
Abstract
Transamniotic stem cell therapy (TRASCET) with mesenchymal stem cells (MSCs) can attenuate placental inflammation and minimize intrauterine growth restriction (IUGR). We sought to determine whether MSC-based TRASCET could mitigate fetal cardiopulmonary effects of IUGR. Pregnant Sprague–Dawley dams were exposed to alternating 12-h hypoxia (10.5% O2) cycles in the last fourth of gestation. Their fetuses (n = 155) were divided into 4 groups. One group remained untreated (n = 42), while three groups received volume-matched intra-amniotic injections of either saline (sham; n = 34), or of syngeneic amniotic fluid-derived MSCs, either in their native state (TRASCET; n = 36) or “primed” by exposure to interferon-gamma and interleukin-1beta before administration in vivo (TRASCET-primed; n = 43). Normal fetuses served as additional controls (n = 30). Multiple morphometric and biochemical analyses were performed at term for select markers of cardiopulmonary development and inflammation previously shown to be affected by IUGR. Among survivors (75%; 117/155), fetal heart-to-body weight ratio was increased in both the sham and untreated groups (P < 0.001 for both) but normalized in the TRASCET and TRASCET-primed groups (P = 0.275, 0.069, respectively). Cardiac b-type natriuretic peptide levels were increased in all hypoxia groups compared with normal (P < 0.001), but significantly decreased from sham and untreated in both TRASCET groups (P < 0.0001–0.005). Heart tumor necrosis factor-alpha levels were significantly elevated in sham and TRASCET groups (P = 0.009, 0.002), but normalized in the untreated and TRASCET-primed groups (P = 0.256, 0.456). Lung transforming growth factor-beta levels were significantly increased in both sham and untreated groups (P < 0.001, 0.003), but normalized in both TRASCET groups (P = 0.567, 0.303). Similarly, lung endothelin-1 levels were elevated in sham and untreated groups (P < 0.001 for both), but normalized in both TRASCET groups (P = 0.367, 0.928). We conclude that TRASCET with MSCs decreases markers of fetal cardiac strain, insufficiency, and inflammation, as well as of pulmonary fibrosis and hypertension in the rodent model of IUGR.
Introduction
Intrauterine growth restriction (IUGR) is the second most common cause of neonatal morbidity after prematurity [1,2]. Cardiopulmonary complications are among the most impactful consequences of IUGR and can persist well into adulthood [3,4]. These include cardiomyopathy, pulmonary hypertension, and pulmonary fibrosis [5 –7]. An overly hypoxic fetal environment derived from placental insufficiency, in turn caused by undue placental inflammation, is thought to be central to the pathophysiology of cardiopulmonary complications of IUGR [8].
Transamniotic stem cell therapy (TRASCET) with mesenchymal stem cells (MSCs) has been recently shown to reverse some of the core effects of experimental IUGR, apparently due to the inordinate capacity of MSCs to modulate inflammation [9 –11]. Utilizing the hypoxia rodent model of IUGR, we sought to determine whether MSC-based TRASCET has any impact on select morphometric variables and markers of fetal cardiopulmonary development previously shown to be affected by this disease.
Materials and Methods
This study was approved by the Boston Children's Hospital Institutional Animal Care and Use Committee under protocol #20-07-4182.
Donor cell procurement and processing
Donor cells were derived from banked amniotic fluid-derived MSCs (afMSCs) that we had previously harvested from healthy Lewis rat (a syngeneic strain) dams as previously described [12]. In brief, a midline laparotomy was performed just before term on gestational day 21 (E21, term = E21-23). Amniotic fluid samples were procured through a 30G needle on a 1-mL syringe (both from Becton Dickinson, Franklin Lakes, NJ). After initial expansion, the mesenchymal progenitor identity of the cells was confirmed by fluorescence-activated cell sorting analysis using a Vantage SE cell sorter (Becton Dickinson).
Unconjugated mouse monoclonal antibodies validated for use in rats were used, namely against CD29, CD44, CD45, and CD90 (all from Becton Dickinson), as well as a purified CD73 (Becton Dickinson) conjugated with an anti-mouse IgG1 against purified CD73 (BioLegend, San Diego, CA). Nonspecific cell staining was excluded with mouse isotype immunoglobulin controls. All donor cells were labeled with green fluorescent protein (GFP) using plasmid pMIG #9044 (Addgene, Cambridge, MA) and Lipofectamine LTX with PLUS Reagent (Invitrogen, Grand Island NY).
Cells were transfected in antibiotic-free mesenchymal media [450 mL Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Waltham, MA) +50 mL fetal bovine serum (Life Technologies, Chicago, IL)] for 6 h at 37°C, and then refed with fresh M-20 media during expansion thereafter. GFP positivity was confirmed using the EVOS® FL Color Imaging System microscope (Life Technologies).
Donor cell priming
A subset of donor afMSCs were exposed to a modified culture media consisting of the standard media added with 30 ng/mL of interferon-gamma and 5 ng/mL of interleukin-1beta (both from PeproTech, Cranbury, NJ) for 24 h before injection in vivo to “prime” the cells toward an enhanced anti-inflammatory phenotype, as previously described [13,14].
IUGR model and intra-amniotic injections
IUGR was reproduced in 12 age-matched, time-dated pregnant Sprague–Dawley dams (Charles River Laboratories, Inc., Wilmington, MA) by exposure to controlled hypoxia, as previously described [15]. Briefly, dams were placed in a hypoxia chamber (Pro Ox P360; Biospherix, Parish, NY) on E15 and submitted to alternating cycles of 10.5% O2 every 12 h, with normoxia taking place overnight during their more active period. On E17, dams were divided into four groups: an untreated group had no further manipulations (n = 42 fetuses); three groups received volume-matched (50 μL) intra-amniotic injections of either saline (sham; n = 34) or a suspension of either regular afMSCs (TRASCET; n = 36) or primed afMSCs (TRASCET-primed; n = 43). All injected cells were administered as a suspension of 2 × 106 cells/mL in phosphate buffered saline.
To complete the injection procedure, dams were removed from the hypoxia chamber for less than 1 h. All fetuses from a single dam received the same treatment, which was executed as previously described [9 –11]. In brief, anesthesia was induced through chamber-inhaled isoflurane (Abbott, Abbott Park, IL) and maintained through nose cone at 2%–4% in 100% O2. The dam underwent a midline laparotomy, allowing evisceration and inspection of the bicornate uterus. Injections were performed under direct visualization to prevent damage to the fetus or umbilical cord. A 33G noncoring needle fitted to a 100-μL syringe (both from Hamilton Company, Reno, NV) was used for each injection. Upon completion, the uterus was returned to the abdomen and the laparotomy was closed in two layers with 3-0 Vicryl (Ethicon, Somerville, NJ) and 5-0 Monocryl (Ethicon) running sutures. Animals were allowed to recover before reintroduction into the hypoxia chamber. Buprenorphine (Zoopharm, Windsor, CO) was utilized for postoperative analgesia.
Uteroplacental tissue extraction and gross morphometrics
Dams underwent euthanasia just before term on E21 by CO2 inhalation. The previous midline laparotomy was reopened and the uterus eviscerated. All subsequent dissection was done under 2.5 × loupe magnification as previously described [9 –11]. Briefly, a uterine myotomy was performed and fetuses were delivered en caul with the placenta bluntly detached from the uterine wall. The amniotic sac was then disrupted and removed, and the fetuses were dried. Their umbilical cords were clamped and cut, and each fetus was weighed. The gestational membranes were dissected sharply from the chorionic plate of the placenta, and the placenta was weighed. Both fetal and placental weights were calculated as ratios controlled by maternal weight to account for variance between dam sizes, which were not statistically significantly different, however. Placental efficiency, a variable commonly measured in IUGR studies, was calculated as a ratio between the fetal and placental weights [16].
Fetal dissection was also done under 2.5 × loupe magnification. The fetal heart was collected in whole through a sternotomy, the major vessels were trimmed so as to include only cardiac tissue in the specimen. The lungs were procured through the same sternotomy, with each side dissected at the hilum and care was taken to trim any diaphragmatic attachments. The heart and lung samples were then weighed, and heart-to-body weight ratios were calculated for each fetus [17]. Fetal lung-to-body weight ratios were not calculated due to variance due to trauma during total lung procurement limiting reliability. After weighing, samples were flash frozen at −80°C and stored.
An additional 30 normal fetuses from 3 Sprague–Dawley dams not exposed to hypoxia were procured on E21 and underwent the same measurements described above to serve as normal controls.
Enzyme linked immunosorbent assay
Fetal heart and lung samples were homogenized, standardized by weight, and submitted to enzyme-linked immunosorbent assay (ELISA) analyses in duplicate for different biomarkers of cardiac and pulmonary pathology previously described in models of IUGR [18,19]. For the heart samples, cardiac inflammation was assessed by tumor necrosis factor-alpha (TNF-α) (Abcam, Cambridge, MA) and heart strain was assessed by B-type Natriuretic Peptide (BNP) (Abcam). For the lung samples, pulmonary hypertension was assessed by Endothelin-1 (ET-1) and pulmonary fibrosis by transforming growth factor-beta (TGF-β) (both Abcam).
Statistical analyses
Survival was compared by Fisher's exact test. Morphometric and ELISA data were compared by nonparametric Wilcoxon rank-sum test. Significance was set at P < 0.05. All analyses were done using IBM SPSS software (SPSS version 21.0; IBM, Armonk, NY).
Results
Overall survival from hypoxia was 75% (117/155), but significantly lower in all three injection groups than in the untreated group (P = 0.003–0.005). There was no difference in survival between sham and either TRASCET group (P = 0.99 for both), or between both TRASCET groups (P = 0.812).
Gross morphometrics
Placental efficiency was significantly lower than normal in untreated, sham, and TRASCET-primed groups (P < 0.001–0.004), but normalized in the TRASCET group (P = 0.915; Table 1; Fig. 1). Both TRASCET groups had significantly improved placental efficiency compared with both sham and untreated groups (P < 0.001–0.012). There was no difference in placental efficiency between the TRASCET groups (P = 0.088).
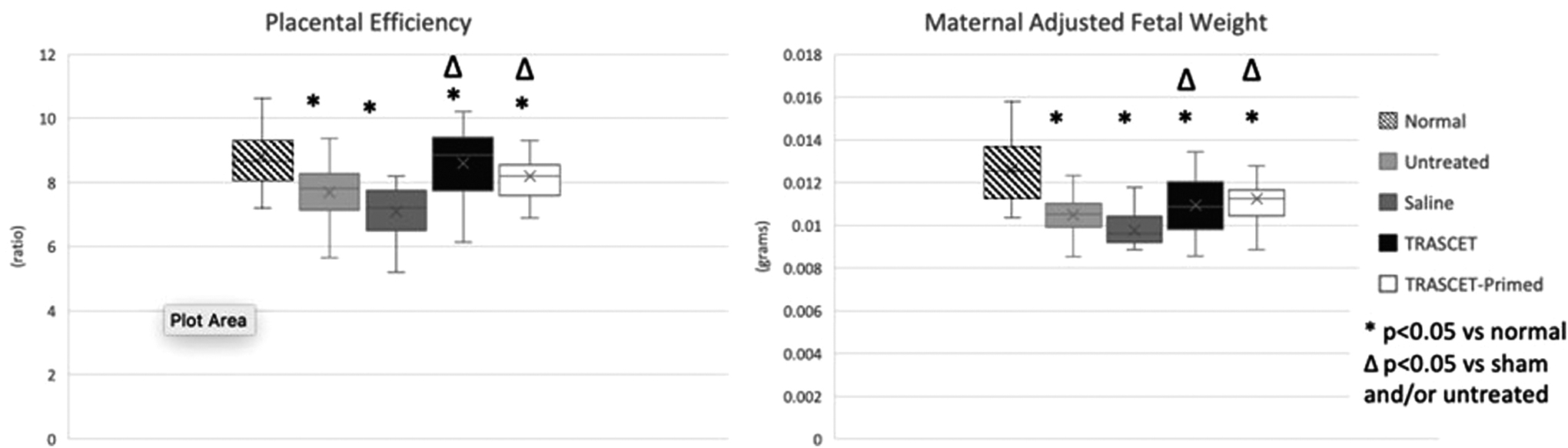
Placental efficiency (placenta-to-fetal weight ratio) and maternal-adjusted fetal weight. Both TRASCET groups had significantly improved placental efficiency compared with the sham and untreated groups (P < 0.001–0.012). TRASCET and TRASCET-primed groups had significantly higher maternal-adjusted fetal weight than sham (P < 0.001–0.006). There was no difference in maternal-adjusted fetal weight or placental efficiency between both TRASCET groups (P = 0.486, 0.088, respectively). TRASCET, Transamniotic stem cell therapy.
Fetal and Placental Gross Morphometrics and Protein Levels of Select Cardiac and Lung Biomarkers
Data presented as median (interquartile range).
BNP, b-type natriuretic peptide; ET-1, endothelin-1; TGF-β, transforming growth factor-beta; TNF-α, tumor necrosis factor-alpha; TRASCET, transamniotic stem cell therapy.
Maternal-adjusted fetal weight was significantly lower in all hypoxic groups compared with normal controls (P < 0.001 for all). However, both TRASCET and TRASCET-primed groups had significantly higher maternal-adjusted fetal weight than sham (P < 0.001–0.006). There was no difference in maternal-adjusted fetal weight between both TRASCET groups (P = 0.486). Maternal-adjusted placental weights were similar to normal controls in all groups (P = 0.0129–0.872).
The fetal heart-to-body weight ratio was normalized in the TRASCET and TRASCET-primed groups (P = 0.275, 0.069; Fig. 2). However, that ratio was significantly elevated in both the sham and untreated groups when compared with normal (P ≤ 0.001 for both).

Gross cardiac morphometrics by fetal heart-to-body weight ratio, which was normalized in the TRASCET and TRASCET-primed groups (P = 0.275, 0.069, respectively), but was significantly elevated in both the sham and untreated groups when compared with normal (P ≤ 0.001 for both).
Cardiac ELISA
Cardiac TNF-α levels were significantly elevated in the sham and TRASCET groups (P = 0.009, 0.002, respectively; Fig. 3) but normalized in the untreated and TRASCET-primed groups (P = 0.256, 0.456, respectively). The TRASCET group's TNF-α levels were significantly elevated compared with TRASCET-primed (P = 0.002).
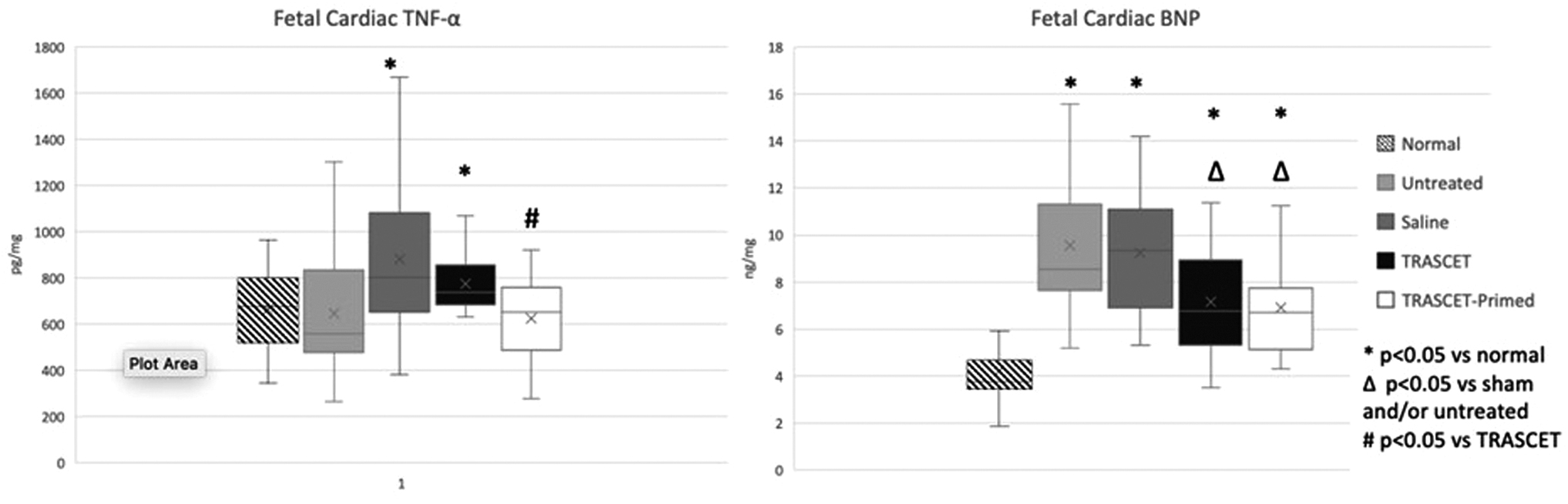
Fetal cardiac TNF-α and BNP levels at term. Cardiac TNF-α levels were significantly elevated in the sham and TRASCET groups (P = 0.009, 0.002, respectively) but normalized in the untreated and TRASCET-primed groups (P = 0.256, 0.456, respectively). Cardiac BNP levels were elevated in all hypoxia groups compared to normal (P < 0.001 for all) but were significantly decreased in both TRASCET and TRASCET-primed groups compared to the sham and untreated groups (P < 0.001–0.005). BNP, b-type natriuretic peptide; TNF-α, tumor necrosis factor-alpha.
Cardiac BNP levels were elevated in all hypoxia groups compared with normal (P < 0.001 for all; Fig. 3). However, cardiac BNP levels were significantly decreased in both TRASCET and TRASCET-primed groups compared with sham and untreated groups (P < 0.001–0.005).
Pulmonary ELISA
Lung TGF-β levels were significantly increased in both sham and untreated groups (P < 0.001, 0.003, respectively; Fig. 4). However, lung TGF-β levels were normalized in both TRASCET and TRASCET-primed groups (P = 0.567, 0.303, respectively). TGF-β levels were significantly lower in fetal lungs of the TRASCET-primed group compared with the TRASCET group (P = 0.03).
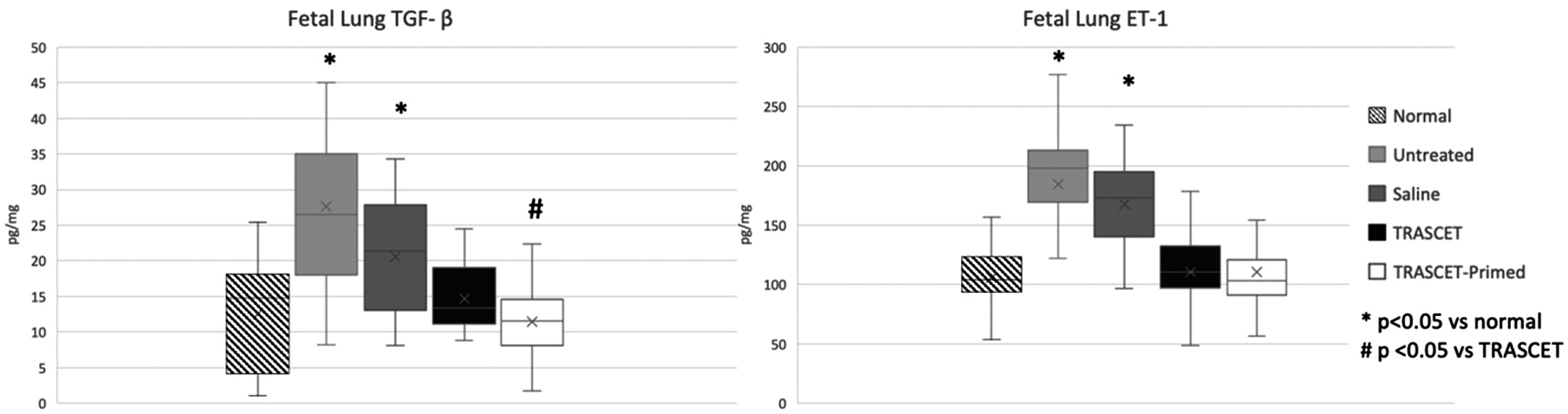
Fetal lung TGF-β and ET-1 levels at term, both of which were elevated in the sham and untreated groups when compared to normal controls (P < 0.001–0.003), but normalized in both TRASCET and TRASCET-primed groups (P = 0.303, 0.928, respectively). ET-1, endothelin-1; TGF-β, transforming growth factor-beta.
Lung ET-1 levels were elevated in sham and untreated groups when compared with normal controls (P < 0.001 for both; Fig. 4), but normalized in both TRASCET and TRASCET-primed groups (P = 0.367, 0.928, respectively). Lung ET-1 levels were similar between the two TRASCET groups (P = 0.423).
Discussion
Beyond reduced neonatal weight and height, the two foremost complications of IUGR are cardiopulmonary dysfunction and neurodevelopmental delay, both often lasting long into adulthood [3,5 –7,20]. Combined, the present data suggest that fetal cardiopulmonary development can be protected at least in part by TRASCET with MSCs in a hypoxia-based model of IUGR, with normalization of fetal heart-to-body weight ratio along with reductions in markers of cardiac strain, cardiac inflammation, pulmonary hypertension, and pulmonary fibrosis. These findings are consistent with previous results involving other facets of this overarching experimental design and continue to support the notion that TRASCET with MSCs could become a viable and practical treatment strategy for IUGR [9 –11].
Interestingly, however, in this experiment there were few differences between the primed and unprimed MSCs for most of the outcomes measured, with the sole exceptions being the direct markers of inflammation and fibrosis, namely cardiac TNF-α and lung TGF-β, both of which were more significantly improved in the TRASCET-primed group when compared with “plain” TRASCET. Prior studies focusing on other variables of IUGR have shown a greater impact from primed MSCs [9 –11]. Although there was clear evidence of the effects of MSC-based TRASCET in modulating inflammation in this study as well, the fact that primed MSCs did not enhance the effects of TRASCET in markers of pulmonary hypertension or cardiac strain would suggest that the MSCs are working in more ways than exclusively in the realm of inflammation. Indeed, MSCs are also known to be antiapoptotic, proangiogenic, and to promote tissue growth in a variety of settings [21 –23]. These properties may have played a greater role in the setting of cardiopulmonary pathology associated with IUGR than the anti-inflammatory and immunomodulatory affects alone.
Further study on the mechanisms of action of the MSCs in this model will hopefully shed more light on the processes at play and possibly optimize them before eventual clinical translation. At the same time, the fact that primed MSCs were equally as effective in improving all cardiopulmonary outcomes hereby measured, in addition to being superior at improving the associated inflammation and fibrosis, not to mention the results of prior studies, still supports them being favored in TRASCET for IUGR.
Expectedly, this study was limited by the very small organ sizes of fetal rats, which restrict one's ability to perform concomitant histologic and/or functional analyses beyond the protein and morphometric ones that we performed. Even the number of biomarkers that we could assess through ELISA was limited in light of the need for duplicate measurements. The choice of these biomarkers was based on them having been previously shown to be affected in this and other models of IUGR, by different groups, so that the results could be more dependably interpreted. Yet another limitation of these small fetal samples was the fact they prevented us from assessing the eventual presence of the donor cells in the fetal heart and lung. This was not anticipated, as we did label the donor cells with GFP in the hopes of adding that analysis to the data. Of note, other studies on MSC-based TRASCET in the rat model have shown that MSCs home in the largest numbers to the placenta and fetal bone marrow, with negligible levels in the fetal heart and lung, pointing to the placenta as the primary site or donor MSC action in this model [24 –26].
Before considering any of the additional analyses highlighted by the discussion of these limitations, we needed to first determine whether there was any cardiopulmonary effect at all from TRASCET for IUGR, hence this experiment. Besides justifying these additional studies, the present data also substantiate the need for functional hemodynamic analyses, which have been proven viable in the nitrofen model of congenital diaphragmatic hernia submitted to TRASCET [27]. We are currently investigating the mechanisms behind MSC-based TRASCET for IUGR and other diseases for which this therapeutic principle has proven applicable experimentally.
Footnotes
Author Disclosure Statement
No competing financial interests exist from any of the authors.
Funding Information
This work was funded by the Kevin and Kate McCarey Fund for Surgical Research at Boston Children's Hospital.