
Abstract
The effects of smoking on fetal development and stem cell differentiation are not fully understood. Although nicotinic acetylcholine receptors (nAChRs) are expressed in many organs of the human body, their significance in human induced pluripotent stem cells (hiPSCs) remains unclear. After expression levels of nAChR subunits in hiPSCs were determined, the effects of the nAChR agonist, nicotine, on undifferentiated hiPSCs were evaluated using a Clariom S Array. We also determined the effect of nicotine alone and with a nAChR subunit antagonist on hiPSCs. nAChR α4, α7, and β4 subunits were strongly expressed in hiPSCs. cDNA microarray, gene ontology, and enrichment analyses showed that exposing hiPSCs to nicotine altered expression of genes associated with immune responses, neurological system, carcinogenesis, cell differentiation, and cell proliferation. Particularly affected was metallothionein, which acts to decrease reactive oxygen species (ROS). The nicotine-induced reduction of ROS in hiPSCs was canceled by an α4 subunit or nonselective nAChR antagonist. HiPSC proliferation was increased by nicotine, and this effect, too, was canceled by an α4 antagonist. In conclusion, nicotine reduces ROS and enhances cell proliferation through the α4 nAChR subunit in hiPSCs. These findings provide new insight into the significance of nAChRs on human stem cells and fertilized human ova.
Introduction
Nicotinic acetylcholine receptors (nAChRs) are expressed throughout the human body and are divided into two types: neuronal and muscle. Muscle-type nAChRs are composed of four subunits (α, β, δ, ɛ), whereas neuronal type are composed alpha (α2–7, α9–10) and beta (β2–4) subunits. Moreover, neuronal-type nAChRs are divided into two subtypes, α4β2 heteromeric and α7 homomeric. The α7 subunit is highly expressed in the hippocampus and hypothalamus, where it plays critical roles in cognitive function and neurotransmission. Deficits in α7 function are known to be linked to mental illnesses such as schizophrenia [1] as well as to the cognitive dysfunction seen in Alzheimer's disease [2]. For that reason, α7 function is being investigated as a potential therapeutic target for these conditions. In addition, deletions and duplications of the α7 subunit of nAChRs highly expressed in the hippocampus are known to cause neurodevelopmental and neuropsychiatric disorders including intellectual disability [3].
Little is known about the expression or function of nAChRs in human stem cells. However, their presence in stem cells may have implications on the effects of smoking on fetal development and aging [4]. For example, nicotine is reported to significantly increase oxidative stress in human neuronal stem cells (NSCs) [5], whereas it reportedly prevents apoptosis induced by amyloid beta via the Wnt/β-catenin pathway [6]. In mouse NSCs, long-term exposure to nicotine decreased cell proliferation by increasing levels of nitric oxide and proinflammatory cytokines [7]. In addition, treating rat mesenchymal stem cells (MSCs) with nicotine significantly enhanced the survival rate and phagocytic activity of cocultured neutrophils and reduced the production of reactive oxygen species (ROS) induced by N-formylmethionine-leucyl-phenylalanine [8].
On the contrary, in human umbilical cord–derived MSCs, nicotine dose dependently enhanced apoptosis and interleukin-6 production and inhibited cell division [9,10]. In human alveolar bone marrow-derived MSCs, exposure to low concentrations of nicotine promoted cell proliferation, whereas exposure to high concentrations significantly inhibited it [11]. Similarly, low concentrations of nicotine promoted proliferation and osteogenic differentiation of human bone MSCs, whereas high concentrations had the opposite effect [12]. Nicotine reportedly decreases survival and adhesion by human embryonic stem cells (hESCs) [13], but it appears to increase proliferation of mouse induced pluripotential stem cells by stimulating CaMKII signaling via stimulation of the α4 and α7 nAChR subunits [14].
The effects of nicotine are thus concentration dependent and may vary between differentiated and undifferentiated cells and among different cell types. In this study, therefore, we investigated the effects of nAchR activation on gene expression in human induced pluripotent stem cells (hiPSCs).
Materials and Methods
Cell culture
Experiments using hiPSCs were approved by the Ethics Committee of Kyoto University. hiPSCs (409B2, 253G1, TIG107) and hESCs (KhES1) were cultured on iMatrix-511 silk (Nippi, Tokyo, Japan)–coated, 24-well culture plates with StemFit® AK02N (Ajinomoto, Tokyo, Japan) without feeder cells. Cells were passaged every 7 days using phosphate-buffered saline containing 0.5 mmol/L ethylenediaminetetraacetic acid (Nacalai Tesque, Kyoto, Japan).
Bioinformatics analysis
Confirmation of nAChR expression
RNA purification was performed using NucleoSpin® RNA (Takara Bio, Shiga, Japan) from hESCs, hiPSCs, MSCs, and human brain specimens purchased from the Lonza Group. Complementary DNA synthesis was performed using a GeneChip WT PLUS Reagent Kit (Thermo Fisher Scientific). The cDNA samples were subjected to PCR amplification using a thermal cycler. PCR was performed using Takara Taq™ (Takara Bio) according to the manufacturer's instructions. Ten microliters of the resulting PCR products were electrophoresed on a 1.5% agarose gel and then visualized by ethidium bromide staining.
The RNA expression levels of the α4, α7, and β4 nAChR subunits were confirmed with quantitative real-time PCR (qRT-PCR) using the primers listed in Supplementary Table S1. Relative expression levels were determined after assigning a value of 1 to the expression levels in untreated control hiPSCs. We used Step One Plus (Applied Biosystems) for analysis and 18s RNA as an internal control.
Genetic analysis by nicotine administration
The hiPSCs were seeded onto 24-well plates and incubated for 8, 24, or 72 h with medium containing 1% dimethyl sulfoxide (DMSO) (control) or 10 μmol/L nicotine [N3876 SIGMA (−) Nicotine; Sigma-Aldrich] in DMSO and then collected for microarray analysis. With cells incubated for 72 h, the culture medium was replaced every 24 h. Cells were collected with 99% 2-mercaptoethanol (Sigma-Aldrich) and diluted 1:100 in lysis buffer. From each of the six cell groups, RNA samples were isolated, cDNAs were synthesized, and hybridization arrays were performed. We used the Clariom™ S Assay, human (Thermo Fisher Scientific) for array analysis. The arrays were heated in a GeneChip Hybridization Oven 645 (Thermo Fisher Scientific) at 45°C while rotating at 60 rpm for 16 h. They were then stained and washed using a GeneChip Fluidics Station 450 (Thermo Fisher Scientific), after which images were acquired using a dedicated array scanner (GeneChip Scanner 3000 7G; Thermo Fisher Scientific). Signal intensities in the images were quantified using Affymetrix GeneChip Command Console Software 4.0. These data were then normalized and corrected for data bias using the same software. Fold changes in the signals were calculated using Transcriptome Analysis Console (Thermo Fisher Scientific) software. Gene ontology (GO) analysis was performed using the Database for Annotation, Visualization, and Integrated Discovery (DAVID) Bioinformatics Resources 6.7. We determined which genes varied significantly among the six conditions and calculated enrichment scores (ESs) to determine which genes were significantly expressed.
Quantitative RT-PCR
The protocol used for qRT-PCR was described previously [15]. Primers used in this study are listed in Supplementary Table S1. mRNA expression levels were normalized to those of 18s RNA.
Cell counting
hiPSCs were seeded onto 96-well plates and incubated for 3 days. The cells were then incubated for an additional 3 days in medium containing 1% DMSO (control), 10 μmol/L nicotine in DMSO, or 10 μmol/L nicotine plus α4 nAChR subunit. On day 6 of incubation, cell numbers were counted in MTT assays.
Measurement of intracellular ROS levels
Cells were stained using an ROS Detection Cell-Based Assay Kit (Cayman Chemical Co.) according to the manufacturer's instructions and analyzed with a multi-detection microplate reader (DS Pharma Biomedical). Cells were plated at 1,500 cells/well in Falcon Microtest Plates with Stem Fit AK02N. After 3 or 5 days, medium with or without nicotine was added to the wells. ROS levels were then measured on day 6 as dihydroethidium (DHE) fluorescence.
Statistical analysis
All data are expressed as mean ± standard error of the mean. For RNA sequencing, a fold change in expression was defined as significant when the signal intensity in the nicotine condition divided by that in the control condition was >1.5 or <2/3. The false discovery rate was defined as 0.05. In enrichment analysis, a significant increase in ES was defined as being >1.3. In comparison in the control, nicotine, nicotine+antagonist, and N-acetylcysteine (NAC) group, statistical analysis was performed using a two-tailed unpaired Student's t-test. Values of P < 0.05 were considered significant.
Results
Expression of nAChRs subunits
We examined the mRNA expression levels of nAChR subunits both in hESCs/hiPSCs and human somatic stem cells (cell line). We also used the commercial human cDNA samples from various organs to compare. The mRNA expression levels of nAChR subunits in hiPSCs were higher than in all organs other than the brain (Fig. 1). Expression levels of the α4, α7, and β4 subunits were ∼1/6, 1/2, and 10 times the levels in brain, respectively (Fig. 1). Expression levels in hiPSCs were much higher than in adult MSCs.
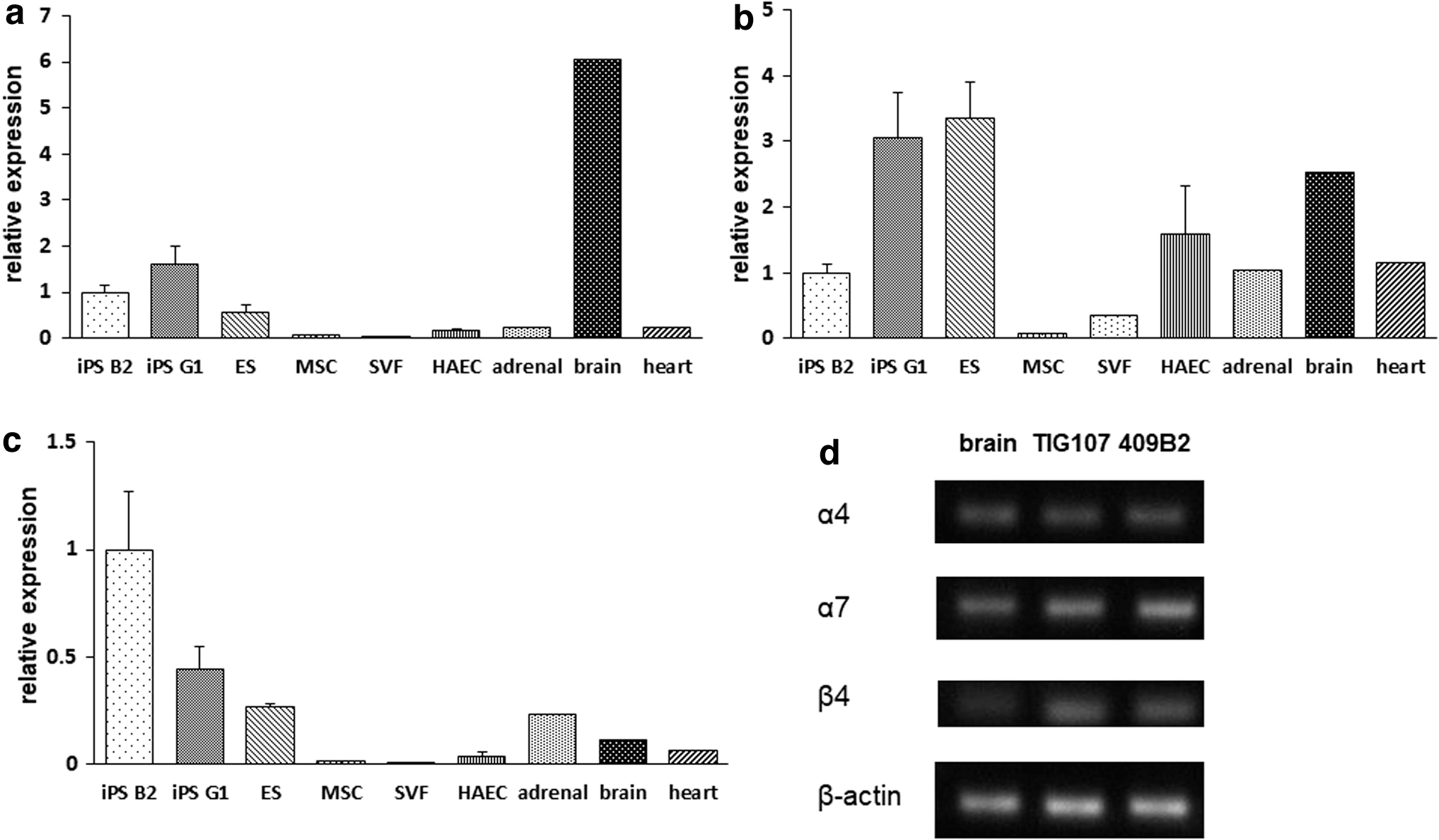
Relative gene expression of nicotine receptor subunits in various tissues and cells.
GO analysis
A total of 21,448 genes were included in this analysis, whose fold change in all genes is given in Supplementary Table S2. Eight hours after nicotine administration, 66 genes were significantly upregulated and 94 were significantly downregulated in nicotine-treated cells as compared with untreated controls (Fig. 2a). The most enriched GO terms were “response to stimulus” in biological process (BP), “ion binding” in molecular function (MF), and “plasma membrane” in cellular component (CC). Among genes involved in the nervous system, a group of genes related to immune responses, including “defense response” and “response to wounding,” as well as “neurological system process” were also significantly changed (Fig. 3a). At 24 h, 112 genes were significantly upregulated and 109 genes were significantly downregulated (Fig. 2b).

Scatter plot comparing 1.5-fold upregulated and downregulated genes (black dots) in nicotine-treated and untreated cells. The gray dots indicate no significant fold changes in gene expression.
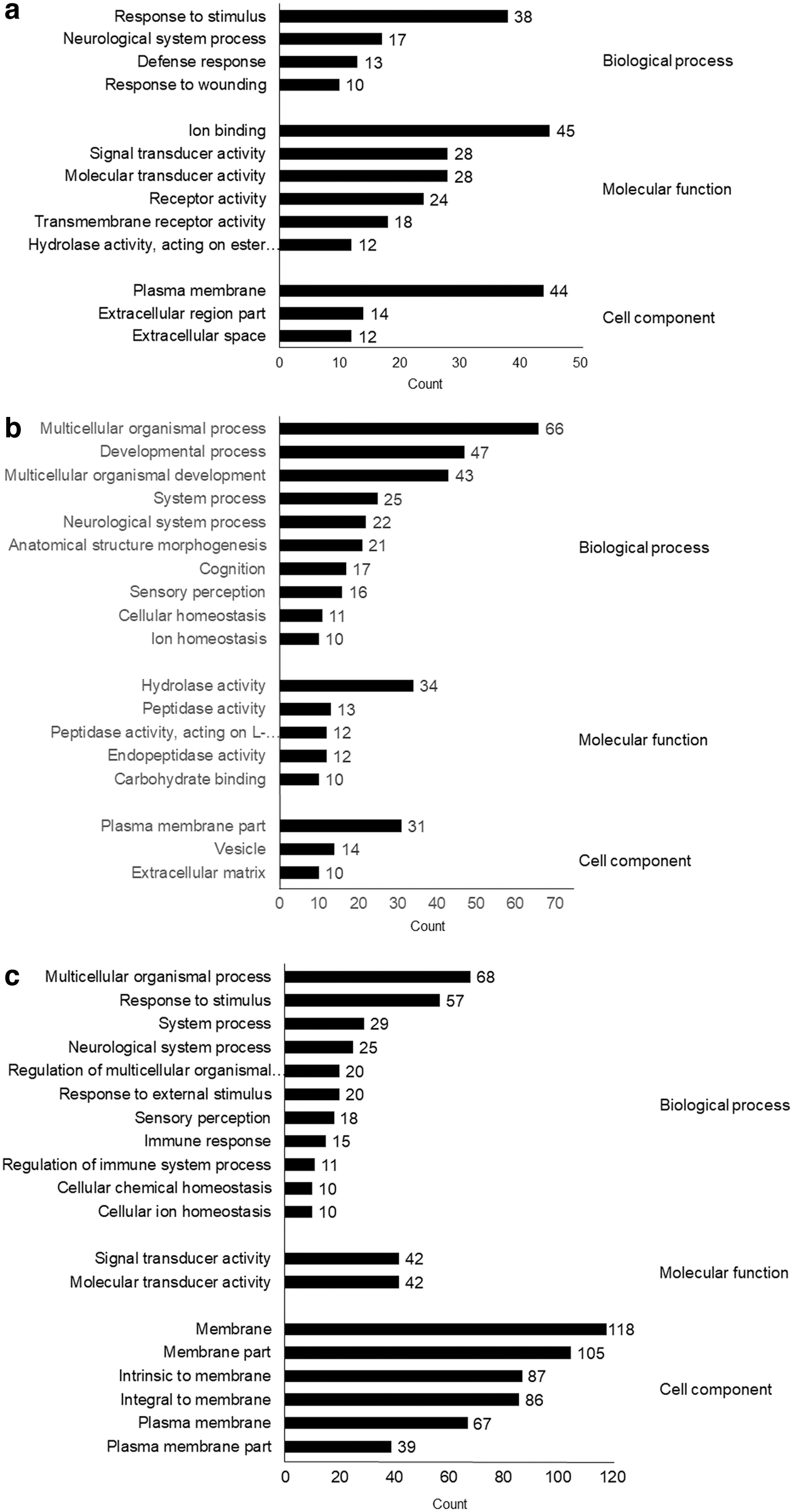
Functional GO analysis of biological processes, molecular function, and cellular components. The number of genes in each GO term is shown on the right side of each bar graph.
The most enriched GO terms were “multicellular organismal process” in BP, “hydrolase activity” in MF, and “plasma membrane part” in CC. As was seen at 8 h, significant changes were observed in “neurological system processes,” as well as “cognition,” “sensory perception,” and “ion homeostasis” (Fig. 3b). At 72 h, 105 genes were significantly upregulated and 181 genes were significantly downregulated (Fig. 2c). The most enriched GO terms were “multicellular organismal process” in BP, “signal transducer activity” in MF, and “membrane” in CC. As at 8 h, a group of genes related to immunity, including “immune response” and “regulation of immune system process,” were also significantly changed. As was seen at 24 h, neurological system genes, including “neurological system process” and “sensory perception,” as well as ion-related genes under the term “cellular ion homeostasis” were also significantly changed (Fig. 3c).
Enrichment analysis revealed that, at 8 h, the highest ES was observed in the defense response gene group (ES = 1.96), followed by the gene group related to calcium ion binding (ES = 1.81) (Fig. 4 and Table 1). At 24 h, the highest ES was detected in the gene group related to metal binding (ES = 3.03), followed by the gene group related to sensory perception (ES = 1.69). Genes related to immune responses were not changed. At 72 h, the ES for metal binding was overwhelmingly high at 9.59, and genes that included metallothionein (MT) were decreased.
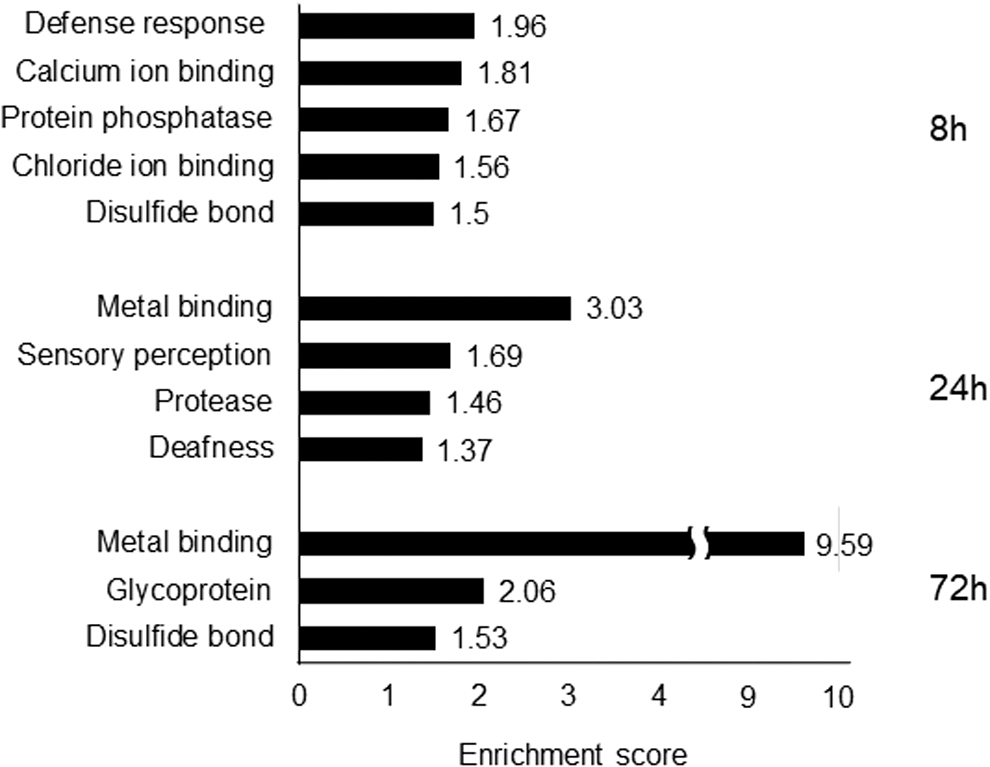
Functions showing significantly increased enrichment scores after nicotine exposure for 8, 24, or 72 h.
Gene Symbols in Enrichment Analysis
MT gene expression and ROS production after nicotine administration
Because the enrichment analysis revealed MT gene expression to be low 72 h after nicotine application and the expression of genes involved in cell proliferation varied, we focused on the antioxidant stress effect of MT and assessed mRNA expression levels of MT and ROS production in hiPSCs in the presence of nicotine or NAC, a known antioxidant (n = 3; Fig. 5). qRT-PCR analysis of MT expression after 72 h of exposure to nicotine showed significantly decreased expression of MT1G and MT1H, and similar decreases were seen in the presence of NAC. At the same time, ROS production was significantly decreased, despite the reduced levels of MT gene expression (Fig. 6a).

Metallothionein mRNA expression in hiPSCs after 72 h in the presence of 1% DMSO (control: black bar), nicotine (gray bar) or NAC (white bar). N = 3 in each condition. DMSO, dimethyl sulfoxide; NAC, N-acetylcysteine.
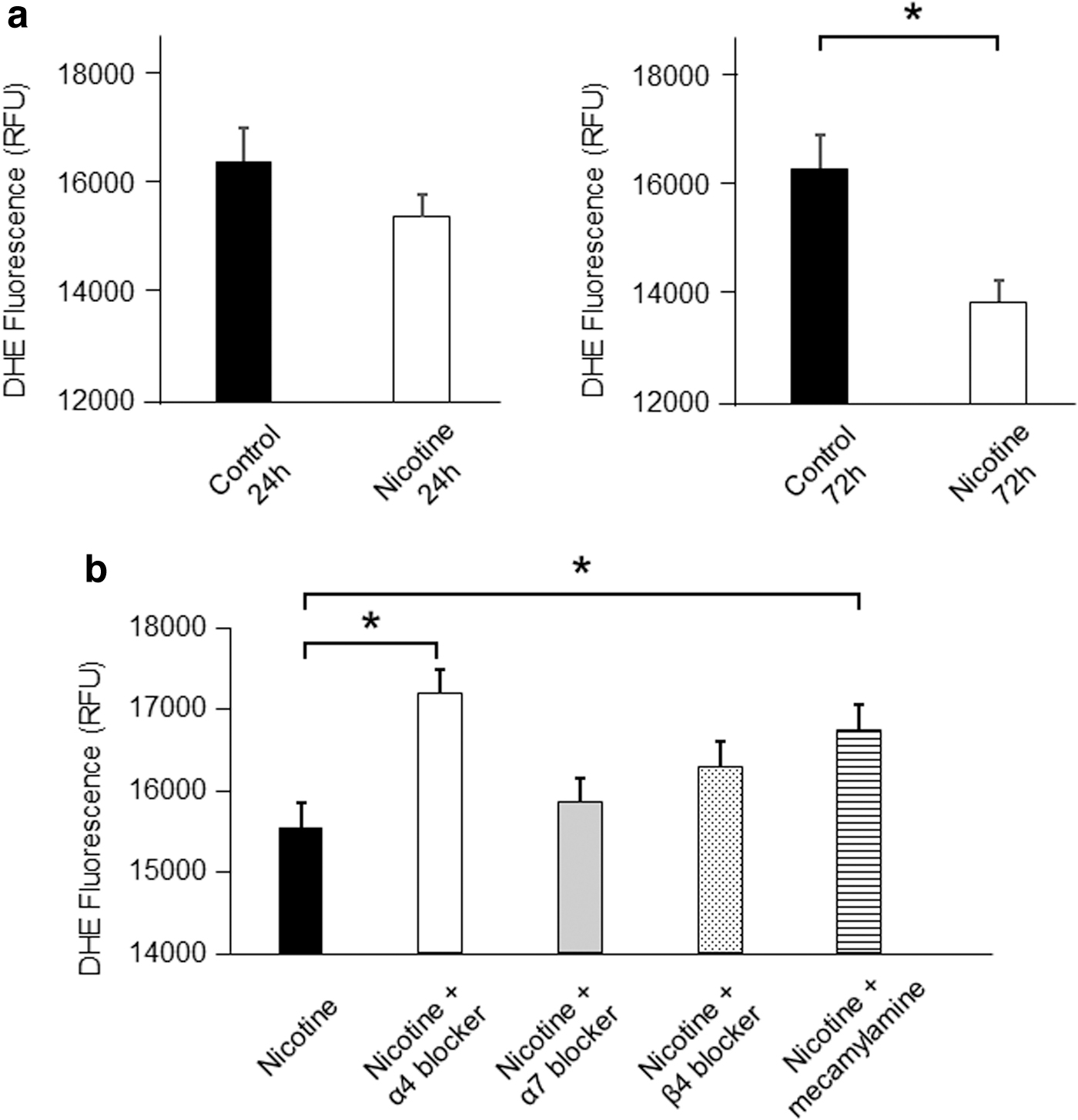
In addition, ROS production in the presence of nicotine plus mecamylamine, a nonselective nAChR inhibitor, or an α4 nAChR subunit antagonist, 3-dihydro-1-(9-methyl-9-azabicyclo[3.3.1]non-3-yl)-2H-indol-2-one, was significantly lower than with nicotine alone (nicotine vs. nicotine+mecamylamine, P < 0.05; and nicotine vs. nicotine+α4 nAChR subunit antagonist, P < 0.05; Fig. 6b). By contrast, ROS production was not significantly changed in the presence of nicotine plus an α7 or β4 nAChR subunit antagonist as compared with nicotine alone. Given that MT gene expression reportedly leads to reductions in ROS production [16,17], we hypothesized that nicotine may contribute to reducing ROS production and affect cell proliferation via the α4 nAChR subunit.
Using MTT assays, we compared estimated cell numbers among untreated control hiPSCs and those incubated in the presence of nicotine or nicotine with α4 nAChR subunit antagonist. We found that nicotine increased cell numbers as compared with control, which was canceled by an α4 nAChR subunit antagonist (Fig. 7). In addition, we checked the genes in Supplementary Table S2 including the term “proliferation” in their description under the nicotine treatment in 72 h, the number of which was 12. Nine of them were correlated with increased cell proliferation, which tended to be significant in comparison with control by binomial test (P = 0.073).
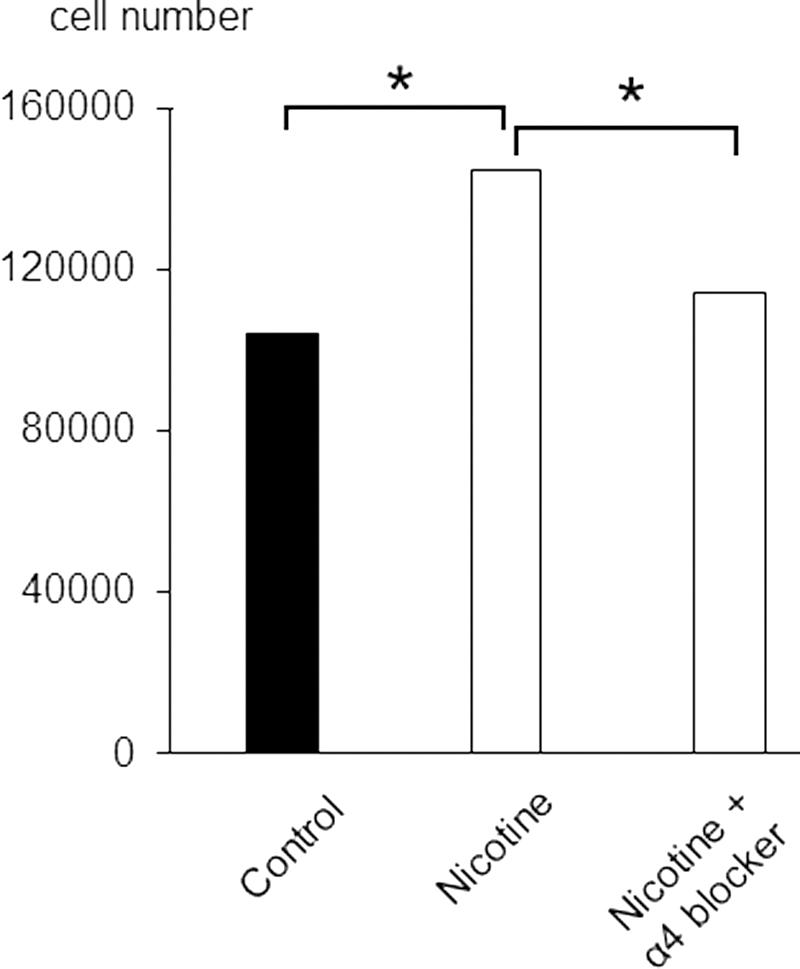
Estimated hiPSC counts after 72 h in the presence of 1% DMSO (Control) nicotine or nicotine with α4 nicotinic acetylcholine receptor subunit antagonist. N = 6 in each condition.
Changes of genes related to differentiation
In the cDNA microarray data, two markers related to differentiation endoderm and mesoderm, as well as ectoderm lineage markers, were significantly changed. There was a −1.63-fold change in BMP4, which is essential for differentiation to ectoderm, and a 1.76-fold change in SNAI2, which is important for epithelial–mesenchymal transition.
Discussion
In this study, we first demonstrated the mRNA expression of nAChRs in hiPSCs. nAChRs were more abundant in hiPSCs than in MSCs and all adult organs except the brain. Of note, expression of the β4 subunit was higher in hiPSCs than in the human brain. In addition, cDNA microarray, GO, and enrichment analyses showed nicotine had multiple effects on hiPSCs, including change of (1) genes associated with immune responses and inflammation, (2) genes involved in the development of diseases associated with abnormalities of the nervous system, (3) genes involved in differentiation into cartilage, and (4) genes involved in inhibiting or promoting cell proliferation. In that regard, the results of our MTT assays suggest nicotine promoted cell proliferation under our experimental conditions. ROS production was significantly reduced after 72 h of nicotine exposure, and that effect was canceled by an α4 nAChR antagonist, 3-dihydro-1-(9-methyl-9-azabicyclo[3.3.1]non-3-yl)-2H-indol-2-one. This suggests that α4 subunit may mediate the signal leading to the decreased ROS production induced by nicotine.
The cDNA microarray analysis and qRT-PCR showed that MT gene expression levels were significantly decreased in cells exposed to nicotine. MT reportedly reduces ROS production [16,17]. We therefore, hypothesized that nicotine would increase ROS production in hiPSCs; however, the result was the opposite. ROS production was significantly decreased in cultured hiPSCs after 72 h of nicotine exposure. This decrease in ROS production may underlie our finding that genes involved in immune responses and inflammation like IL23R and IL25 were suppressed after nicotine application (Table 1) [18]. These results also suggest that declines in MT gene expression do not contribute to the observed decrease in ROS production. Considering that overexpression of MT1G reportedly inhibits proper differentiation of myeloid cells [19], changes in MT gene expression levels may influence hiPSC proliferation/differentiation.
Nicotine concentration of 10 μmol/L used in this study is ∼1,600 ng/mL, which is well above the average blood nicotine concentration of 200 ng/mL seen in female smokers [20] and is extremely higher than the average blood nicotine concentration of 0.73 ng/mL measured in nonsmokers [21]. That may mean our findings more closely reflect the effects of smoking on hiPSCs, and not the effects of nicotine synthesized in vivo on hiPSCs. As the concentration of nicotine in previous reports varies greatly, we had already carried out the experiment about the dose dependency from 10−3 to 10−7 mol/L for the cell proliferation by nicotine in human iPSCs, showing human iPSCs proliferated the most at a concentration of 10−5 mol/L (10 μmol/L) (Supplementary Fig. S1). We, therefore, adopted 10 μmol/L for this study.
In earlier reports, low nicotine concentrations such as 10−8 mol/L nicotine or 300 nmol/L increased cell proliferation among ESC-derived endothelial cells and mouse iPSCs [14,22]. At high concentrations, such as 100 μmol/L to 1 mmol/L, nicotine inhibited myofibroblast differentiation from human gingival and lung fibroblasts and the differentiation of bone marrow MSCs into cartilage [23 –25]. High nicotine such as 1 to 10 μmol/L concentrations also suppressed proliferation of mouse ESCs [26]. These reports suggest that although high concentrations of nicotine have adverse effects on cells, low levels of nicotine may promote cell survival.
It was previously reported that nicotine increases ROS production in undifferentiated neural stem cells [5]. Although excessive increases in ROS are cytotoxic and suppress BMP1 expression, which may affect skeletal cell differentiation [27], moderate increases are thought to promote iPSC differentiation [28,29]. On the contrary, the capacity of mouse iPSCs to differentiate into mesodermal or neural progenitor cells was unaffected by nicotine [14]. In our study, BMP1 was suppressed (Table 1) despite decreased ROS. In addition, our cDNA microarray data indicate that, with the exception of BMP4 and SNAI2, differentiation markers were not significantly affected by nicotine. In addition, our ES analysis showed that PCSK2 and CAPN9 expression, which may be related to tumorigenesis [30,31], was reduced. The nicotine-induced reduction of ROS may have suppressed cell differentiation and contribute to the maintenance of hiPSCs in an undifferentiated state. Consistent with that idea, we observed nicotine application increased in the number of undifferentiated hiPSCs (Supplementary Fig. S2).
Conclusion
Undifferentiated hiPSCs express nAChRs, which may contribute to decreasing oxidative stress and proliferation, especially via the α4 subunit, as nicotine decreases ROS levels and enhances hiPSC proliferation. Although further studies are needed to more clearly define the function of nAChRs in hiPSCs, our study provides new insight into the significance of nAChRs on human stem cells and the potential effects of smoking on fertilized human ova.
Footnotes
Acknowledgments
The iPS cell line TIG107 is a kind gift from Dr. Kazutoshi Takahashi. The authors thank Yoshie Fukuchi for her technical assistance on cell culture and RT-PCR.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Smoking Research Foundation. This study was also supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant No. JP 22K08651.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.