
Abstract
Mesenchymal stem cells (MSCs) play an essential role in multiple physiological processes in vivo and a promising cell-based therapy for various diseases. Nonetheless, MSCs suffer from senescence with expansion culture, leading to a limitation for their clinical application. Recently, it was reported that small extracellular vesicles (sEVs) are involved in regulation of senescence in tumor cells and fibroblasts. However, the biological roles of sEVs in senescent MSCs (Sen MSCs) are poorly understood. In this study, we established a replicative senescence model of MSCs by successive passages and compared the phenotypic changes between presenescent MSCs (Pre-Sen MSCs) and Sen MSCs and found that Sen MSCs exhibited a diminished adipogenic and osteogenic differentiation potential and elevated senescence-associated secretory phenotype levels. In addition, we found that sEV secretion was increased in Sen MSCs, and inhibition of sEV secretion led to apoptosis, DNA damage, and decreased cell viability, suggesting that increased sEV secretion plays an important role in maintaining Sen MSC homeostasis. To further investigate the molecular mechanisms, metabolomic profiling of Pre-Sen MSC-derived sEVs (Pre-Sen-sEVs) and Sen MSC-derived sEVs (Sen-sEVs) was performed. The results showed that lipid metabolites were significantly increased in Sen-sEVs and these significantly upregulated lipid metabolites were shown to be toxic for inducing cellular senescence and apoptosis in previous studies. Kyoto Encyclopedia of Genes and Genomes analysis revealed enrichment of differential metabolites between Pre-Sen-sEVs and Sen-sEVs mainly in 25 signaling pathways, of which 21 metabolic pathways have been shown to be closely associated with senescence. Taken together, our findings suggested that increased sEV secretion maintains Sen MSC homeostasis, at least in part, by excreting harmful lipids, thus providing new insights into the regulation of senescence by sEVs.
Introduction
Mesenchymal stem cells (MSCs), identified as multipotent stromal cells, play a vital role in various aspects of tissue development, function, maintenance, and regeneration in vivo [1]. They have long been considered as a source of cells for tissue regeneration due to their potential for self-renewal and multilineage differentiation. MSCs also have complex paracrine activities that allow them to coordinate multiple physiological pathways such as immune regulation, innervation, and revascularization [2]. Furthermore, coupled with their low immunogenicity and wide source [3], MSCs are being intensively investigated as a promising cell-based therapy in clinical trials [4,5].
Nonetheless, with expansion culture, MSCs gradually lose their function and stemness and eventually become senescent cells [6]. Senescent MSCs (Sen MSCs) exhibit the typical features of cellular senescence—irreversible cell cycle arrest and activation of the senescence-associated secretory phenotype (SASP) program, severely limiting their clinical applications [7].
Small extracellular vesicles (sEVs) are lipid bilayer membrane vesicles <200 nm in diameter [8]. MSC-derived sEVs, due to their outstanding advantages of abundant functional cargos [9], similar therapeutic effects to the parent cells [10], low immunogenicity, and high biosafety [11], have been studied mainly for their clinical therapeutic functions, such as the use of sEVs as cell-free therapeutic agents and engineered carriers modified by siRNA, targeted peptides, or thermosensitive hydrogel for the treatment of multiple diseases [12 –16]. However, the biological functions and related mechanisms of sEVs in senescent sEV-secreting cells have not been well understood, especially in Sen MSCs.
Therefore, we investigated the role of sEVs in regulation of senescence in a replicative senescence model of MSCs and analyzed the molecular mechanisms by metabolomic profiling. Our findings revealed that sEVs may play an important role in maintaining Sen MSC homeostasis by excreting harmful lipids.
Materials and Methods
Cell culture
Human adipose-derived MSCs were isolated from human adipose tissues of three different healthy volunteers, as previously reported [17]. All human adipose tissues were obtained according to the procedures approved by the Ethics Committee at Peking Union Medical College and the Chinese Academy of Medical Sciences. MSCs were resuspended in 12 mL of culture medium and seeded in a 75-cm2 flask, then maintained in a 37°C incubator with 5% carbon dioxide and passaged using trypsin/ethylenediaminetetraacetic acid.
Induction of replicative senescence in MSCs
MSCs were cultured under the above conditions and passaged at a ratio of 1:2 after reaching 90% confluence, and the culture medium was changed every 2 days. MSC senescence was evaluated using the senescence β-galactosidase staining kit (Beyotime, Shanghai, China) following the manufacturer's instructions, and images were taken with an inverted microscope (Olympus, Japan) and counted with ImageJ.
Induction of adipogenic and osteogenic differentiation
MSCs were incubated in adipogenic or osteogenic differentiation medium [18]. Adipogenic differentiation potential was detected with the Oil Red O staining kit (Sigma-Aldrich, Shanghai, China) at day 12 of induction, and osteogenic differentiation potential was detected with alkaline phosphatase (Sigma-Aldrich) and the alizarin red (Sigma-Aldrich) staining kit at days 5 and 12 of induction, respectively.
The staining procedures were performed according to manufacturer's instructions.
Flow cytometry analysis
MSCs at passage 3 (P3) and passage 16 (P16) were harvested and washed twice with phosphate-buffered saline (PBS; Servicebio, Wuhan, China). Then, MSCs were incubated with FITC-conjugated antibody, CD206, and phycoerythrin (PE)-conjugated antibodies, CD29, CD44, CD73, CD90, CD105, CD34, CD45, CD106, and HLA-DR, or isotype control antibodies (BD Biosciences, Heidelberg, Germany) at 4°C for 30 min in the dark.
After two washes, the MSC phenotype was analyzed using an Accuri C6 flow cytometer and FlowJo software (BD Biosciences).
Quantitative real-time polymerase chain reaction
Total RNA was extracted by TRIzol (Invitrogen, Shanghai, China) and its concentration and purity were determined by optical density. Then, cDNA was synthesized with the M-MLV reverse transcriptase kit (Takara, Tokyo, Japan). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using SYBR Green (YEASEN, Shanghai, China) and relative mRNA expression was calculated by the 2-ΔΔCt method and normalized to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
All procedures followed the manufacturer's instructions.
Western blotting
Proteins were extracted with radio-immunoprecipitation assay lysis buffer containing phenylmethanesulfonyl fluoride (1:100) and their concentrations were determined with a bicinchoninic acid (BCA) protein assay kit (Beyotime). Western blotting was performed in triplicate according to a previously reported method [19], with GAPDH as an internal control.
Enzyme-linked immunosorbent assay
Culture medium of MSCs at P3 or P16 was collected and centrifuged at 3000 rpm for 20 min at 4°C. Then, the supernatant was gathered and IL-1β, IL-6, IL-8, and MCP-1 were detected by the human enzyme-linked immunosorbent assay (ELISA) kit (Jiangsu Meimian Industrial Co., Ltd., Jiangsu, China). All procedures were performed according to the manufacturer's instructions.
sEV isolation
The normal culture medium was replaced with serum-free Dulbecco's modification of Eagle's medium/F12 medium when MSCs reached 90% confluence. After 48 h of incubation, the conditioned medium was collected and then centrifuged at 3,000 rpm for 20 min to remove cells and debris. Next, the supernatant was filtered through a 0.1-mm-pore membrane filter (Corning, NY, USA) to eliminate large vesicles.
Finally, sEVs were concentrated using CentriPlus-70 ultrafiltration membranes (Millipore, NY, USA) and resuspended in PBS.
Transmission electron microscopy
Purified sEVs were fixed with PBS (pH 7.4) containing 1% glutaraldehyde, and then 20 μL of sEV suspension was loaded onto a carbon-coated copper grid for 10 min at room temperature. Subsequently, they were stained with 2% uranyl acetate for 2 min and visualized under transmission electron microscopy (TEM).
Nanoparticle tracking analysis
Purified sEVs were typically diluted about 100-fold in 1 mL of PBS, and then their size distribution and concentration were recorded by nanoparticle tracking analysis (NTS) using a ZetaView instrument (Particle Metrix).
Cell apoptosis assay
A 5-mM GW4869 or spiroepoxide solution was prepared in dimethyl sulfoxide, or a 10-mM 1-stearoyl-GPC (18:0) solution was prepared in PBS, and then diluted as needed in cell culture medium. MSCs at P3 or P16 were seeded in 12-well culture plates and exposed to different doses of GW4869 or spiroepoxide (1, 2, and 4 μM) for 2 h or treated with different concentrations of 1-stearoyl-GPC (18:0) (20, 40, and 80 μM) for 24 h when MSCs reached 70% confluence.
The percentage of apoptotic cells was examined with the Annexin V-FITC/PI apoptosis detection kit (YEASEN) following the manufacturer's instructions. Apoptosis was analyzed using an Accuri C6 flow cytometer (BD Biosciences).
Cell viability assay
MSCs were seeded in 96-well culture plates and then cocultured with different doses of GW4869 or spiroepoxide (1, 2, and 4 μM) for 2 h. Cell viability was measured using the Cell Counting Kit-8 (YEASEN) following the manufacturer's instructions, and absorbance was measured at 450 nm using a microplate reader (Multiskan FC).
Statistical analysis
All data are presented as mean ± standard deviation. Comparisons between groups were analyzed by Student's t-test, two-way analysis of variance (ANOVA), followed by Šídák's multiple comparisons test, and one-way ANOVA, followed by Dunnett's multiple comparisons test. Statistically significant differences were defined at *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.
Results
Establishment of a replicative senescence model for MSCs
Senescence of MSCs from three different healthy volunteers was induced by serial passage in vitro. When cells were cultured to P16, the percentage of senescence-associated β-galactosidase (SA β-gal) staining-positive cells significantly increased to >80%, with the senescence-related morphological features: flat and irregular appearance (P < 0.0001; Fig. 1A).
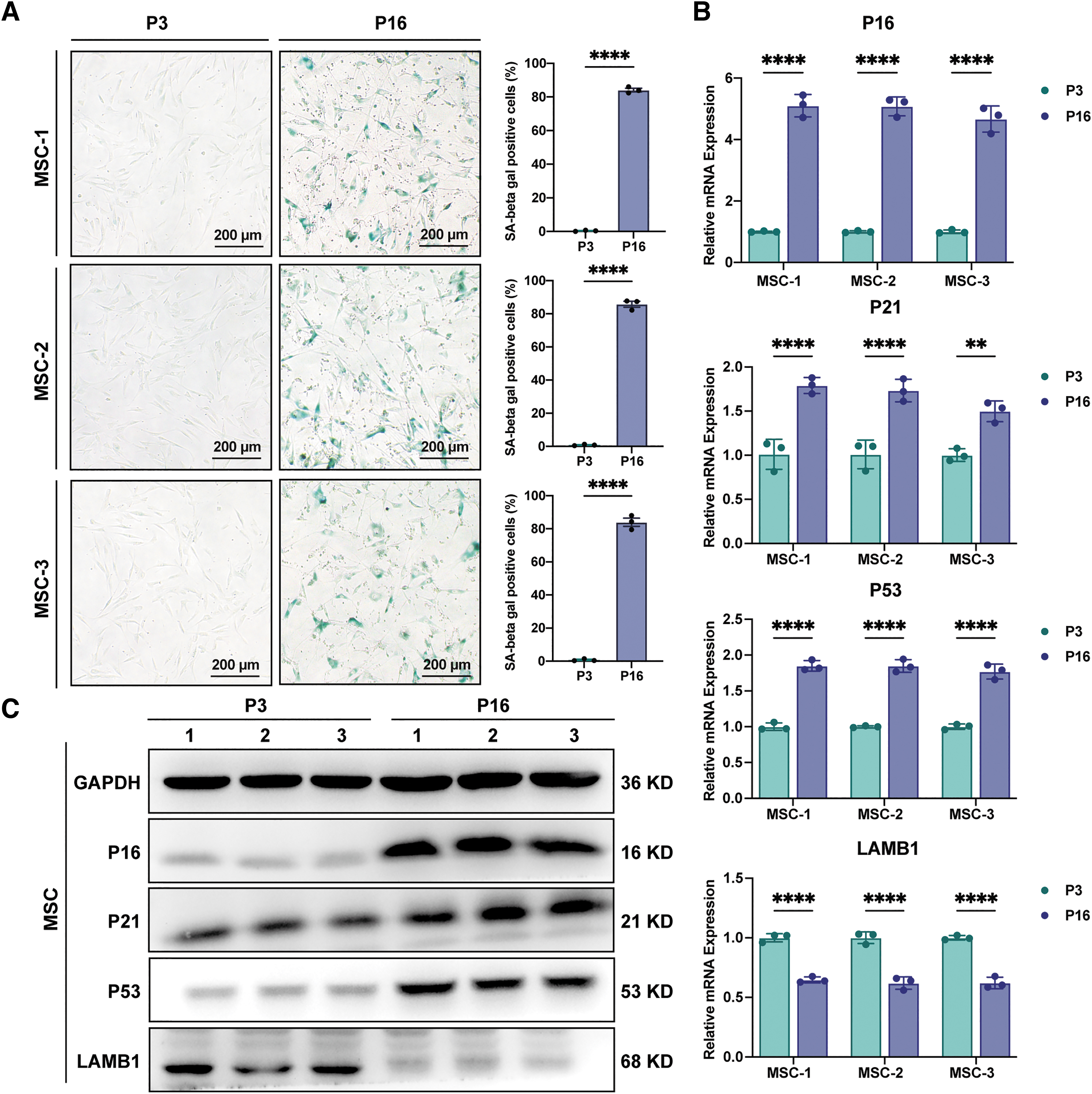
Establishment of a replicative senescence model for MSCs. MSCs 1, 2, and 3 represent MSCs from three different volunteers.
To further verify the senescence of MSCs, we examined the expression of senescence biomarkers. qRT-PCR revealed that the positive senescence markers, P16, P21, and P53, were elevated and the negative nuclear morphology marker, LMNB1, was greatly downregulated in P16 MSCs (P < 0.0001 and P < 0.01; Fig. 1B), which were also confirmed by western blotting (Fig. 1C).
These results indicated that serial passage of MSCs to P16 successfully established the replicative senescence MSC model in vitro, and thereafter, P3 MSCs were noted as presenescent MSCs (Pre-Sen MSCs) and P16 MSCs as Sen MSCs in this article.
Features of Sen MSCs
We then compared the phenotypic changes between Pre-Sen and Sen MSCs. The MSC immunophenotype remained stable from Pre-Sen MSCs to Sen MSCs, expressing CD29, CD44, CD73, CD90, and CD105, but not CD34, CD45, CD106, CD206, and HLA-DR (Fig. 2A). However, the adipogenic and osteogenic differentiation potential was markedly decreased in Sen MSCs compared with Pre-Sen MSCs (Fig. 2B–D).
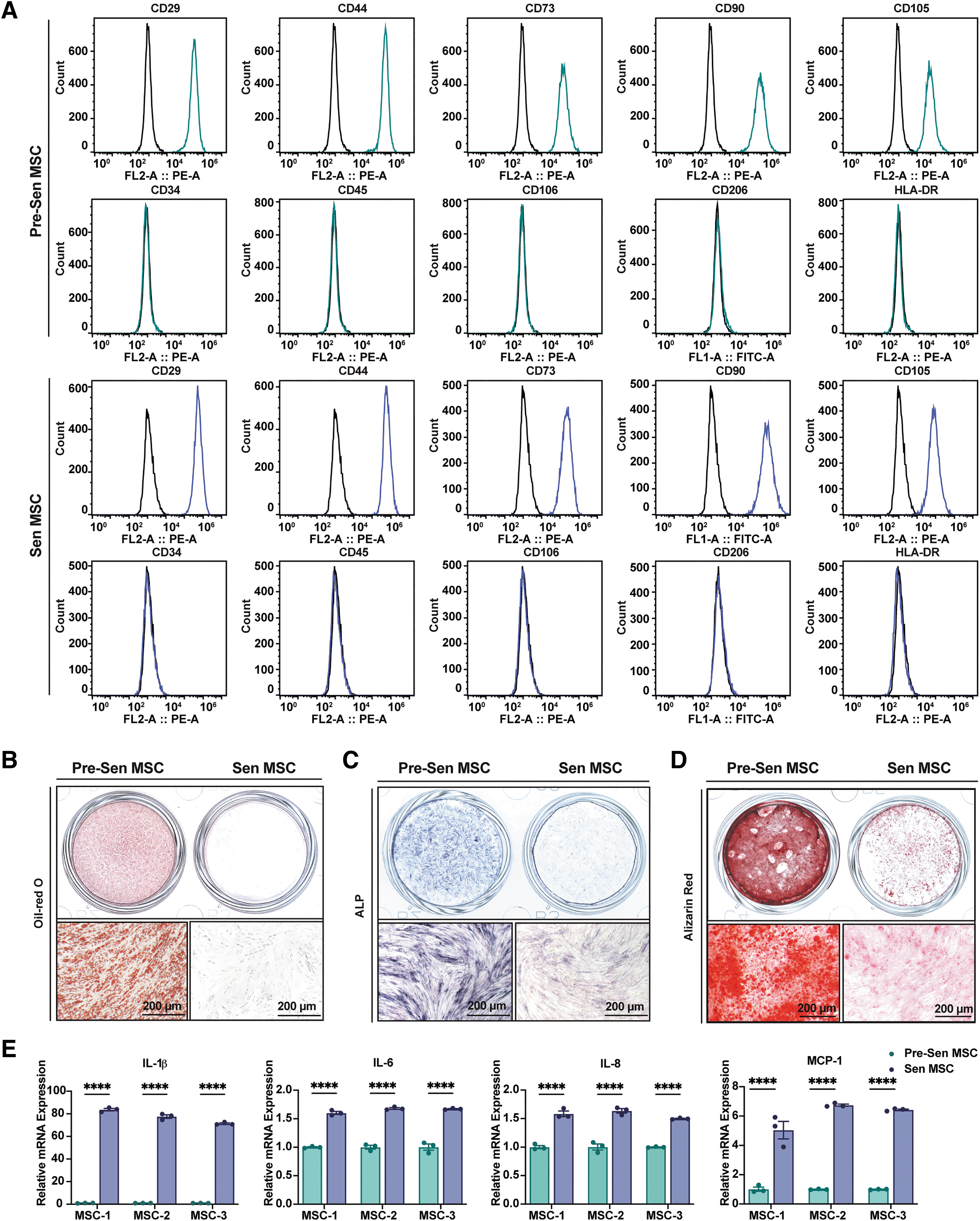
Features of Sen MSCs. MSC-1, 2, and 3 represent MSCs from three different volunteers.
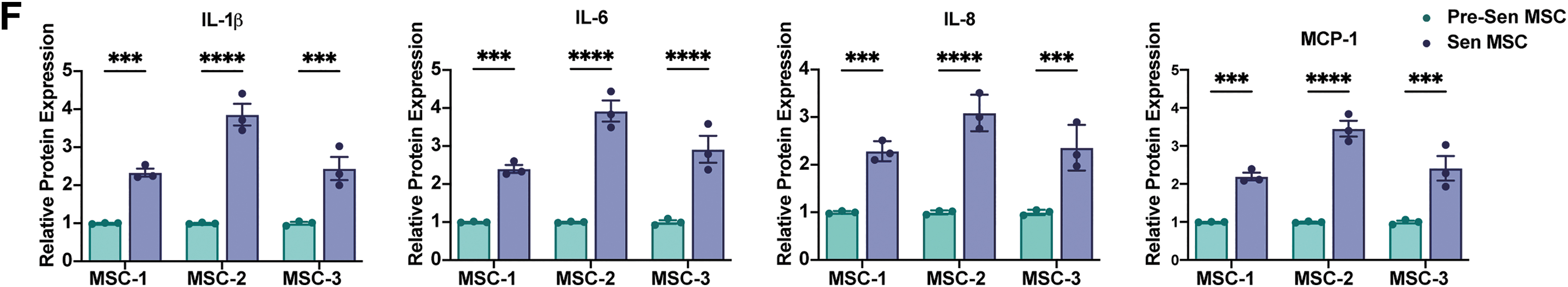
Moreover, proinflammatory cytokines, IL-1β, IL-6, and IL-8, and the chemokine, MCP-1, which are markers of the SASP, were tested by qRT-PCR and ELISA and showed significant upregulation in Sen MSCs at both mRNA (P < 0.0001; Fig. 2E) and protein levels (P < 0.0001 and P < 0.001; Fig. 2F).
These results revealed that Sen MSCs exhibited an unaltered MSC immunophenotype, diminished adipogenic and osteogenic differentiation potential, and elevated levels of SASP factors.
Increased sEV secretion in Sen MSCs
sEVs were isolated from the conditioned medium of Pre-Sen MSCs or Sen MSCs using ultrafiltration and characterized by TEM, western blot, and NTA. Under TEM, both Pre-Sen MSC-derived sEVs (Pre-Sen-sEVs) and Sen MSC-derived sEVs (Sen-sEVs) exhibited typical cup-shaped or sphere-shaped morphology (Fig. 3A).
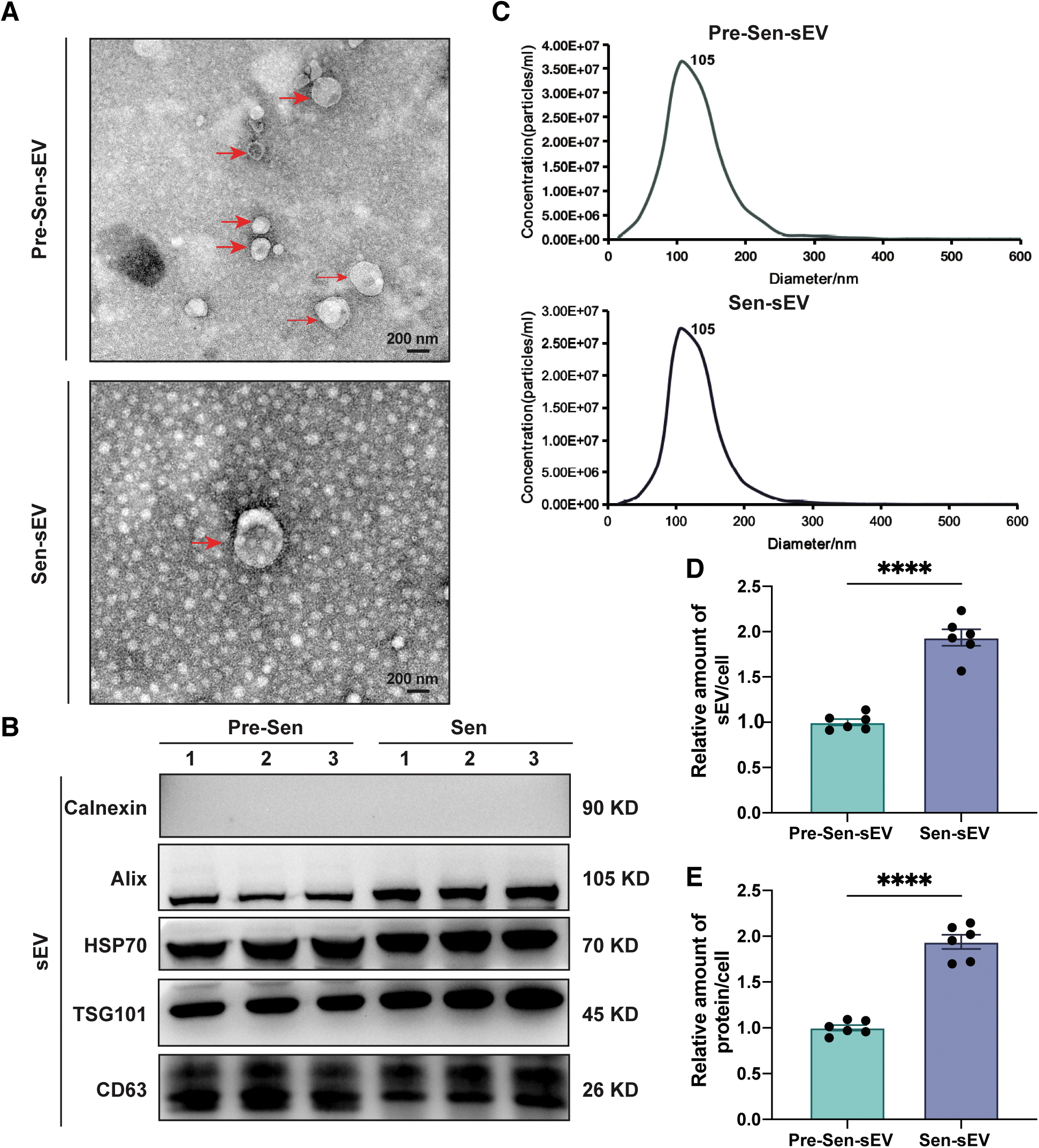
Increased sEV secretion in senescent MSCs. sEVs were obtained from MSCs derived from one volunteer.
Western blotting showed that sEV-associated protein markers, CD63 (transmembrane protein), HSP70 (heat shock protein), Alix, and TSG101 (cytosolic protein relevant to sEV biogenesis), were abundant in both Pre-Sen-sEVs and Sen-sEVs, and the negative marker, calnexin (a protein in endoplasmic reticulum), was not found (Fig. 3B).
According to NTA, the size distribution of Pre-Sen-sEVs and Sen-sEVs was not significantly different between 55 and 200 nm (Fig. 3C), but interestingly, there was an obvious increase in the quantity of Sen-sEVs when compared with Pre-Sen-sEVs obtained from an equivalent number of cells (P < 0.0001; Fig. 3D).
Consistent with this, the BCA assay revealed that the concentration of Sen-sEV lysates was also increased in comparison with Pre-Sen-sEVs derived from the same number of cells (P < 0.0001; Fig. 3E). These results revealed that production of sEVs was increased in Sen MSCs.
Increased sEV secretion maintains senescent MSC homeostasis
Previous studies supported the notion that sEVs may act as cellular garbage bags to expel cellular metabolic waste [20,21], and subsequent research found that sEVs have the potential to eliminate cytotoxic cytoplasmic DNA from cells [22]. Therefore, we speculated that the increased secretion of sEVs from Sen MSCs may be aimed at maintaining cellular homeostasis.
To prove this speculation, GW4869 or spiroepoxide, two well-known inhibitors of sEV production [22,23], were used to inhibit sEV secretion and they led to decreased sEV secretion in Pre-Sen MSCs and Sen MSCs, as judged by NTA (P < 0.01, P < 0.001, and P < 0.0001; Fig. 4A). Inhibition of sEV secretion resulted in increased apoptosis in both Pre-Sen and Sen MSCs.
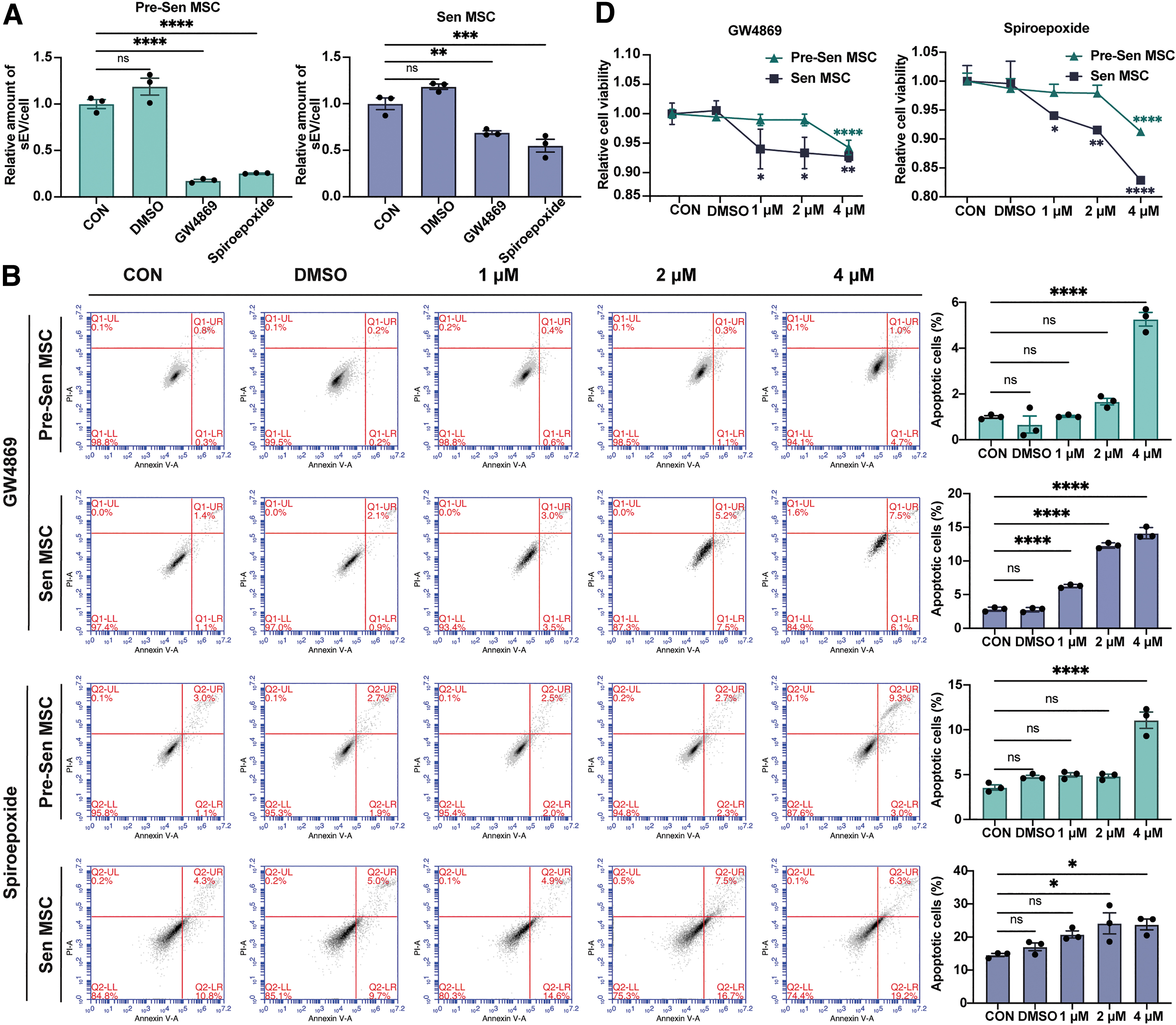
Increased sEV secretion maintains Sen MSC homeostasis. Experiments were repeated on MSCs derived from one volunteer.
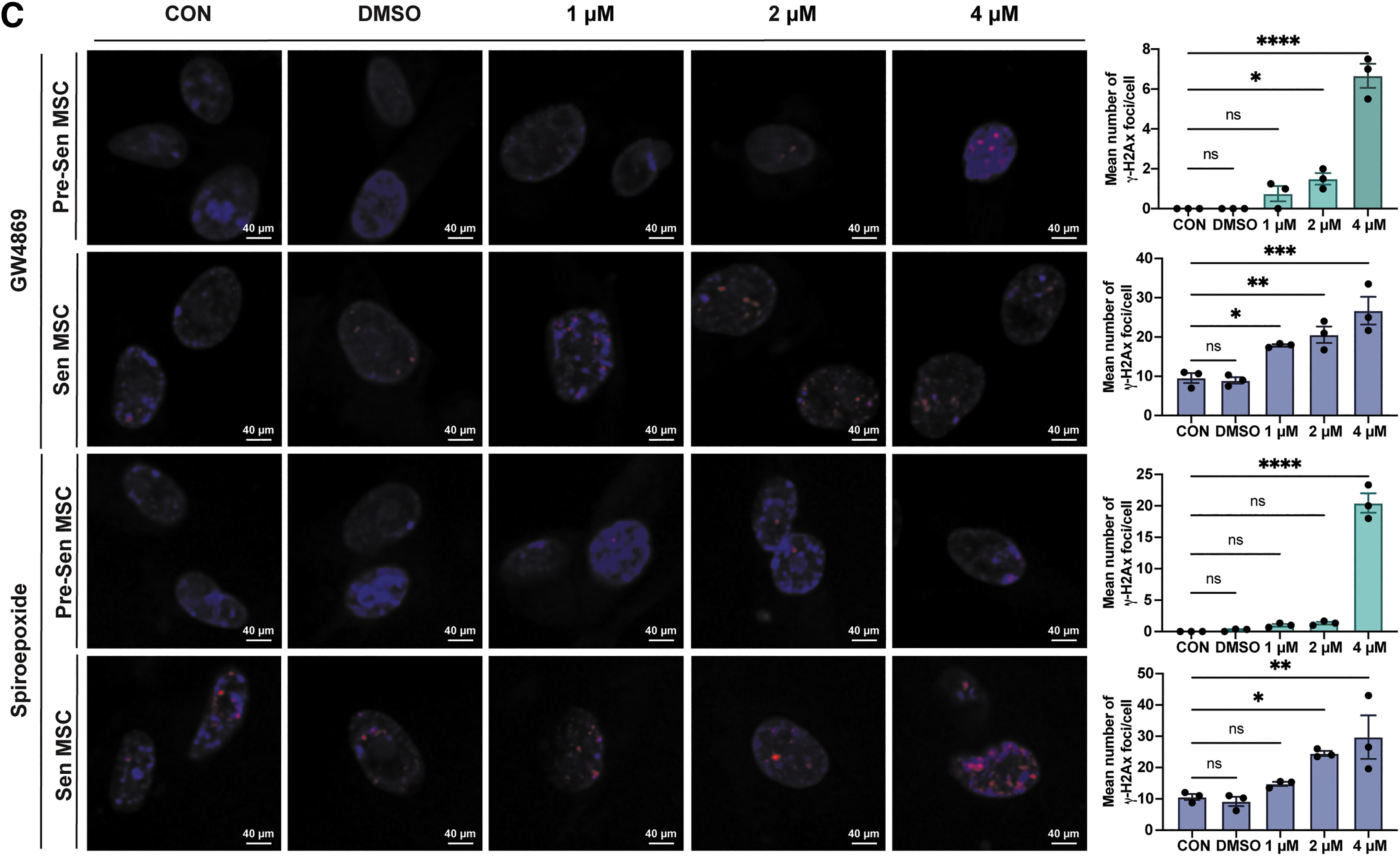
Most notably, Sen MSCs already showed significant apoptosis at a GW4869 concentration of 1 μM in a concentration-dependent manner, but the apoptosis of Pre-Sen MSCs started when GW4869 was elevated to 4 μM, and a similar trend was also observed in inhibition of sEV secretion by spiroepoxide (P < 0.05 and P < 0.0001; Fig. 4B), suggesting that maintenance of Sen MSC homeostasis is more dependent on sEV secretion, which may explain the biological purpose of the increased sEV secretion in Sen MSCs.
Consistently, γ-H2Ax immunofluorescence staining showed that inhibition of sEV secretion was more likely to result in significant DNA damage in Sen MSCs (P < 0.05, P < 0.01, P < 0.001, and P < 0.0001; Fig. 4C). Moreover, the CCK8 assay showed that suppression of sEV secretion resulted in decreased cell viability of Pre-Sen and Sen MSCs, but Sen MSCs were more sensitive to the inhibitor (P < 0.05, P < 0.01, and P < 0.0001; Fig. 4D).
Collectively, these results demonstrated that the increased sEV secretion plays an important role in maintaining cellular homeostasis of Sen MSCs.
Changes of individual metabolites in Pre-Sen-sEVs and Sen-sEVs
To further investigate the molecular mechanisms by which sEVs maintain Sen MSC homeostasis, metabolomic profiling was performed on Pre-Sen-sEVs and Sen-sEVs. A total of 216 metabolites were detected, including 13 unique metabolites in Pre-Sen-sEVs, 4 unique metabolites in Sen-sEVs, and 199 metabolites in both groups (Fig. 5A).
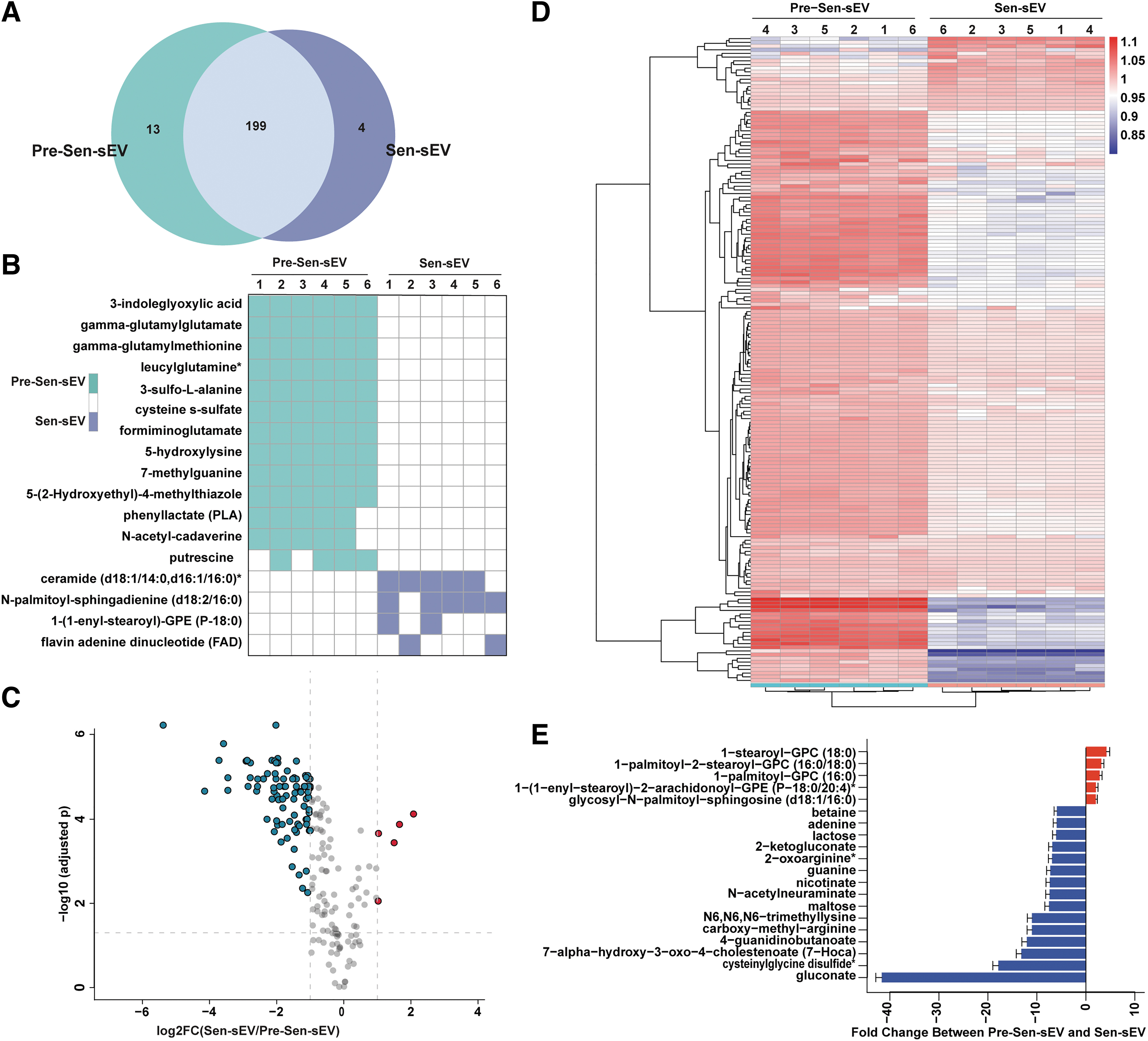
Relative changes of individual metabolites in Pre-Sen-sEVs and Sen-sEVs. sEVs were obtained from MSCs derived from one volunteer.
The distribution of unique metabolites in each sample is presented in Fig. 5B, which reveals that three of the four unique metabolites in Sen-sEVs were lipid metabolites, including ceramide (d18:1/14:0, d16:1/16:0)*, N-palmitoyl-sphingadienine (d18:2/16:0)*, and 1-(1-enyl-stearoyl)-GPE (P-18:0)* (Fig. 5B). Significant differences were observed in 182 metabolites, of which 26 metabolites were upregulated and 156 metabolites were downregulated in Sen-sEVs (Fig. 5C).
A circular heat map provided the intuitive visualization of concentration trends across groups for all differential metabolites (Fig. 5D), with special attention to the top 5 upregulated and top 15 downregulated metabolites in Sen-sEVs (Fig. 5E), and the top 5 metabolites significantly upregulated in Sen-sEVs were all lipid metabolites.
These results tentatively suggested that the unique and significantly upregulated metabolites in Sen-sEVs were mainly lipid metabolites.
Classification and enriched metabolic pathways of differential metabolites
To further comprehensively demonstrate this distinction between Pre-Sen-sEVs and Sen-sEVs, differential metabolites were classified, and the top 5 differential metabolites of each type are listed in box plots (Fig. 6A). A bubble map showed that (as previously expected) the metabolites significantly upregulated in Sen-sEVs were indeed mainly lipid metabolites, including ceramides, lysophosphatidylcholines, phosphatidylcholines, and sphingomyelins (Fig. 6B), which have been shown to be toxic for inducing cellular senescence and apoptosis.
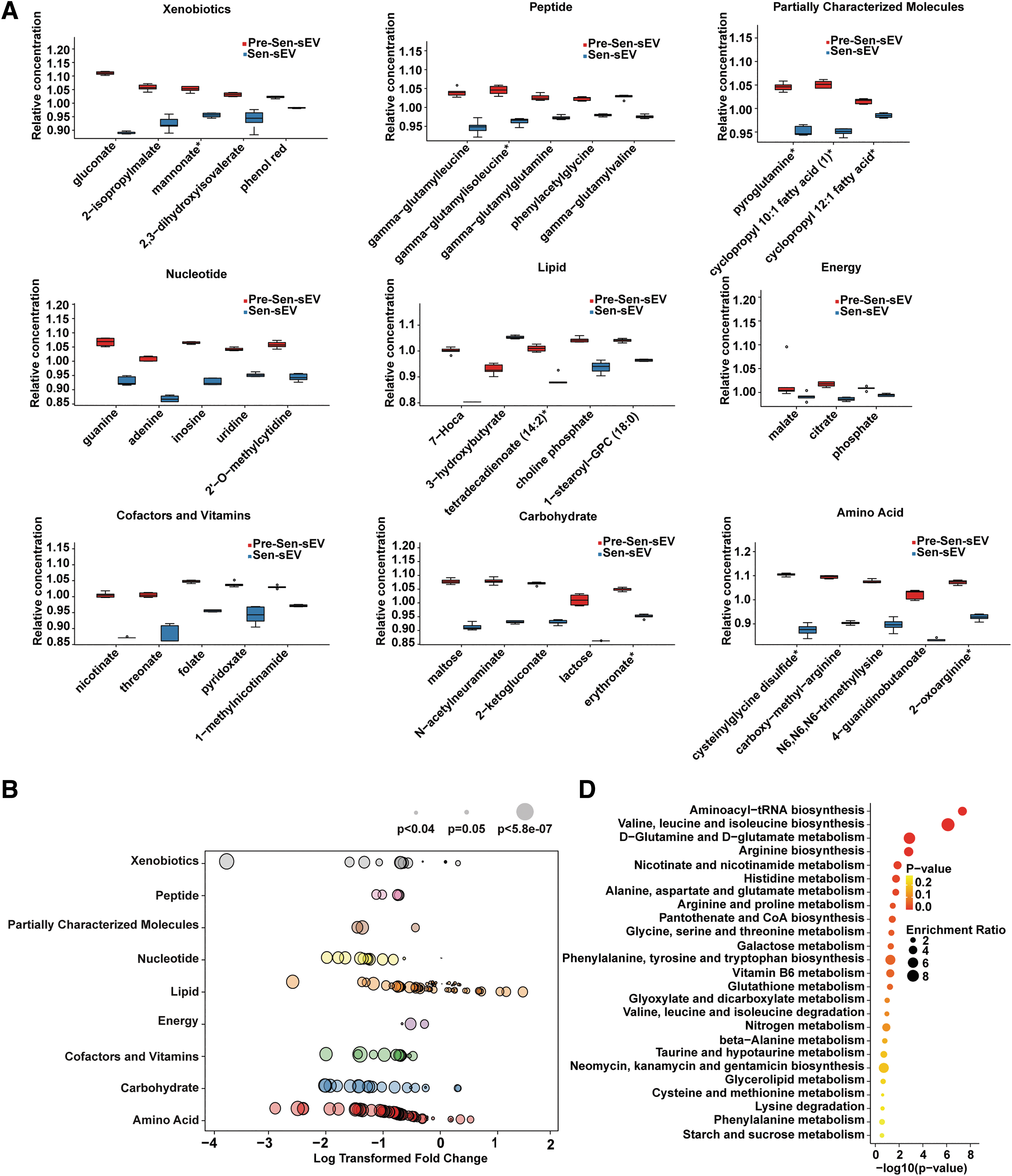
Classification and enriched metabolic pathways of differential metabolites. Experiments were repeated on MSCs derived from one volunteer.
Ceramides accumulate in replicative and stress-induced senescent cells [24 –26] and could induce cellular senescence at low concentrations by activating protein phosphatases (PP1 and PP2A) and promoting P21 expression to inhibit cell cycle-dependent kinase 2 activity [27]. High concentrations of ceramides led to apoptosis, likely through disruption of mitochondrial dynamics and release of cytochrome C [28]. Sphingomyelins could be degraded to ceramides by sphingomyelinase [29], and neutral sphingomyelinase activity was found to be significantly increased in senescent cells [30].
Senescent fibroblasts were enriched in lysophosphatidylcholines [31] that are involved in the regulation of oxidative stress and inflammation [32,33]. Low levels of lysophosphatidylcholines induced upregulation of SA β-gal and p21 expression and increased secretion of SASP in bile duct cells [34]. High levels of lysophosphatidylcholines triggered apoptosis due to endoplasmic reticulum stress and mitochondrial integrity damage in hepatocytes [35 –37].
Phosphatidylcholines could be cleaved to lysophosphatidylcholines by phospholipase A2, and dysregulation of phosphatidylcholines may lead to disruption of lysophosphatidylcholine levels [38,39]. These bioactive lipids were released by the cell either as free molecules or as part of the extracellular vesicle (EV) [32]. Coincidentally, recent studies have found that DNA damage-induced senescent fibroblasts enhance EV biogenesis by upregulating ceramide synthesis, which could protect cells from inflammatory damage [40].
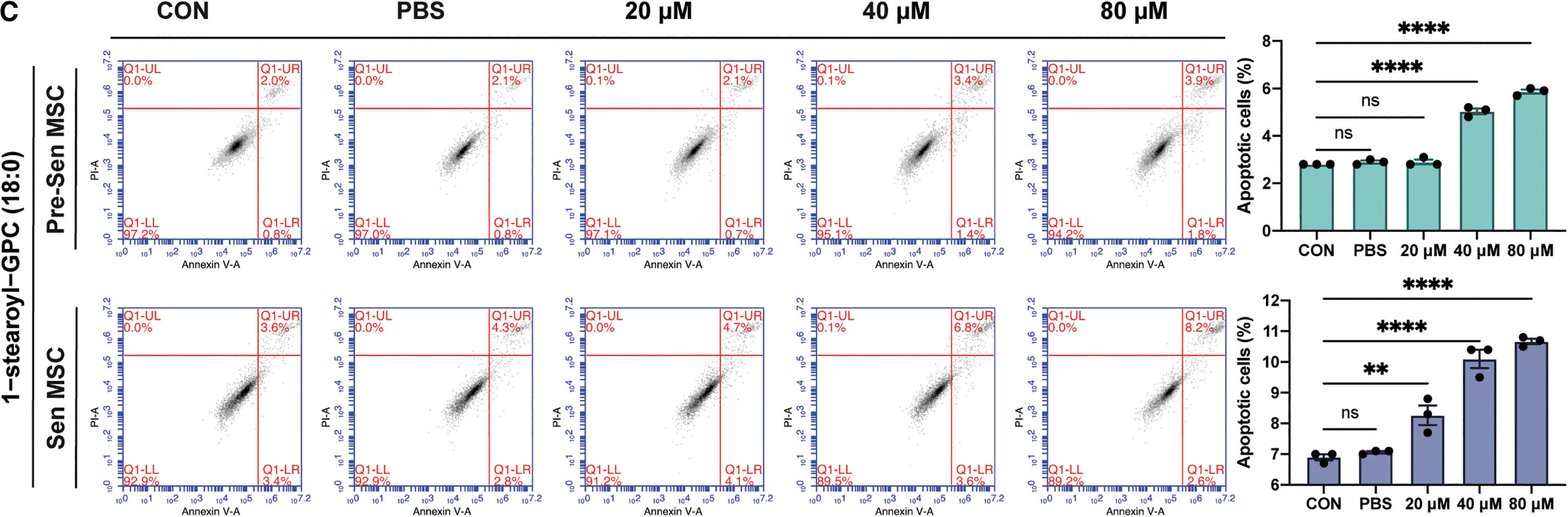
H-Ras-induced fibroblast senescence resulted in increased EV secretion and marked changes in lipid composition of EVs, such as enrichment of sphingomyelins and lysophosphatidylcholines in EVs [41]. 1-Stearoyl-GPC (18:0) was the most significantly elevated lipid metabolite in Sen-sEVs (Fig. 6A). Treatment of MSCs with different concentrations of 1-stearoyl-GPC (18:0) resulted in increased apoptosis in Pre-Sen and Sen MSCs (P < 0.01 and P < 0.0001; Fig. 6C), indirectly suggesting that some lipids upregulated in Sen-sEVs have apoptosis-inducing toxicity and that sEVs may protect MSCs from apoptosis by excreting harmful lipids.
To further explore the signaling pathways involved in the differential metabolites, KEGG analysis was performed and results showed enrichment of these differential metabolites mainly in 25 signaling pathways (Fig. 6C), of which 21 metabolic pathways have been shown to be closely associated with senescence. Aminoacyl-tRNA biosynthesis decreased with age [42,43], and alanine, aspartate, and glutamate metabolism was impaired in aging rat brain cortex cells [44].
The reduction of glycine, serine, and threonine metabolism in dental pulp stem cells resulted in increased P16 expression and cellular senescence [45]. Increasing vitamin B6 metabolism promoted 5-hydroxytryptamine production, which was a valuable therapeutic target for reversing aging-related degeneration in the liver [46]. Fibroblast-derived extracellular vesicles enriched with glutathione metabolism pathway-related proteins ameliorated the aging-related phenotype of aged mouse tissues [47].
In addition, a series of metabolomic analyses based on senescence showed that valine, leucine, and isoleucine biosynthesis [48]; d-glutamine and d-glutamate metabolism [49]; arginine biosynthesis [50]; histidine metabolism [49]; phenylalanine, tyrosine, and tryptophan biosynthesis [51]; glyoxylate and dicarboxylate metabolism [49]; valine, leucine, and isoleucine degradation [48]; and cysteine and methionine metabolism were all closely associated with senescence [52].
Various studies on improvement of aging through therapy have shown that the following metabolic pathways were involved in regulation of aging, including nicotinate and nicotinamide metabolism [53], arginine and proline metabolism [54], nitrogen metabolism [55], taurine and hypotaurine metabolism [56], glycerolipid metabolism [57], lysine degradation [58], phenylalanine metabolism [59], and starch and sucrose metabolism [60].
Therefore, our findings suggested that sEVs may maintain Sen MSC homeostasis by regulating complex senescence-related pathways. Taken together, these results demonstrate that the increased sEV secretion plays a vital role in maintaining Sen MSC homeostasis, at least in part, through excretion of harmful lipids.
Discussion
Cell metabolism inevitably produces reactive oxygen species (ROS), and excess ROS leads to DNA damage, protein damage, mitochondrial dysfunction, autophagy inhibition, telomere loss [61 –63], and eventually cellular senescence [64]. To protect cells from oxidative stress damage, ROS are reduced by glutathione [65], a process that has recently been found to occur mainly on the cytoplasmic surfaces of intracellular vesicles [66].
Interestingly, many intracellular vesicles are generated during the biogenesis of sEVs, such as early-sorting endosomes, late-sorting endosomes, and multivesicular endosomes [8]. This intrinsic link between reduction of senescence-related damage and sEV biogenesis further exemplifies the important role of sEVs in maintaining cellular homeostasis.
MSC-derived sEVs are promising, cell-free therapeutic agents for a variety of diseases. To obtain sufficient sEVs, MSCs need to be extensively expanded, which poses the risk of senescence. sEVs are cellular secretions whose content is functionally dependent on the state of the parent cell, so the quality of sEVs needs to be tightly controlled. However, there is no exact indicator to define whether sEVs are affected by the senescence of parent cells. The metabolites identified by metabolomic profiling in this study, which are unique or significantly different between Pre-Sen-sEVs and Sen-sEVs, may provide candidate markers for identifying the eligibility of senescent sEVs.
In addition, sEVs have the potential to assist in disease diagnosis [67]. sEVs can be obtained from all biological fluids and are more convenient to sample compared with tissue biopsies [68]. sEVs contain complex cellular components that provide a more complete picture of the physiological state of cells [68]. In addition, the sEV-based histological analysis provides a solid theoretical basis for screening diagnosis markers [69 –71]. MSC senescence is closely related to many aging-related diseases.
Decreased osteogenic capacity and increased secretion of SASP in senescent MSCs play an important role in the pathogenesis of osteoporosis and osteoarthritis [72]. In Hutchinson–Gilford progeria syndrome, MSC senescence is one of the major pathological changes [73], and upregulation of MSC senescence-related genes predicts poor prognosis in multiple myeloma [74]. The significant difference between Pre-Sen-sEVs and Sen-sEVs found in this study may provide new directions for the diagnosis of aging-related diseases.
Conclusions
This study suggested that increased sEV secretion maintains Sen MSC homeostasis, at least in part, by excreting deleterious lipids, which provides new insights into the regulation of senescence by sEVs and enhances our understanding of maintenance of cellular homeostasis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (Grant No. 81902848), CAMS Innovation Fund for Medical Sciences (Grant No. 2022-I2M-1-012, 2021-I2M-1-051), Key Area Research and Development Program of Guangdong Province (Grant No. 2021B0909060001), and 2021 General Research Institute for Nonferrous Metals “Unveiling and Commanding” Project (Grant No. 2021JC0103).