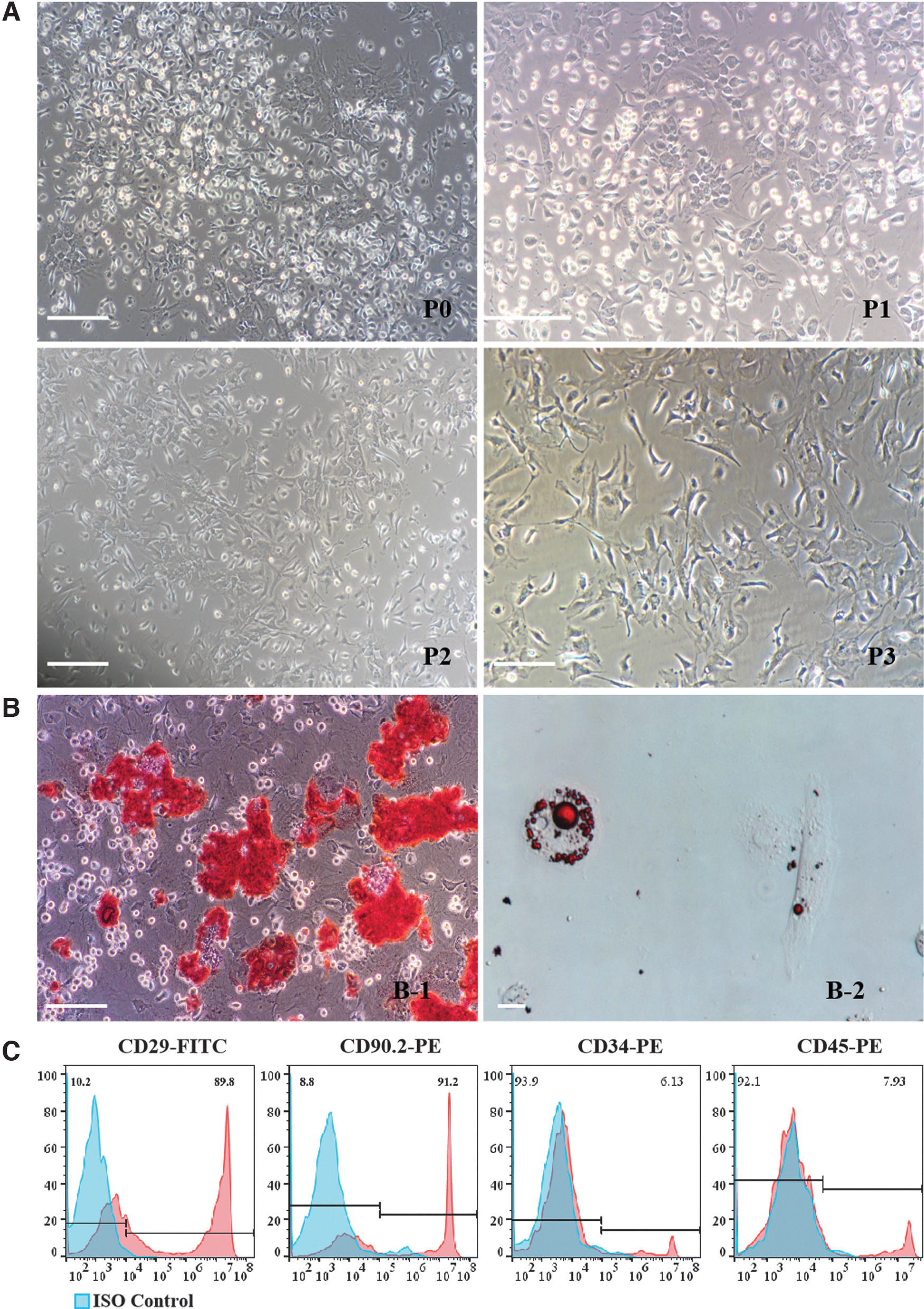
Characteristics of murine BM-MSCs. BM-MSCs were isolated from BALB/c mice, and the cells were collected from the femur and tibia.
Stable transfection with Pax6
Successful transfection of Pax6 and GFP was observed in BM-MSCs, as evidenced by green fluorescence from the BM-MSCs transfected with the GFP control or Pax6 expression plasmid (Fig. 2A). RT-PCR and Western blot confirmed that Pax6 messenger RNA and protein were expressed in the Pax6/BM-MSCs, but not in the shPax6/BM-MSCs and GFP/BM-MSCs (Fig. 2B, C/Supplementary Fig. S1). No morphological differences were observed among the three groups of cells.
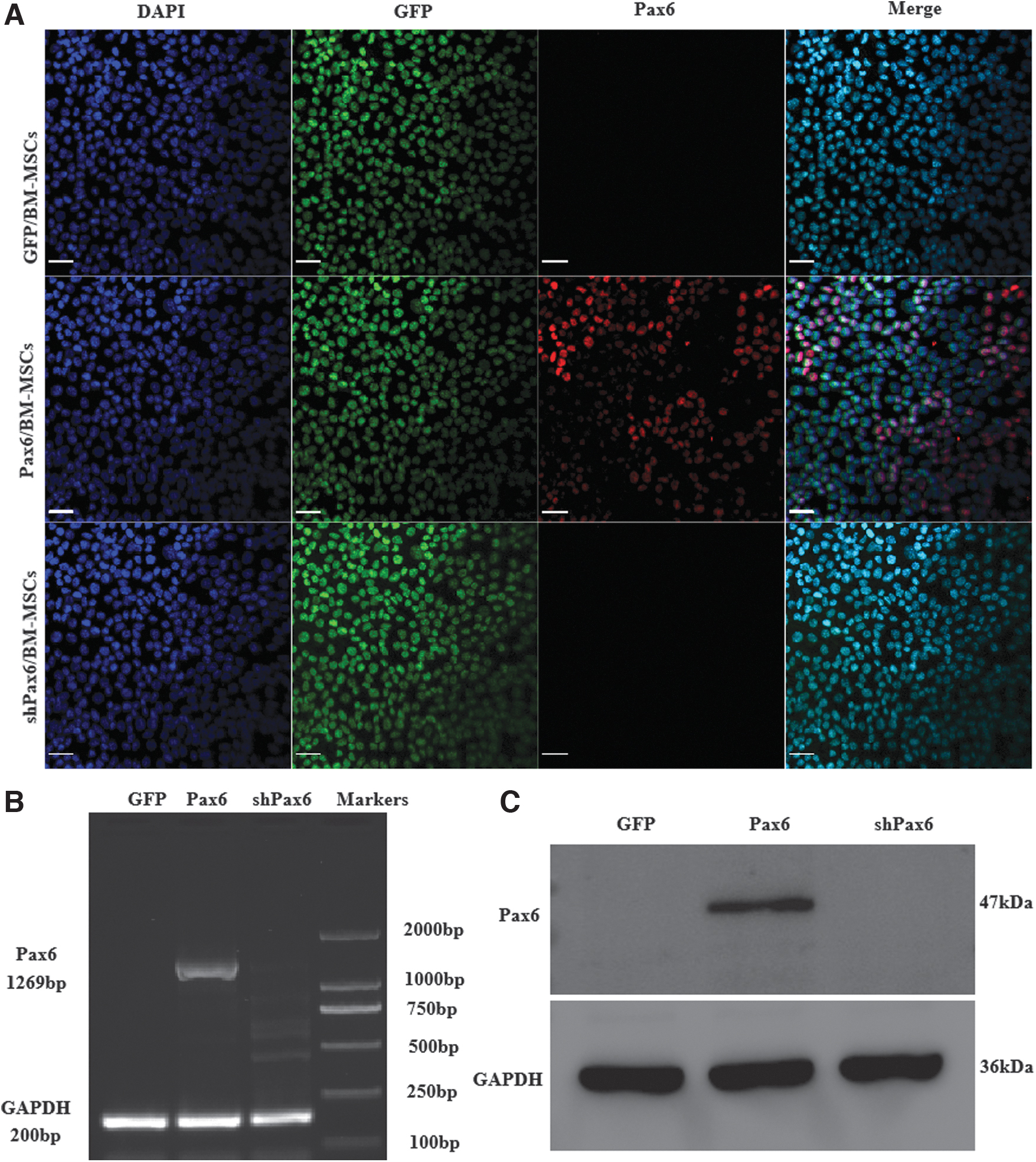
Stable Pax6 transfection into mouse BM-MSCs.
Effect of Pax6 expression on the proliferation and apoptosis of mouse BM-MSCs
The colony formation in the Pax6/BM-MSCs group was significantly higher than in the GFP/BM-MSCs group on the 10th day of culture, while the colony formation in the shPax6/BM-MSCs group was lower than in the GFP/BM-MSCs group (Fig. 3A). Compared with the GFP/BM-MSCs group, the expression of Ki67 in the Pax6/BM-MSCs group was higher (P < 0.05), while Ki67 expression in the shPax6/BM-MSCs group was lower than in the GFP/BM-MSCs group (P < 0.05) (Fig. 3B/Supplementary Fig. S2). Compared with the GFP/BM-MSCs group, the apoptosis rate was significantly decreased in the Pax6/BM-MSCs group (P < 0.05). In the shPax6/BM-MSCs group, the apoptosis rate was significantly increased (P < 0.05) (Fig. 3C).

Effect of Pax6 expression on the proliferation capacity and apoptosis of mouse BM-MSCs.
Differentiation of BM-MSCs into LESCs
In the first 5 days of differentiation, the cells proliferated, most of the cells became round and oval, and the spindle cells decreased. On day 10, the number of cells increased, almost all the cells were round and oval, and a small number of spindle cells remained (Fig. 4A). The morphological changes of the cells in the GFP/BM-MSCs and Pax6/BM-MSCs groups were similar, but the cells in shPax6/BM-MSCs group grew more slowly than those in the GFP/BM-MSCs and Pax6/BM-MSCs groups, and some epithelial cell-like islands appeared, showing typical pebble-like features (Fig. 4A).
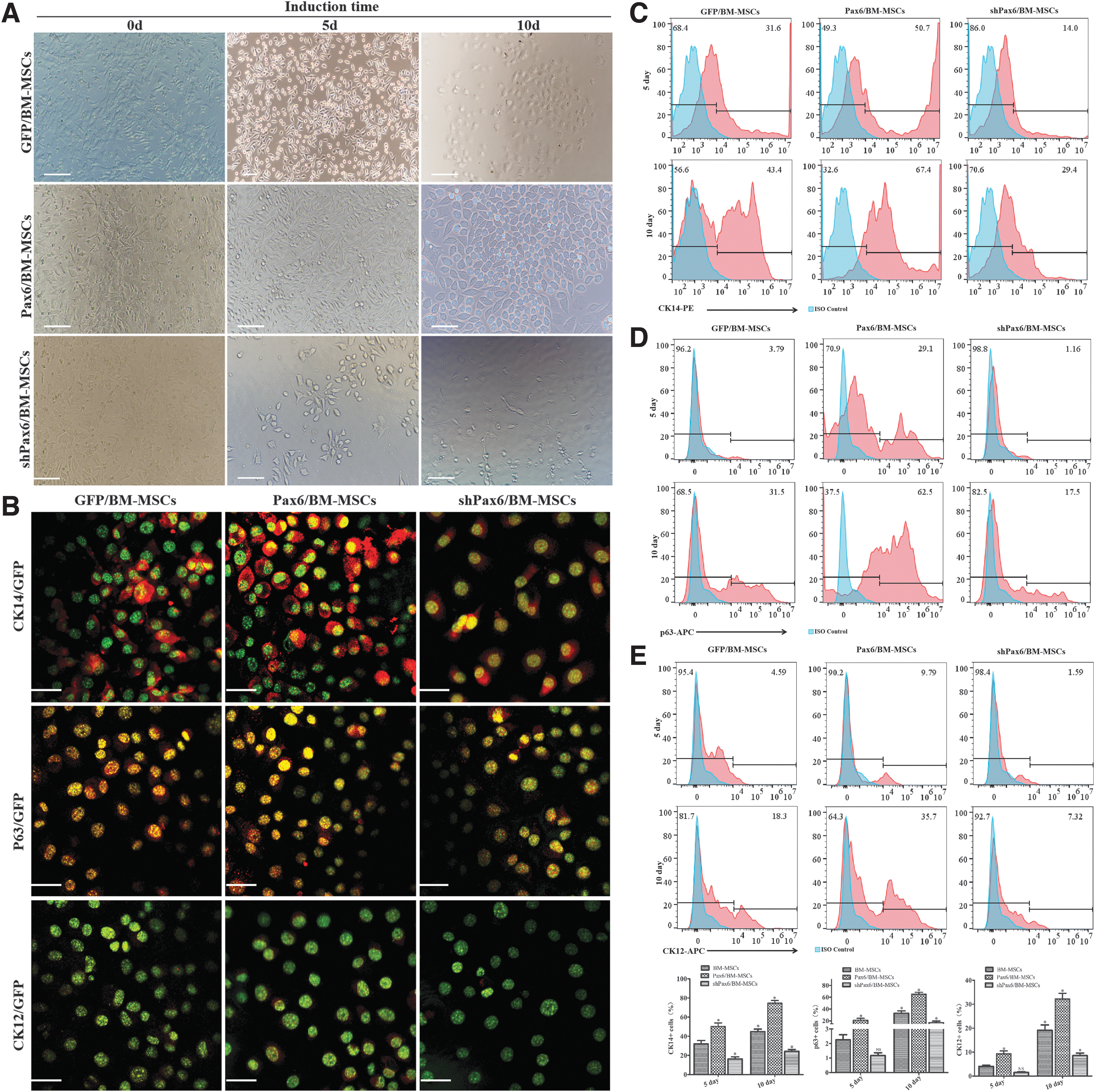
Morphology of cells from BM-MSCs at different stages of differentiation.
The immunofluorescence of the cells showed that CK14 and p63, which are markers for LESCs [30,31], were strongly expressed in the nucleus and cytoplasm of the induced cells in the Pax6/BM-MSCs and GFP/BM-MSCs groups. CK12 is a marker of terminally differentiated CECs [32] and was identified at the periphery of some colonies on day 10 of differentiation in the Pax6/BM-MSCs group (Fig. 4B/Supplementary Fig. S3). The immunofluorescence of p63 could not be detected in the cells in the shPax6/BM-MSCs group, and the expression of CK14 was also very weak. The data showed that the number of p63-positive cells and CK14-positive cells in the Pax6/BM-MSCs group was significantly higher than in the other groups, with an average of 77.9% of cells expressing p63 and an average of 76.8% of cells expressing CK14 at day 10 of differentiation (Fig. 4C, E). There were significant differences between the Pax6/BMMSC-LESCs group and the other groups in CK14, p63, and CK12 expression (P < 0.05). These data indicate that BM-MSCs cultured using a cell culture medium can differentiate into LESCs. The level of expression of Pax6 is more conducive to the differentiation of LESCs from BM-MSCs. The expression of CK12 demonstrated that this culture condition might be helpful in the maintenance of stem cell properties, and the cells at the periphery of some colonies can differentiate into CECs.
Transplantation of LESC-like cells to the damaged mouse cornea
The corneal tissue structure of the left eyes of the mice was intact. The corneal epithelium was nonkeratinized stratified squamous epithelium with 5–6 layers. Corneal stromal collagen fibers were arranged in parallel, with a small number of spindle corneal cells, without inflammatory cells and neovascularization. The corneal epithelium of the right eyes (alkali burned) was damaged. The corneal stroma structure was disordered, with numerous inflammatory cells and neovascularization (Fig. 5A).
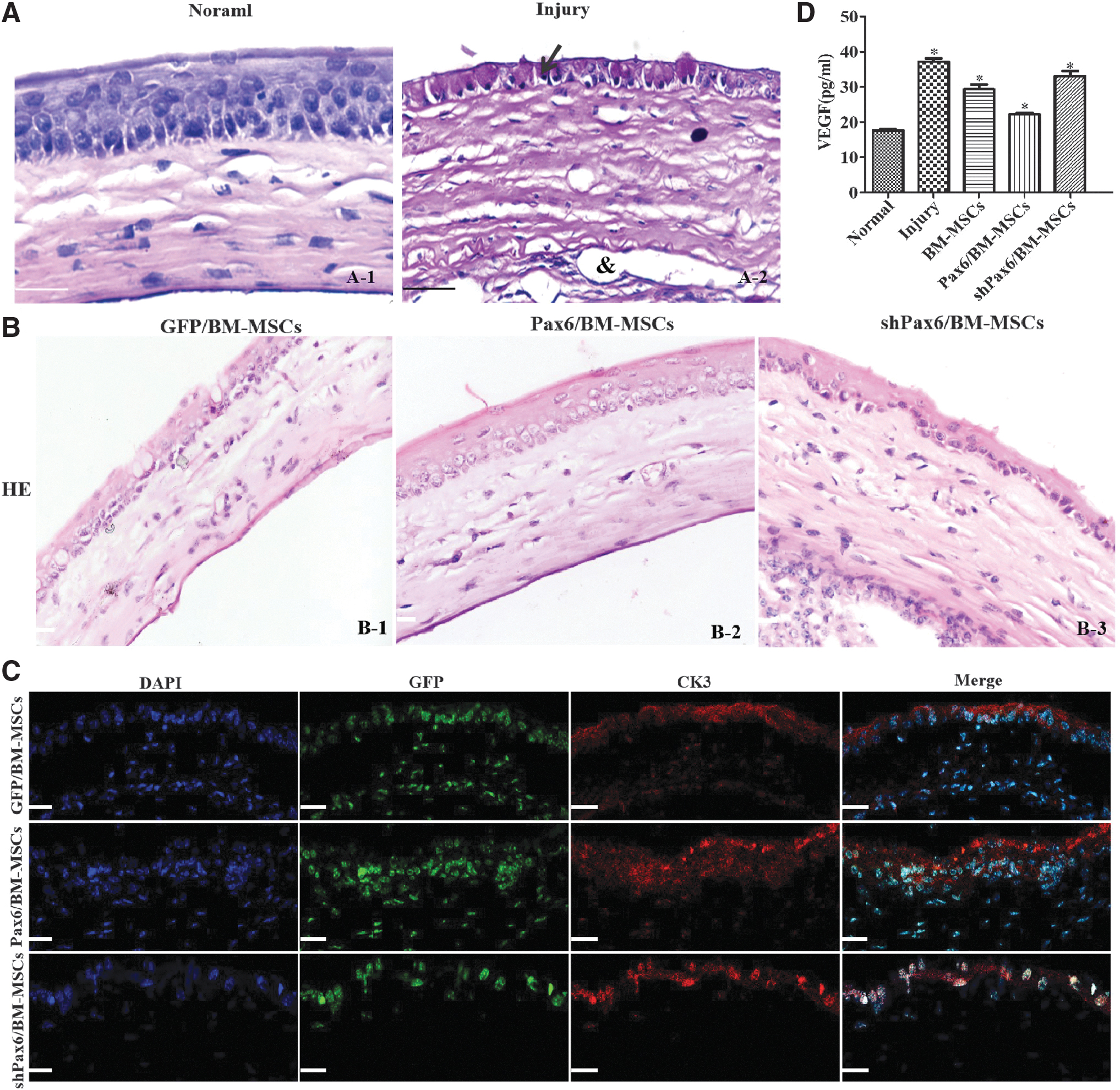
Cell engraftment and corneal epithelium repair in the mice model of alkali burn.
In the Pax6/BMMSC-LESCs group, HE staining revealed that the injured cornea demonstrated an intact morphology of the corneal surface, with stratified epithelial cells, a few lymphocytes, and neovascularization infiltration of the corneal stroma (Fig. 5B-2). In the GFP/BMMSC-LESCs group, the epithelium was integrated, and some inflammatory cells infiltrated the corneal stroma (Fig. 5B-1). Incomplete epithelial hyperkeratosis and inflammatory cell infiltration were observed. In the shPax6/BMMSC-LESCs group, the cornea exhibited epithelial and stromal defects to various degrees and complete epithelial hyperkeratosis, and the stromal layer had many inflammatory cells' infiltration (Fig. 5B-3). The confocal image of the damaged cornea that underwent transplantation of LESC-like cells revealed attachment of GFP-positive cells to the impaired corneal surface, with concomitant expression of the CEC-specific marker CK3. These results indicated that the grafted Pax6-overexpressing cells possessing the characteristics of mature CECs grew onto and reconstructed the damaged corneal surface. In the two other groups, there were few CK13-positive cells in the corneal epithelium (Fig. 5C/Supplementary Fig. S4). There were significant differences between the Pax6/BMMSC-LESCs group and the other groups regarding the corneal epithelial, corneal opacity, and neovascularization grades at 42 days after transplantation (all P < 0.05, Table 1).
Corneal Scores After Different Treatment for Corneal Alkali Burn Injury in Mice
p < 0.05, vs Injury group.
BM-MSCs, bone marrow mesenchymal stem cells; GFP, green fluorescent protein; Pax6, paired box 6; shPax6, short hairpin RNAs Pax6.
The expressions of VEGF in the serum were examined by ELISA. As shown in Fig. 5D, the VEGF expression was significantly increased after damaged mouse cornea; the LESC-like cell transplantation caused a decreased expression of VEGF in alkali mice. There were significant differences between the Pax6/BMMSC-LESCs group and the other groups (P < 0.05).
Discussion
This study developed a novel method to differentiate BM-MSCs into LESC-like cells using a cell culture medium and Pax6 transfection. The results suggest that the conversion of BM-MSCs into LESC-like cells might be driven by Pax6 transfection. Therefore, BM-MSCs might be a promising cell candidate for the treatment of LSCD.
Corneal damage can be caused by trauma, contact lens-related injury, foreign bodies, and chemical/radiation burns [1,2,4]. The integrity of the corneal surface is maintained by the CECs, and the CECs are continuously renewed by the LESCs. LSCD will lead to failure of epithelial regeneration or nontransparent cornea, causing corneal surface disease and blindness [23,29]. Although allogeneic and autologous limbal transplantations are useful treatment modalities for these pathological conditions, their clinical application is currently limited due to potential complications such as corneal graft immunosuppression and impairment of the contralateral healthy eye [5,10]. Our study showed that BM-MSCs could be differentiated into LESC-like cells. BM-MSCs possess multiple advantages, and their main advantages include easy accessibility, abundant supply in each individual or animal, potent pluripotency, and no ethical issues [19 –22]. Thus, mice BM-MSCs were used as seed cells in the present study to explore whether the Pax6 gene induces the transdifferentiation of BM-MSCs into LESC-cells like that could be used to manage corneal damage.
Previous studies tried to use MSCs to induce differentiation into LESCs [16 –18], but the results were disappointing. BM-MSCs can differentiate into LESCs, but the process is challenging because the proposed methods rely on precise environmental and growth conditions [23 –27]. It has been revealed that Pax6-transfected SESCs can differentiate into LESC-like cells in vitro [27]. Therefore, Pax6 might be a key to improving the differentiation efficiency of BM-MSCs into LESCs. Pax6 is a homeobox transcription factor acknowledged to have a critical role in CECs' fate determination and differentiation of LESCs and CECs. Previously, Ueno et al. [1] revealed that the overexpression of Pax6 promoted the differentiation of mouse ESCs into CEC-like cells in vitro. Upon transplantation onto eyes in a rabbit corneal injury model, these Pax6-reprogrammed ESCs could replenish the CECs and repair the corneal epithelial defects. A study by Ouyang et al. [27] confirmed that Pax6 is the main driver of the differentiation of CECs and that rabbit SESCs with Pax6 expression could transdifferentiate into CEC-like cells. These findings offer new hope for treating corneal injuries, but there are still some hurdles for their clinical application. Although SESCs have priorities over ESCs due to easier accessibility and avoiding ethical controversy, they continue to pose the potential risk of damaging the donor's appearance. In recent years, MSCs derived from bone marrow have been paid more attention due to the absence of the disadvantages of using SESCs and ESCs.
In the present study, we established an effective and feasible strategy for the differentiation of BM-MSCs from LESCs in vitro under defined conditions and after Pax6 overexpression. The expression of p63, CK14, and CK12 remained stable for 10 of differentiation. The expression of LESC markers, particularly CK14, was higher in the differentiated cells in the Pax6/BM-MSCs group than that in the shPax6/BM-MSCs group, indicating that elevated Pax6 was conductive to the differentiation of LESCs from BM-MSCs. Furthermore, Ki67 and colony-forming assays demonstrated that the viability and proliferative capacity of the BM-MSCs were promoted by Pax6 transfection, supported by Ueno et al. Pax6 overexpression reduces apoptosis, improving cell survival [1,33].
When the Pax6-transfected cells were transplanted onto the damaged cornea, the cornea demonstrated an intact morphology of the corneal surface with stratified epithelial cells at 42 days after transplantation. In addition, the grafted Pax6+ cells possessed the characteristics of mature CECs and grew onto and reconstructed the damaged corneal surface. VEGF is a kind of vascular endothelial cell that specifically promotes division and plays a central role in the formation of new blood vessels [34,35]. VEGF not only participates in the regulation of corneal neovascularization after alkali burn but also plays an important role in promoting the induction and maintenance of new blood vessel growth [36]. Previous studies have found that tissue inflammation and ischemia and hypoxia-induced neovascularization are the causes of corneal scarring and symblepharon adhesions [37 –40]. LESC-like cell transplantation reduced VEGF expression and reduced inflammatory cell infiltration in corneal tissue. Therefore, BM-MSC transplantation in the treatment of corneal chemical burns is mainly due to the functions of repairing tissue, anti-inflammatory expression, and antiangiogenesis (reducing the expression of VEGF).
Still, only mice were used in this study, and there might be some differences among species. These results should be confirmed in other animal models and clinical experiments. In addition, the exact molecular involved in differentiation was not analyzed. Future studies should examine the genes that are induced or inhibited by Pax6, which could help improve the process or identify potential safety issues. Furthermore, the grafting experiment was stopped at 42 days, and long-term data should be needed to determine the long-term patency and safety of the corneal tissue grown using the LESC-like cells. These issues should be examined in future studies.
Conclusion
Pax6 induces BM-MSCs to differentiate into LESC-like cells in vitro. The Pax6-reprogrammed BM-MSCs were able to form stratified epithelium in vivo. These findings provide evidence that BM-MSCs might serve as an excellent candidate for generating bioengineered corneal epithelium and provide a new strategy for the treatment of clinical corneal damage. We will provide further clarification of the BM-MSC niche and the specific mechanisms underlying the preferential differentiation of BM-MSCs into LESC-like cells in the near future.
Footnotes
Author Disclosure Statement
None of the authors has a conflict of interest.
Funding Information
This work was supported by grants from Guizhou Science and Technology Fund Project [Qiankehejichu(2020)1Y408, Qiankehejichu-ZK(2022)yiban344, to J.G.), the National Natural Science Foundation of China [NSFC 82060151, to M.S.], Science and Technology Support Program of Guizhou Province [Qiankehezhicheng (2020)4Y230, Qiankehepingtairencai (2020)4130, to M.S.].
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.