
Abstract
Seipin deficiency is an important cause of type 2 Berardinelli-Seip congenital dyslipidemia (BSCL2). BSCL2 is a severe lipodystrophy syndrome with lack of adipose tissue, hepatic steatosis, insulin resistance, and normal or higher bone mineral density. Bone marrow mesenchymal stem cells (BMSCs) are believed to maintain bone and fat homeostasis by differentiating into osteoblasts and adipocytes. We aimed to explore the role of seipin in the osteogenic/adipogenic differentiation balance of BMSCs. Seipin loxP/loxP mice are used to explore metabolic disorders caused by seipin gene mutations. Compared with wild-type mice, subcutaneous fat deficiency and ectopic fat accumulation were higher in seipin knockout mice. Microcomputed tomography of the tibia revealed the increased bone content in seipin knockout mice. We generated seipin-deficient BMSCs in vitro and revealed that lipogenic genes are downregulated and osteogenic genes are upregulated in seipin-deficient BMSCs. In addition, peroxisome proliferator-activated receptor gamma (PPARγ) signaling is reduced in seipin-deficient BMSCs, while using the PPARγ activator increased the lipogenic differentiation and decreased osteogenic differentiation of seipin-deficient BMSCs. Our findings indicated that bone and lipid metabolism can be regulated by seipin through modulating the differentiation of mesenchymal stem cells. Thus, a new insight of seipin mutations in lipid metabolism disorders was revealed, providing a prospective strategy for MSC transplantation-based treatment of BSCL2.
Introduction
Berardinelli-Seip congenital dyslipidemia (BSCL), a disease characterized by complete absence of adipose tissue and metabolic disorders, is also known as congenital generalized lipodystrophy (CGL) [1]. CGL includes four genetic variant types of genes: 1-acylglycerol-3-phosphate O-acyltransferase (AGPAT2), type 2 Berardinelli-Seip congenital lipodystrophy (BSCL2), caveolin (CAV1), and caveola-associated protein (CAVIN) [2]. Clinically, BCLC2 is one of the most common subtypes of CGL.
BSCL2 is caused by mutations in the seipin gene. Patients with BSCL2 showed severe lipodystrophy, insulin resistance, heterotopic lipid deposition of muscle, and hepatic steatosis [3,4]. Seipin plays a vital part in adipogenesis, lipid droplet (LD) homeostasis, and cellular triglyceride lipolysis [5,6]. As a regulator of adipogenic differentiation, seipin promotes normal lipogenesis, and deficiency of seipin can lead to abnormal LD formation.
Seipin–/– mice were shown to exhibit less adipose tissue, insulin resistance, and hepatic steatosis compared with wild-type mice (WT mice) [7]. Thus, the seipin–/– mouse model is considered as strong evidence to investigate the function of seipin and development of BSCL2.
In the process of adipocyte differentiation, seipin is required to regulate the morphology of LDs and development of adipocytes by activating glycerol-3-phosphate acyltransferase [8]. Seipin could form a complex with promethin to participate in the function and identity of LDs [9]. Moreover, the seipin mutation was found to play an important role in maturation of adipocytes from mesenchymal stem cells (MSCs) [10].
As multipotent stem cells, MSCs can differentiate into osteoblasts, chondroblasts, and adipocytes. MSCs maintain a mutually exclusive balance in adipogenesis and osteogenesis, with factors that inhibit osteogenesis, promoting lipogenic processes [11]. Peroxisome proliferator-activated receptor gamma (PPARγ) can promote the differentiation of MSCs to adipocytes and also inhibit their differentiation to osteoblasts [12]. Knockdown of seipin reduces the differentiation of adipocytes and this reduction is rescued by the PPARγ activator, pioglitazone [13].
Therefore, we explored the link between seipin and mutation-induced lipodystrophy by the establishment of a seipin–/– mouse model. We generated seipin-deficient bone marrow mesenchymal stem cells (BMSCs) in vitro and revealed the critical role of seipin in BMSCs by activating the PPARγ signaling pathway.
This investigation will help us to better understand the relationship between seipin and fat metabolism and bone metabolism. With the development of tissue engineering and regenerative medicine, our data may provide evidence for MSCs in the treatment of lipid metabolism disturbance (Fig. 1).
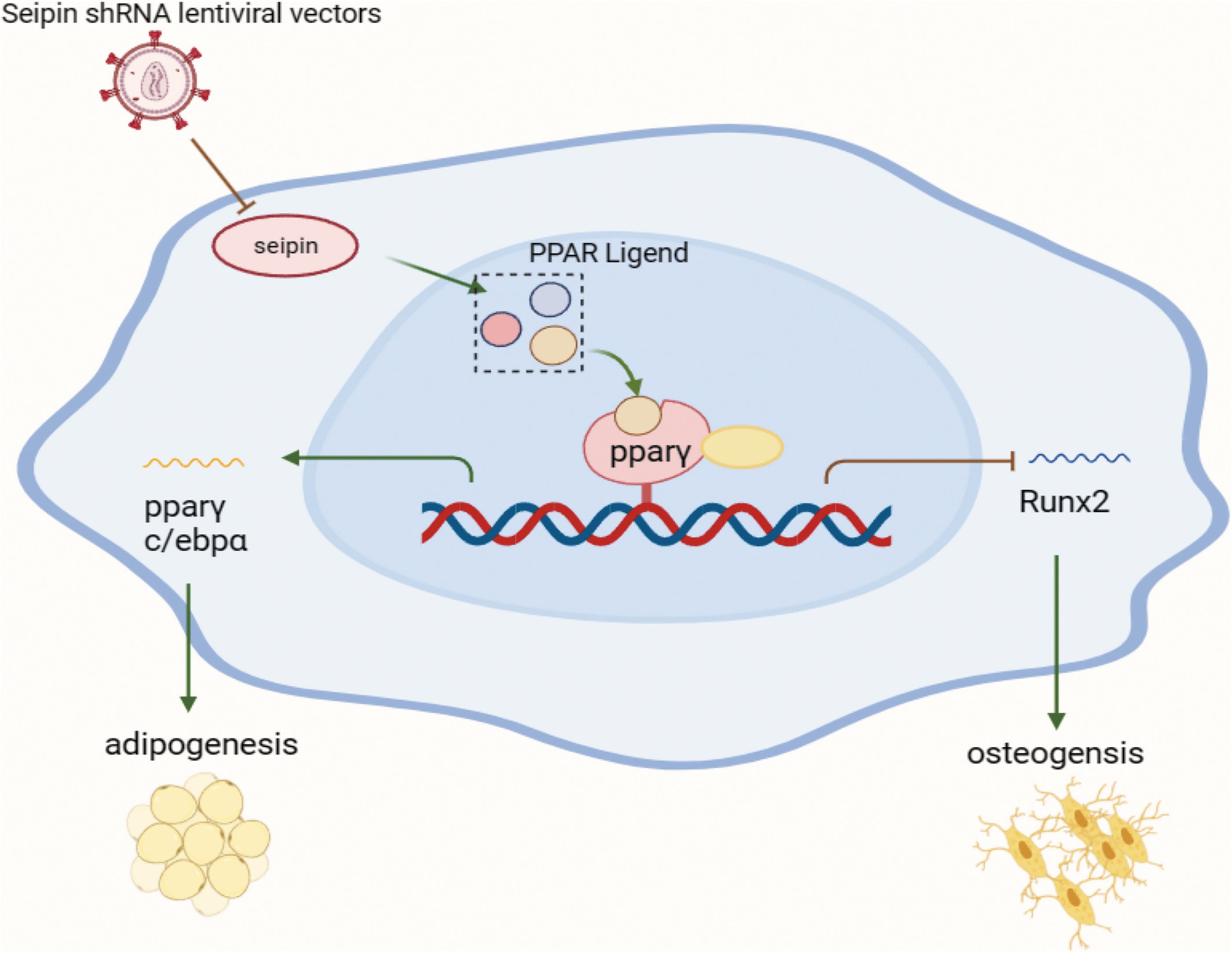
Schematic illustration. Regulation of osteogenic and adipogenic differentiation in BMSCs by seipin. The reduced ligand production of PPARγ after Seipin knockdown inhibited PPAR activity, promoting the expression of the osteogenic differentiation-associated factor runx2 and facilitating osteogenic differentiation of BMSCs; inhibition of the expression of the lipogenic differentiation-associated factor (PPARγ, C/EBPα) inhibited adipogenic differentiation of BMSCs. BMSCs, bone marrow mesenchymal stem cells; PPARγ, peroxisome proliferator-activated receptor gamma; EBPα, enhancer-binding protein alpha.
Materials and Methods
Experimental animals
All animal handling procedures in this study were approved by the Research Ethics Committee of the First Affiliated Hospital, School of Medicine, Shihezi University, and conformed to the National Institutes of Health Guide for the care and use of laboratory animals. Seipin global knockout mice (C57BL/6) were acquired from the laboratory of Peking University Health Science Center.
Seipin knockout homozygous mice (SKO; Seipin140 –/–) and WT (Seipin +/+) mice were produced by breeding of heterozygous mice (heterozygote; HET; Seipin +/–). Mice were fed in accordance with the SPF level condition and allowed access to standard food. They were also fed normally and kept at a temperature of 19°C–25°C for 12-day–12-night periods before and after the experiments.
Genotype identification
Seipin loxP/loxP mice were produced as depicted (17). Nonadipose-specific deletion of seipin exon 3 was induced by crossing mice with the loxP seipin allele with transgenic mice expressing Cre recombinase driven by a promoter. The specific DNA was amplified with the following primers: loxp-1 (5′-CTTGTCTCAAAGGGGTCT-3′), loxp-Ko (5′-TCAACAGAACAGACGCT-3′), PNRI3-F (5′-TCTATGGCTCCT TCTACTACTC-3′), and PNRI3-R (5′-CGAATGATATGACGACGACT-3′).
The reaction conditions include 30 s of denaturation, 30 s of annealing, and 10 min of elongation at temperatures of 95°C, 56°C, and 72°C, respectively. The sizes of amplified products of PCR were 1,100 and 300 bp, specific for blank and wild allele.
In vivo microcomputed tomography imaging
A microcomputed tomography (micro-CT) X-ray scanner (SkyScan1176u X-ray microtomography system, Belgium) was used for noninvasive determination of the bone mass of male rats at 30 and 60 days. After mice were successfully anesthetized with 1.25% tribromoethanol (0.2 mL/10 g; Jitianbio, Beijing, China), they were fixed in a micro-CT bubble, keeping one of their legs unbent and keeping the tibia parallel to the X-ray long axis.
The following operation was performed by a professional according to instructions on the machine. Proximal tibiae were imaged in a micro-CT scanner within saline. The CT scanner was calibrated weekly for the mineral equivalent value and monthly for determining in-plane spatial resolution. The selected images were compiled into a three-dimensional (3D) image using NRecon software (V1.6.5.2; SkyScan, Belgium). Information of the proximal tibia bone mineral density (BMD) and trabecular thickness (
Isolation and culture of bone MSCs
Bone MSCs were isolated from the femur and tibia of 3- or 4-week-old newborn WT and SKO male mice. Femur and tibia were successfully obtained under anesthesia and aseptic conditions. After cutting off the metaphysis of the tibia and femur, the bone marrow cavity was flushed with a 16-gauge syringe needle (Saint Dragon, Ningbo, China).
The bone marrow tissue was flushed into 60 × 10-mm cell culture dishes (Corning, Shanghai, China) and centrifuged at 400 g for 6 min. The supernatant was then discarded and the precipitate was resuspended with complete medium: Dulbecco's modified eagle medium (DMEM) (Gibco) +10% fetal bovine serum (BI, Israel) +1% penicillin–streptomycin (Solarbio, Beijing, China).
Cell number counting was carried out using the Cedex XS cell count analyzer (Roche, Switzerland) after the supernatant was removed. Cells were then resuspended and seeded at a density of 107 cells in 60 × 10-mm cell culture dishes. Subsequent experiments were put into effect utilizing BMSCs between the second and third passages.
Flow cytometry
Phenotypic identification of BMSCs was performed by flow cytometry. Single-cell suspensions were prepared by taking BMSCs at passage 3 and adjusting the cell density to 1 × 106/mL. CD29 (Abcam), CD45 (Abcam), and CD11b (Abcam) antibodies were added and incubated for 30 min at room temperature, taking care to avoid light. Cells were washed with phosphate-buffered saline (PBS), resuspended in PBS, and then detected by flow cytometry.
Colony-forming unit assays
The colony-forming efficiency was analyzed by colony-forming unit assay. BMSCs were seeded at a density of 1,000 cells/well in a 35 × 10-mm cell culture dish covered with complete medium. After 14 days, the cells were washed using PBS buffer three times. BMSCs were fixed with 4% paraformaldehyde (Solarbio).
Cells were stained with 1 mL of 0.1% crystal violet solution (Beyotime Biotechnology, Shanghai, China). The number of clones was observed using an inverted microscope (OLYMPUS, Japan). Each dish was evaluated in triplicate. The colonies were defined as >50 cells/colony.
Adipogenic and osteogenic differentiation of BMSCs
BMSCs were purified using the whole bone marrow adherent method and cultivated in DMEM low-glucose culture solution with the adipogenic or osteogenic differentiation kit (Cyagen Biosciences, Inc., Guangzhou, China). These induced differentiation media were changed every 3 or 4 days. After 21 days of osteogenic and adipogenic induction, cells were, respectively, stained with Alizarin Red and Oil Red O. The results were observed under the inverted microscope (OLYMPUS).
Oil Red O staining and Alizarin Red staining
The culture plates were fixed with 4% paraformaldehyde for 30 min after adipogenic and osteogenic induction. Mineral deposition of osteoblasts was evaluated by staining with Alizarin Red S (GenMed Scientifics, Inc.). Oil Red O staining (Sigma) was conducted to detect LDs of adipocytes. The images were obtained by employing an inverted microscope (OLYMPUS). Three different images were taken to quantify the number of LDs and bone nodules.
Lentivirus-infected cells
BMSCs grown in the logarithmic phase were inoculated into 12-well plates and cultured in a 5% CO2 incubator at 37°C. Lentivirus infection (GenePharma, China) was carried out when the cell growth density reached 40%–60%. After 24 h, the medium was replaced with fresh medium containing 10% serum. Seventy-two hours after the cells were infected with lentivirus, fluorescence of cells was observed under the fluorescence microscope.
The sequences of oligonucleotides were as follows: small hairpin RNA (shRNA)-seipin#1: 5′-TTAGTCACTGTTTCATGTTAT-3′; shRNA-seipin#2: 5′-CATGCAGAGACCAGATCAAAG-3′; shRNA-seipin#3: 5′-TAGAACTCTACTCTGACTATA-3′; and NC-siRNA, 5′-TTCTCCGAACGTGTCACGT-3′. The effectiveness of seipin deficiency was validated by Western blot analyses.
Western blot
BMSCs were lysed on ice with RIPA lysis solution (Solarbio Life Sciences, Beijing, China). The concentration of total protein was determined using a bicinchoninic acid Protein Assay kit (Biosharp, China). Proteins were separated through 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis at 120 V for 3 h. The proteins were electroblotted onto a polyvinylidene difluoride membrane (Millipore).
After the polyvinylidene fluoride membranes were blocked by bovine serum albumin (Solarbio Life Sciences), they were stained with primary antibodies overnight at 4°C. After 2 h of incubation with the appropriate horseradish peroxidase conjugate secondary antibody, proteins were detected using a ChemiDoc XRS imaging system and Quantity One (Bio-Rad) analysis software.
RNA extraction and quantitative real-time PCR
We used the TRIzol reagent (Life Technologies) to obtain total RNA from BMSCs. Two micrograms of RNA was applied as the template in a 20-μL reaction system equipped with the HiFiScript cDNA synthesis kit (CWBIO, Beijing, China), according to the instruction manual. cDNA was synthesized by incubation at 42°C for 15 min and 85°C for 5 min.
Quantitative real-time PCR was conducted on the 7500 Fast Real-Time PCR System (Life Technologies, Shanghai, China) using the QuantiFast SYBR Green PCR kit (CWBIO), according to manufacturer's instructions.
Statistical analyses
All data are presented as mean ± standard deviation (standard error of mean). The statistical analyses of data were performed with SPSS 26.0. Differences between groups were analyzed for statistical significance using Student's t-tests or two-way analyses of variance, followed by post hoc Bonferroni's test for multiple comparisons. All statistical tests were two-sided and differences were considered to be significant at P < 0.05.
Results
Genotype analysis of the seipin knockout mice
The Cre/loxP recombinant system was used to eliminate seipin and provide a basis for exploring animal models of clinical diseases related to mutations of seipin. The genotype was detected by PCR using genomic DNA. The seipin–/– mice (SKO mice) were identified with a single PCR product at 1,100 bp. Both the 1,100 and 300-bp PCR products were detected in seipin +/– heterozygous mice (HET mice). Seipin +/+ mice (WT mice) were screened with a single PCR product at 300 bp (Supplementary Fig. S1).
Significant lack of adipose tissue and ectopic adipose formation in SKO mice
Eight-week-old mice were selected to investigate whether seipin affected the development of total fat. Compared with WT mice, SKO mice showed significant absence of dorsal subcutaneous adipose tissue (Supplementary Fig. S2A). The livers of 8-week-old SKO mice were dramatically enlarged and abnormally pale, indicating massive fat deposition (Supplementary Fig. S2B).
Increased bone content of SKO mice
The micro-CT findings, including the BMD and 3D trabecular structure, of the tibia are shown in Fig. 2. The results of micro-CT showed that BMD in the 30- and 60-day-old SKO groups was significantly higher than in WT mice (Fig. 2A–F). Based on micro-CT images, 3D models of tibia were created. The
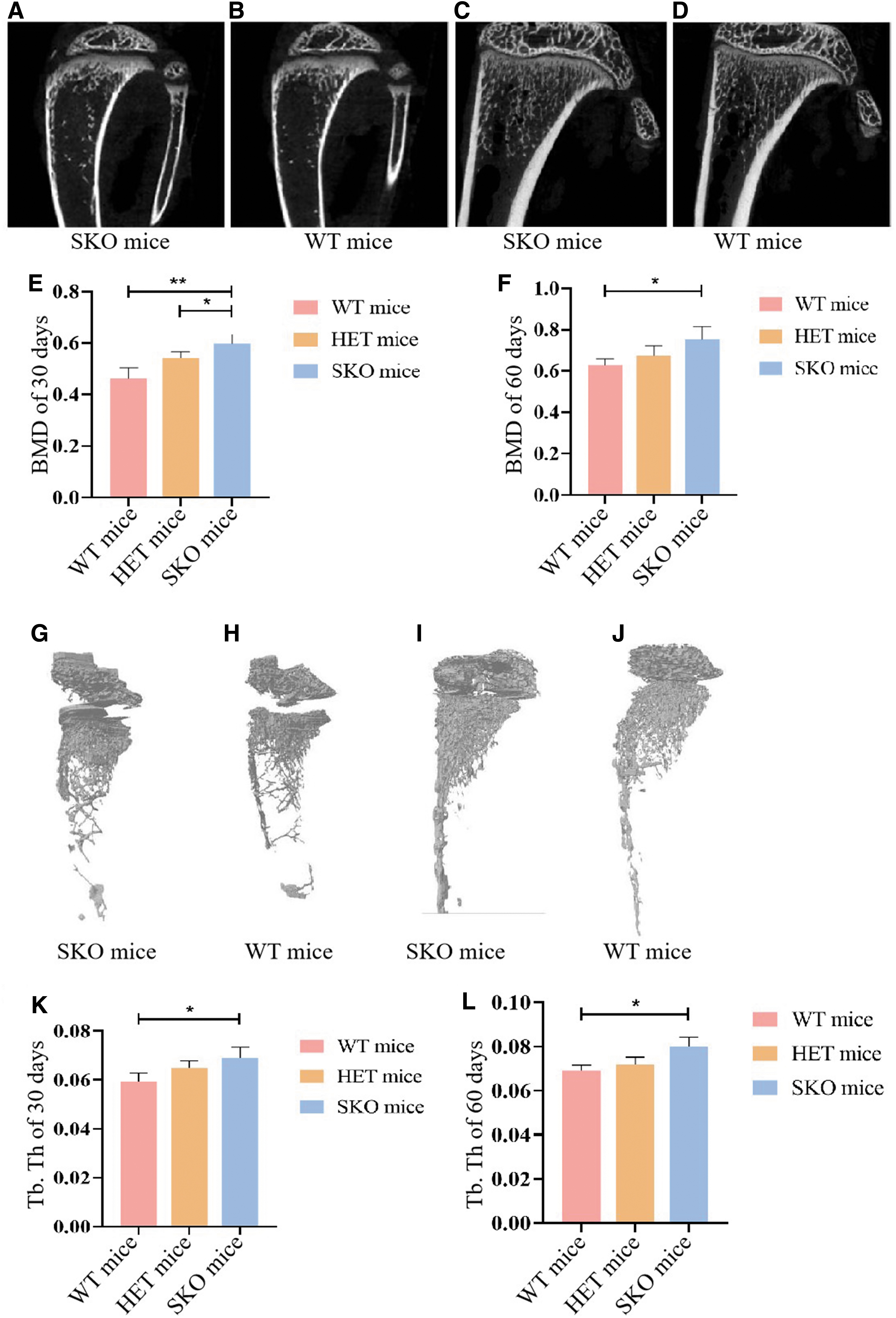
Representative micro-CT images of tibia
Effect of seipin on osteogenic and adipogenic differentiation of BMSCs
We isolated and expanded BMSCs from SKO mice and WT mice. After 3 days of cell inoculation, cell apposition was obvious, and the cells were observed under the microscope to have a polygonal or long, spindle-shaped, fibroblast-like cell morphology with a vortex distribution (Fig. 3A, B). Flow cytometry results showed that P3-generation BMSCs positively expressed CD29 and Sca-1. BMSCs negatively expressed CD11b and CD45 (Fig. 3C, D).
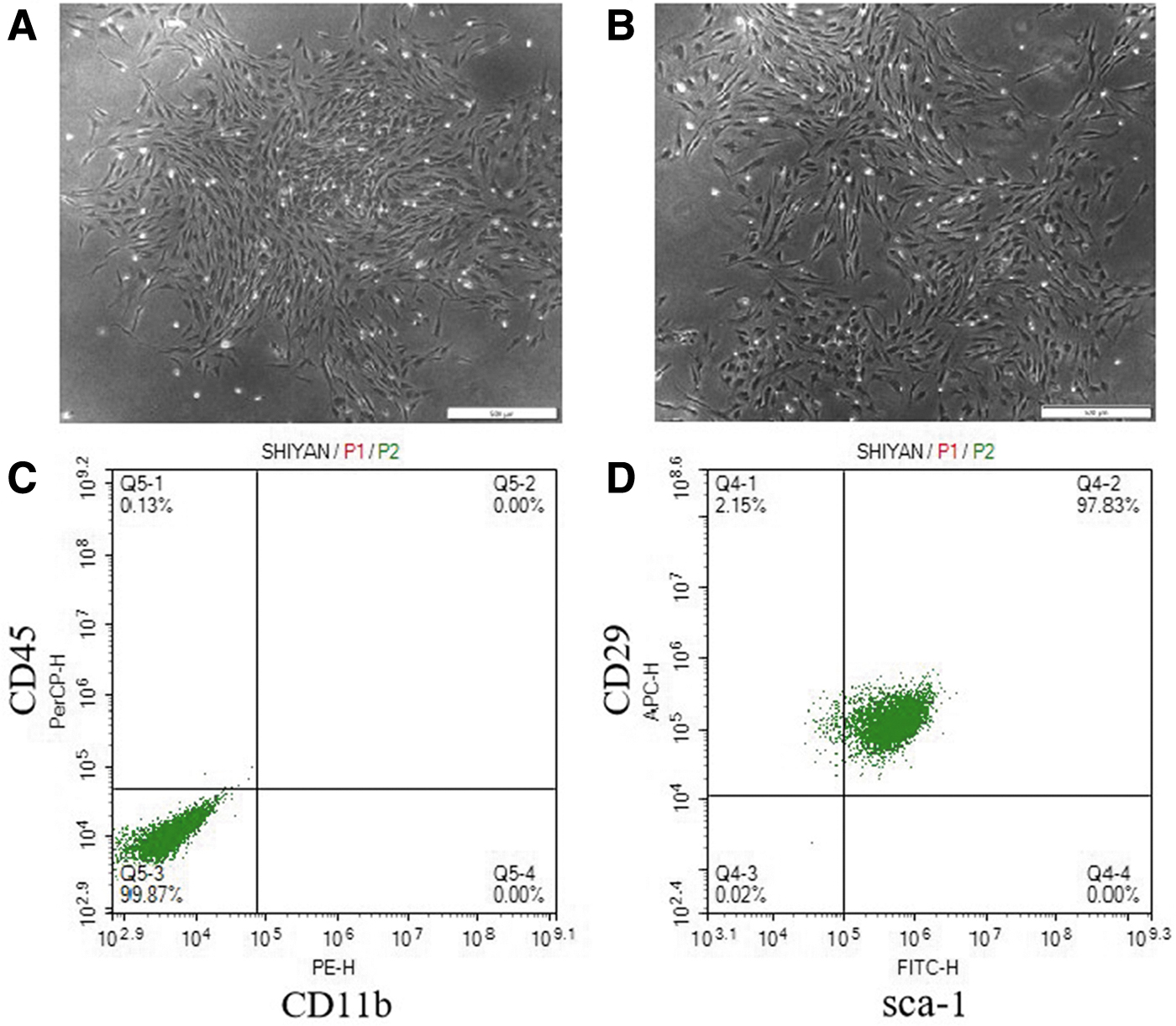
Identification of BMSCs.
Third-generation BMSCs were used for osteogenic lipogenesis induction culture. To explore the effect of seipin on BMSC differentiation, osteogenic and adipogenic differentiation of BMSCs was observed. Alizarin Red staining revealed that the number of bone nodules increased in BMSCs of SKO mice after induction of osteogenic differentiation for 21 days. However, there was no significant difference in the average size of bone nodules after induction of osteogenic differentiation in vitro (Fig. 4A–D).
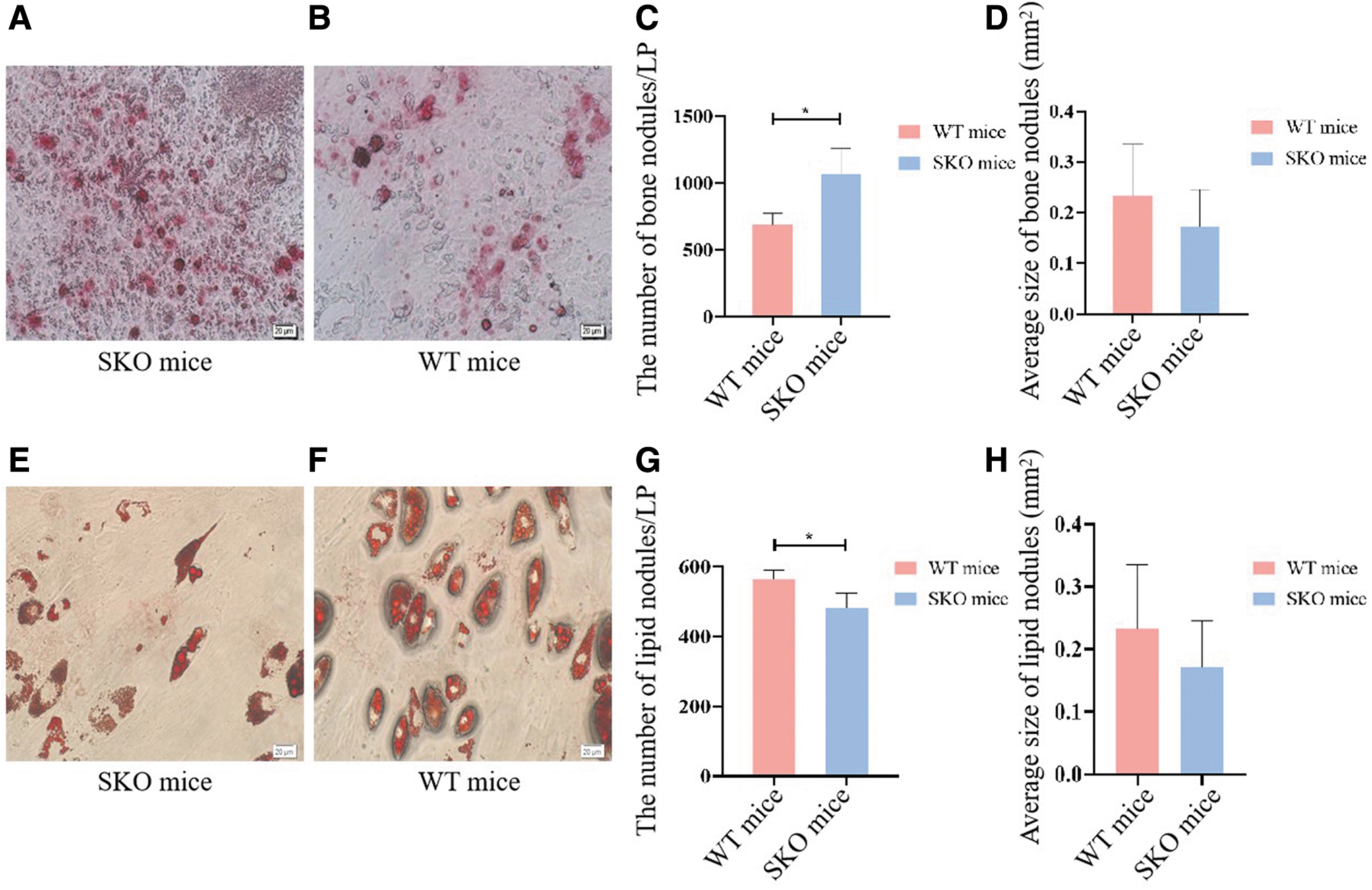
Osteogenic and adipogenic differentiation of BMSCs.
Adipocyte formation was detected after adipogenic induction culture for 21 days, and cells were stained with Oil Red O. The number of LDs was lower in BMSCs of SKO mice compared with WT mice (Fig. 4E–H).
Effect of seipin on proliferation of BMSCs
The plate colony assay was performed to determine the effect of seipin absence on BMSC proliferation. After culturing for 14 days, the total rate of the colony formation ability in the WT group was 3.97% ± 0.35%, while it was 2.27% ± 0.4% in the SKO group. The results demonstrated that the rate of colony-forming units in BMSCs of SKO mice was lower than in WT groups (P < 0.05) (Fig. 5).
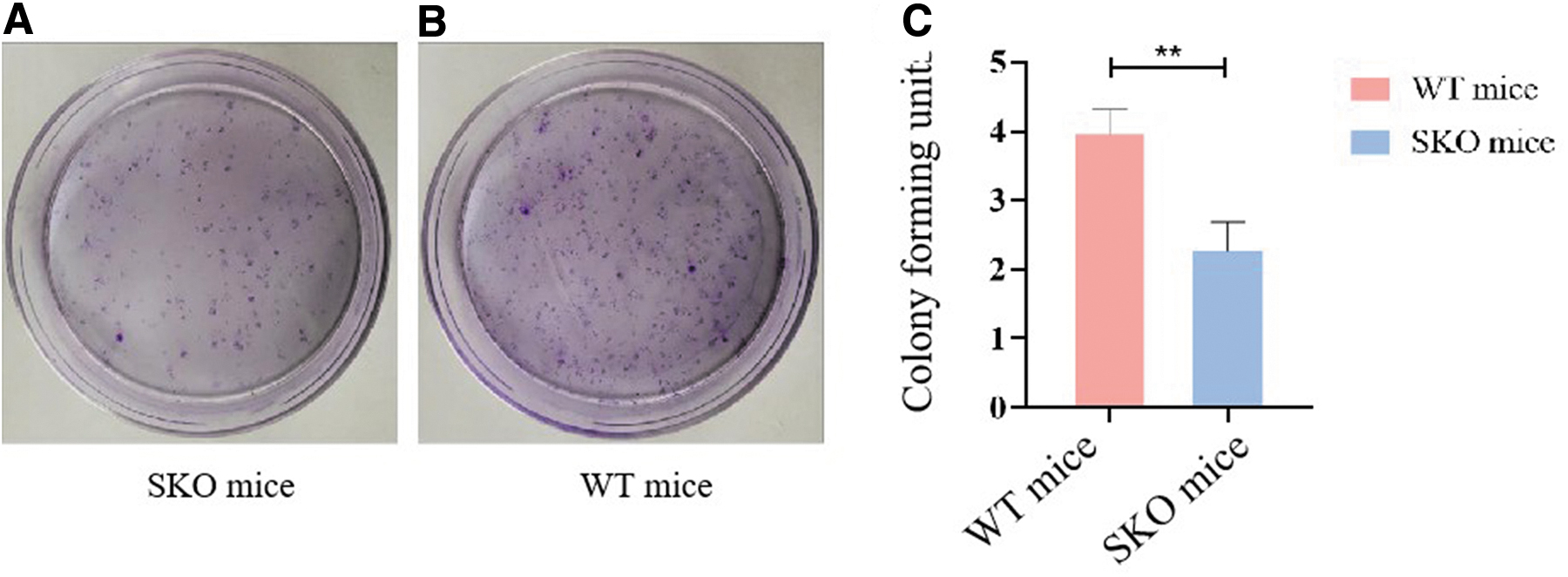
Colony-forming units of seipin-KO BMSCs.
Seipin deficiency reduces PPARγ expression to alter osteogenic and adipogenic differentiation of BMSCs
To further elucidate the effect of seipin on adipogenic differentiation and osteogenic differentiation, seipin expression in BMSCs from WT mice was suppressed by shRNA-mediated seipin knockdown. By screening of 3 shRNAs from lentiviral expression, we identified one shRNA that effectively suppressed seipin expression. Western blots showed significant reduction in seipin expression in seipin-deficient BMSCs.
qRT-PCR and Western blotting showed increased mRNA and protein levels of adipocyte-specific and osteoblast-specific genes, including PPARγ, enhancer-binding protein alpha (C/EBPα), and runt-related transcription factor 2 (RUNX2). After adipogenic and osteogenic induction, PPARγ and C/EBPα were significantly downregulated during adipogenesis with low expression of seipin, while RUNX2 was significantly increased.
Oil Red O staining showed reduced lipidogenic differentiation of the seipin-deleted BMSCs, while phosphatase [alkaline phosphatase (ALP)] staining and Alizarin Red staining indicated enhanced osteogenic differentiation of seipin-deleted BMSCs (Figs. 6 and 7).
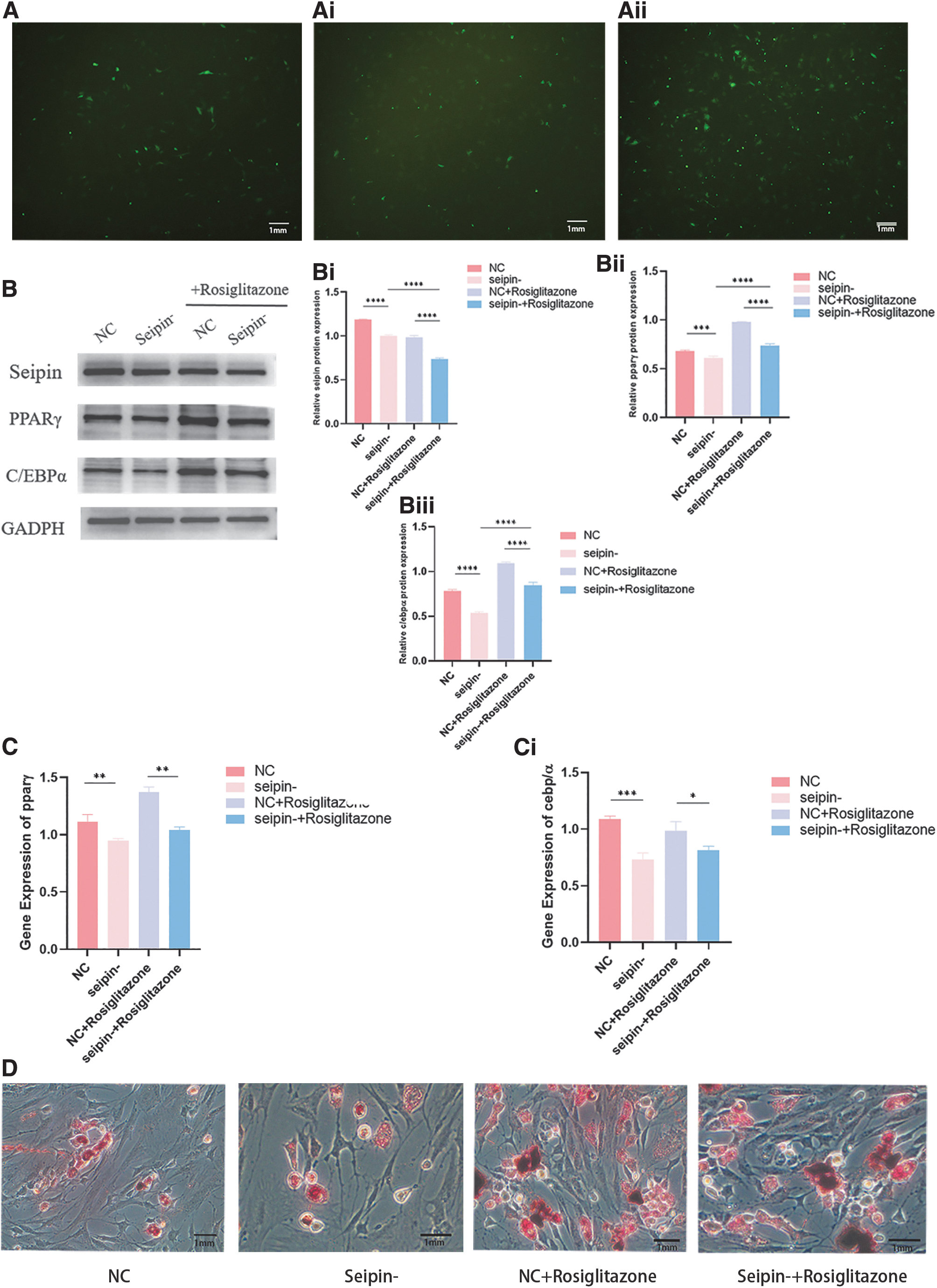
Seipin knockdown inhibits the adipogenic differentiation of BMSCs.

Seipin knockdown promotes the osteogenic differentiation of BMSCs.
To verify whether PPARγ is the link between adipogenic and osteogenic differentiation in the absence of seipin, seipin-deficient BMSCs were treated with the PPARγ-specific activator, rosiglitazone. After lipogenic induction, activation of PPARγ partially restored the expression of PPARγ and C/EBPα. In addition, an increase in Oil Red O staining intensity indicated that seipin-deficient BMSCs partially restored the potential for lipogenic differentiation.
After osteogenesis induction, PPARγ expression was upregulated in the rosiglitazone-treated group, while RUNX2 expression was downregulated, and the intensity of ALP and Alizarin Red staining was reduced. (Fig. 7).
Discussion
The seipin gene was screened by Magré J in 9 families with congenital generalized lipodystrophy (CGL) in 2001 [14]. BSCL2 is caused by seipin mutation. However, the relationship between seipin and the balance of lipid and bone metabolism has not been found. In this study, we established a seipin–/– mouse model to explore the effects of seipin on BMD, Tb.Th, and osteogenic–lipogenic differentiation of BMSCs, providing new ideas for the pathogenesis and treatment of lipid metabolism disturbance.
Patients with seipin deficiency have clinical features of lipoatrophy. With hepatic steatosis, patients usually demonstrate hepatomegaly, dyslipidemia, insulin resistance, and hyperinsulinemia. Patients have severe loss of adipose tissue throughout the body [15]. Due to the severe absence of adipose tissue, patients show a lower body weight. Low body weight is considered a risk factor for osteoporosis, yet patients have been reported to typically demonstrate higher BMD [16]. The seipin–/– mice demonstrated key phenotypic features similar to patients with BSCL2.
Seipin deficiency leads to a lack of subcutaneous adipose and lipid translocation deposition [17]. Nettebrock and Bohnert found a relationship between morphological changes in the endoplasmic reticulum and lipid abnormalities in SKO mice [18]. McGrath et al. found that seipin KO mice show an increased density mainly at the trabecular sites, while the cortical sites, such as the radial diaphysis, show decreased density [19]. Seipin mutations may impair homeostasis in bone metabolism and lipid metabolism.
Lipid metabolism and bone metabolism are thought to be mutually exclusive processes. Statins that regulate lipid metabolism have been shown to improve bone density and quality. Bone homeostasis and fat metabolism are mainly maintained by osteoblasts and adipocytes, with MSCs being their common ancestor [20]. BMSCs are adult stem cells with the potential of multiple differentiation, and BMSCs can differentiate into adipocytes and osteocytes. The isolated and purified BMSCs were used to explore the effect of seipin on cells.
There are two stages of MSC differentiation into adipocytes: MSC differentiation into the preadipocyte stage and preadipocyte differentiation into the functional mature adipocyte stage [21]. Prieur et al. found that mouse embryonic fibroblasts (MEFs) of seipin–/– mice were not different from WT mice at the beginning of adipogenic differentiation. However, the adipocytes of seipin–/– MEFs lacked intracellular LDs at the later stage of differentiation [22].
Increasing evidence has shown that osteogenesis and adipogenesis during MSC differentiation can transform into each other. Factors that inhibit differentiation of MSCs to adipocytes can promote their differentiation to osteoblasts [23]. These results suggest that seipin is a suppressor of MSC differentiation into adipocytes, but promotes MSC differentiation into osteoblasts.
Seipin deficiency causes glucose intolerance and insulin synthesis by reducing PPARγ in islet β-cells [13]. Li et al. showed that the proliferative capacity of stem cells in the adult hippocampal dentate gyrus was significantly reduced in seipin–/– mice compared with WT mice, which was associated with a reduction in PPARγ [24]. Bscl2 deficiency suppresses ADD1/Srebp1c upregulation, which leads to the decreased expression of key transcription factors such as PPARγ and C/EBPα, lipin 1, and AGPAT2 [25].
Srebp1c expression could not be induced in seipin-deficient cells. Instead, Srebp1c both directly activates the transcription of adipogenic genes and enhances the production of endogenous ligands for PPARγ [26]. The loss of SREBP1c-dependent PPARγ ligand production may contribute to decreased PPARγ and C/EBPα expression in maturing adipocytes lacking seipin. Seipin deficiency leads to an abnormal accumulation of phosphatidic acid (PA) in yeast, and accumulation of PA can inhibit the activity of PPARγ [27].
Huanxian et al. found that the increased GSK3β activity by seipin deficiency was sensitive to the PPARγ activator [28]. The severity of fatty liver in seipin knockout mice improved after PPARγ activator administration [29].
PPARγ is required in the process of adipose differentiation. C/EBPb and C/EBPm can regulate the adipogenic program required to promote adipocyte differentiation by inducing PPARγ. PPARγ and C/EBPα maintain fat cell differentiation through a positive feedback pathway [30]. Several transcription factors of adipogenesis (C/EBPβ, C/EBPδ, STAT5, and EBF1) induce the expression of PPARγ.
However, transcriptional repressors of adipogenesis (GATA2, KLF2, and CHOP) have been shown to have decreased PPARγ expression [31]. In addition to its role in adipocyte differentiation, PPARγ is also crucial for controlling gene networks involved in bone formation. Osteoblast differentiation and bone formation have been shown to be inhibited by PPARγ [32].
Xiong et al. found that 15d-PGJ2 inhibits osteoclastogenesis by regulating the Wnt/β-catenin signaling pathway through the PPARγ-dependent pathway, thereby reducing bone formation in bone metabolism [33]. Increased expression of PPAR in bone marrow stromal progenitors was found to lead to increased adiposity and inhibition of osteoblastogenesis, probably by suppressing expression of the pro-osteogenic transcription factor, Runx2 [34].
Stem cells have shown great therapeutic potential in the treatment of human diseases such as osteoporosis [35], diabetes [36], and hepatic disorders. MSCs are pluripotent adult stem cells characterized by self-renewal capacity and multidirectional differentiation. MSCs contribute to the development, maintenance, and repair of tissues without immune rejection when MSCs are transplanted [37]. Ma et al. found that MSCs can treat type 2 diabetes mellitus (T2DM) and lipid metabolism disorders in mice by lowering fasting blood glucose levels and increasing fasting insulin levels [38].
Liu et al. showed that transplantation of adipose-derived MSCs significantly improved lipid metabolism disturbance in mice [39]. Exosomes derived from MSCs can activate autophagy through the AMPK pathway and improve hepatic glucose and lipid metabolism in T2DM rats [40]. Therefore, it is possible that lipid metabolism disturbance in BSCL2 patients can be treated by regulating the differentiation of BMSCs.
Conclusions
The absence of seipin could lead to the reduced proliferation ability of BMSCs, decreased adipogenesis ability, and increased osteogenic differentiation.
Footnotes
Acknowledgments
The authors thank Rong Qi and Guoqing Liu for technical assistance. Seipin knockout homozygous mice were provided by the Peking University Institute of Cardiovascular Sciences, Beijing 100191, China.
Ethics Approval and Consent to Participate
All animal handling procedures in this study were approved by the Research Ethics Committee of the First Affiliated Hospital, School of Medicine, Shihezi University, and conformed to the National Institutes of Health Guide for the care and use of laboratory animals.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (82060054); the Discipline Construction Fund of Central People's Hospital of Zhanjiang (2022A15, 2022A16); the Zhanjiang Science and Technology Special Fund project—the special topic for basic research (2022A01028); and the High-level Hospital Construction Special Fund of Central People's Hospital of Zhanjiang—key projects of disease prevention and control (2022A01103, Major Projects: 2023A214).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2