
Abstract
Mesenchymal stromal cells have shown promising effects in the treatment of immune thrombocytopenia. However, the underlying mechanisms are not fully understood. In this study, we investigated the therapeutic effects of human bone marrow mesenchymal stromal cells (hBMSCs) and analyzed their unique role in regulating the M1/M2 macrophage ratio. We established a passive immune thrombocytopenia (ITP) mouse model and showed that there was a significant M1/M2 imbalance in ITP model mice by assessing the M1/M2 ratios in the liver, spleen, and bone marrow; we observed excessive activation of M1 cells and decreased M2 cell numbers in vivo. We have shown that systemic infusion of hBMSCs effectively elevated platelet levels after disease onset. Further analysis revealed that hBMSCs treatment significantly suppressed the number of proinflammatory M1 macrophages and enhanced the number of anti-inflammatory M2 macrophages; in addition, the levels of proinflammatory factors, such as interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α), were significantly decreased in vivo, while the levels of the anti-inflammatory factor interleukin-10 (IL-10) were increased. In conclusion, our data suggest that hBMSCs treatment can effectively increase platelet counts, and the mechanism is related to the induction of macrophage polarization toward the anti-inflammatory M2 phenotype and the decrease in proinflammatory cytokine production, which together ameliorate innate immune disorders.
Introduction
Immune thrombocytopenia (ITP) is an acquired autoimmune disease characterized by thrombocytopenia, and among blood disorders, it is the most common bleeding disorder [1]. However, the pathogenesis of ITP is complex and heterogeneous, involving multiple aspects of the immune system [2,3]. Antiplatelet antibody (APA) production [4], T-cell dysfunction, such as imbalances of the Th1/Th2 and Th17/regulatory T cell (Treg) ratios [5 –7], and abnormal plasmacytoid dendritic cell ratios [8] contribute to its pathogenesis. In particular, macrophages play a prominent role in ITP, both as effector cells that phagocytose platelets and as antigen-presenting cells that activate humoral and cellular immunity, promoting immune disorders in ITP and exacerbating thrombocytopenia [9].
Macrophages are highly variable and transform into different phenotypes depending on the environment. Macrophages polarize toward the M1 phenotype in response to stimulation by tumor necrosis factor-α (TNF-α), bacterial lipopolysaccharide, and interferon-γ (IFN-γ). They perform proinflammatory, antimicrobial, and antitumor functions by releasing high levels of proinflammatory factors such as TNF-α, interleukin-6 (IL-6), IL-1, and iNOS. In contrast, with the stimulation of interleukin-4 (IL-4), IL-13, and interleukin-10 (IL-10), macrophages polarize toward the M2 phenotype and participate in immunomodulation, angiogenesis, and tumor progression via the production of anti-inflammatory factors (IL-10, transforming growth factor-beta [TGF-β]) [10,11]. The M1/M2 polarization state can disrupt the balance of lymphoid subpopulations, thereby promoting the development of immune diseases [12].
Furthermore, an imbalance in the M1/M2 ratio has been observed in patients with ITP, where macrophages are hyperpolarized toward the M1 phenotype, leading to an increase in proinflammatory factors and aggravating the disruption of humoral and cellular immunity; M1 macrophages can repolarize toward the M2 phenotype when exposed to high doses of dexamethasone or eltrombopag [13,14]. Therefore, an imbalance in macrophage polarization is a key potential therapeutic target for ITP.
Mesenchymal stromal cells (MSCs) are widely used in the treatment of autoimmune diseases due to their promising immunomodulatory ability [15]. MSCs act on lymphocytes, macrophages, and plasmacytoid dendritic cells by secreting soluble mediators (TGF-β, PGE2, IL-6, IL-10, IL-10), which maintain the immune environment in vivo [16]. In recent years, MSCs have been shown to reverse the polarization of macrophages under inflammatory conditions, both in vivo and in vitro [17,18]. In animal models of rheumatoid arthritis (RA) [19], Sjogren syndrome (SS) [20], systemic lupus erythematosus (SLE) [21], and inflammatory bowel disease (IBD) [22], MSCs have been shown to effectively reverse the M1 phenotype of macrophages to an anti-inflammatory M2 phenotype, resulting in improved disease alleviation.
Compared to MSCs from other sources, bone marrow mesenchymal stromal cells (BMSCs) also modulate the hyperpolarization of macrophages toward the M1 phenotype under inflammatory conditions [23]; in particular, BMSCs are more accessible and as such are the only MSCs that can promote hematopoietic stem cell maturation in vivo [24]. Hence, we selected BMSCs as an option for ITP treatment. Establishing the effectiveness of MSCs through animal studies is essential before their clinical application in ITP treatment. Human-derived MSCs, with their low immunogenicity, have demonstrated efficacy in a range of disease models, making them a feasible option for utilization in animal models [25,26]. Consequently, in this study, we utilized human BMSCs to treat a mouse model of ITP and assessed their therapeutic efficacy.
In our previous study, it was demonstrated that BMSCs can effectively elevate platelets in ITP model mice and can increase the levels of Treg cells [27]. However, it remains unknown whether there is an imbalance in macrophage polarization in ITP model mice and whether human BMSCs can improve symptoms in ITP mice and reverse the M1/M2 imbalance. In the current work, we assessed platelet profiles and M1/M2 macrophage polarization in multiple immune organs after transplanting human bone marrow mesenchymal stromal cells (hBMSCs) into ITP model mice. The therapeutic effect of hBMSCs may be related to the altered macrophage polarization status.
Materials and Methods
Cell culture
Extraction and culture of hBMSCs were performed by Guangzhou Saliai Stem Cell Science and Technology Co. Ltd. Cryopreserved P2 generation cells were routinely recovered by thawing with 10% fetal bovine serum (FBS)-supplemented Dulbecco's modified Eagle's medium (DMEM) and suspended in complete medium (DMEM, 10% FBS, 2 mM
The murine RAW264.7 macrophage cell line was purchased from Procell (CL-0233, Shanghai, China) and was cultured in DMEM (10-040-CVR; Corning) supplemented with 10% FBS and 1% penicillin‒streptomycin. Cells were plated in T-25 tissue culture flasks and incubated in a humidified atmosphere at 37°C in 5% CO2.
Generation of passive ITP model and cell transplantation
This study used BALB/c mice to establish the model (n = 24, 18–20 g; 8–10 weeks of age), which were purchased from Charles River Laboratories. This study was reviewed and approved by the Sun Yat-Sen University Animal Medicine Center Ethics Committee, approval number SYSU-IACUC-2023-000150. As previously described [28], we established a dose-escalating mouse model of passive ITP. The mice received intraperitoneal injections of escalating doses of monoclonal MWReg30 (BD Pharmaingen) diluted in 200 μL of phosphate-buffered saline (PBS). The mice were injected with different doses of antibodies daily (68 μg/kg on days 1 and 2, 102 μg/kg on day 3, and 136 μg/kg on days 4–8), and hBMSCs were injected via the tail vein on day 3.
In addition, BALB/c mice were divided into three groups: control group (daily intraperitoneal injection of IgG1 + 200 μL PBS), ITP group (modeled as above), and ITP+MSC group (tail vein injection of 5 × 107/kg quantity of hBMSC on the third day of modeling).
Coculture of hBMSCs with M1 macrophages
Murine RAW264.7 macrophages were stimulated with LPS (100 ng/mL; ELGBIO, Guangzhou, China) plus IFN-γ (30 ng/mL; Chamot Biotech, Shanghai, China) and considered the M1 group. RAW264.7 cells were seeded at 2 × 106 cells/well in the lower chamber of a six-well plate and then stimulated for 12 h as described above. On the following day, hBMSCs were placed into 0.4 mm pore size Corning Transwell culture plate inserts at a density of 4 × 105 cells/well (Corning). The coculture time was 24 h. After incubation, the macrophages were collected for further experiments. The cells and supernatants were collected and stored at −80°C for further use.
Cytokine level analysis and platelet level testing
On day 8 of modeling, 100 μL of blood was collected from the medial canthal vein. Subsequently, serum was separated by centrifugation at 2,500 rpm for 15 min and stored at −80°C for subsequent analysis. The levels of interleukin-1β (IL-1β), IL-6, IL-4, interleukin-17 (IL-17), interleukin-2 (IL-2), IL-10, IFN-γ, TNF-α, and TGF-β in mouse plasma as well as in coculture supernatants were measured by enzyme-linked immunosorbent assay (ELISA) kits (Mlbio or ELGBIO). The absorbance was measured at 450 nm. The cytokine concentration was calculated by drawing a standard curve based on cytokine standards.
Platelet counts were measured by daily blood collection from the same medial canthus vein as before antibody injection, and blood was collected using EDTA anticoagulation tubes. Then, the platelet levels were measured by an automatic blood cell analyzer (BC-2800vet; Mindray).
Flow cytometry
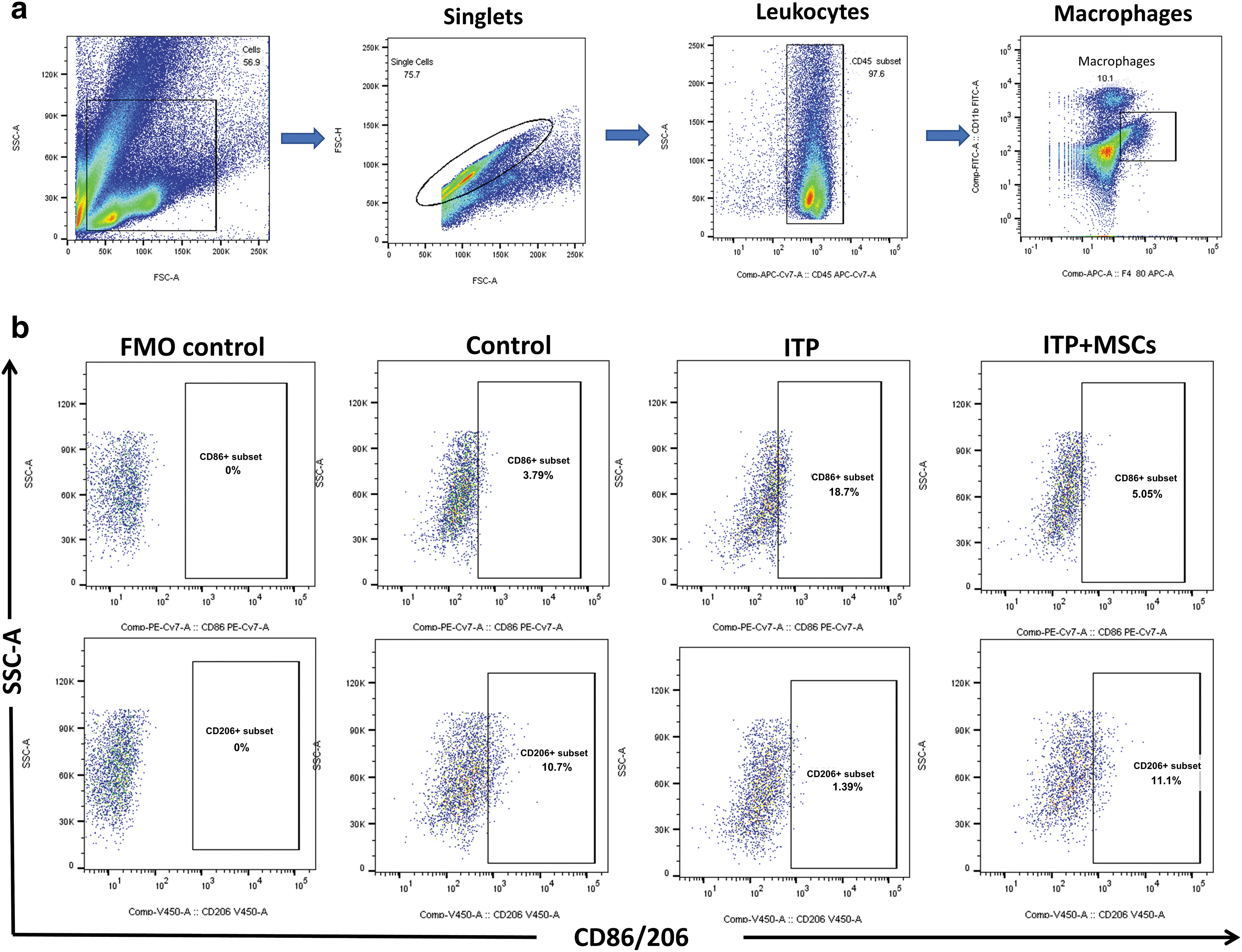
RAW264.7 cells were collected, and an Fc receptor blocking agent (BD Biosciences) was added and incubated for 5 min at 4°C to block the Fc receptors. RAW 264.7 cells were incubated with anti-mouse CD45-APC-Cy7, F4/80-APC, CD86-PE-Cy7, and CD206-BV421 (BioLegend) in the dark at 4°C for 30 min and then washed in PBS, and CD206 needs to be permeabilized before addition.
To assess the M1/M2 polarization of macrophages, mice were sacrificed by cervical dislocation on day 8, spleens and livers were collected, and each organ was ground and processed into a single-cell suspension using a cell strainer (70 μm; Jet Bio-Filtration, GuangZhou, China). Tibias and femurs were dissected, and bone marrow was harvested by repeated flushing with glucose-supplemented DMEM through a 1 mL syringe. The bone marrow flush was centrifuged at 250g for 5 min, and after filtering using a cell strainer, the mononuclear cells were resuspended in PBS and the operation was repeated twice. Erythrocytes were removed from single-cell suspensions of the spleen, liver, and bone marrow by incubation for 20 min with red cell lysis buffer (Abisin). After two rounds of centrifugation and resuspension in PBS, the cells were blocked for 5 min at 4°C using Mouse BD Fc Block (BD Biosciences).
Single cells were washed three times with PBS, incubated with anti-mouse CD11b-FITC, CD45-APC-Cy7, F4/80-APC, CD86-PE-Cy7, and CD206-BV421 (BioLegend) for 30 min at 25°C, and CD206 needs to be permeabilized before addition. The ultimate assay was conducted using a FACSCalibur analyzer from BD Biosciences. Moreover, wherein the inclusion of suitable control groups was prioritized to facilitate the auxiliary flow cytometry's gating process. The data were processed using FlowJo software (Ashland, OR).
Western blotting analysis
Frozen bone marrow and splenocytes were thawed and homogenized in ice-cold lysis reagents. Both bone marrow and spleen specimens underwent the same processing procedure as the samples in flow cytometry and were also subject to lysed red blood cell. Extracted proteins were quantified with a BCA protein analysis kit (Beyotime). Ten micrograms of protein was separated by sodium dodecyl sulfate‒polyacrylamide gel electrophoresis (SDS‒PAGE) and transferred onto polyvinylidene difluoride (PVDF) membranes. After blocking in Fast Blocking Solution for 20 min, the membranes were incubated with primary antibodies against iNOS (1:1,000, bs90716; Bioworld), mannose receptor (1:1,000, ab64693; Abcam), and GAPDH (1:2,000,AP0063; Bioworld) at 4°C overnight. The binding of specific antibodies was detected with the appropriate HRP-conjugated secondary antibody (1:500, No. 7076s; Cell Signaling). The protein bands were detected by a multispectral imaging system.
Statistical analysis
All the results are presented as the mean ± standard deviation. The obtained data were analyzed using GraphPad Prism 8 (GraphPad Software). Analysis of variance (ANOVA) was used for comparisons between groups, and Tukey's test was used when ANOVA results were significant. P ≤ 0.05 was considered a statistically significant difference.
Results
hBMSCs reverse inflammatory M1/M2 macrophage imbalances in vitro
We initially evaluated whether hBMSCs can ameliorate the dysregulation of inflammatory macrophages in vitro. Therefore, RAW264.7 cells were stimulated with LPS+IFN-γ and cocultured with hBMSCs in a Transwell system for 24 h. We used flow cytometry to measure CD206 and CD86 levels to evaluate M1 and M2 cell ratios. The results showed that the RAW264.7 (LPS+IFN-γ) group had a decrease in CD206+ macrophages and an increase in CD86+ macrophages compared to the RAW264.7 (LPS+IFN-γ)+BMSCs group, but neither was statistically significant. However, further analysis showed that the M1/M2 ratio in the RAW264.7 (LPS+IFN-γ)+BMSCs group (6.701 ± 2.130) decreased significantly compared to that in the RAW264.7 (LPS+IFN-γ) group (11.35 ± 2.340, P < 0.02), recovering to normal levels in a statistically significant manner (Fig. 1). In summary, hBMSCs can indeed ameliorate the imbalance in the M1/M2 ratio under inflammatory conditions in vitro.

In vitro, hBMSCs effectively restored an imbalanced M1/M2 macrophage polarity. RAW264.7 group represented the untreated control group. The RAW264.7 (LPS+IFN-γ) group received pretreatment with LPS and IFN-γ for 12 h for subsequent evaluation. The RAW264.7(LPS+IFN-γ)+BMSCs group, however, underwent 12 h of exposure to inflammatory factors, followed by a 24 h incubation with hBMSCs, before being analyzed for polarization utilizing flow cytometry.
Administration of hBMSCs effectively elevates platelet levels in ITP model mice
To evaluate the therapeutic effect of BMSCs, we transplanted hBMSCs into our mouse model on the third day of modeling, which was also the day on which the mice had the lowest platelet counts, and observed the change in platelet counts daily. As shown in Fig. 2a, the platelet levels were significantly higher in the ITP+MSCs group on days 5, 6, and 8 than in the ITP group. Consistent with the test results, the petechiae at the injection sites were alleviated in the treated group compared to the untreated group (Fig. 2b, c).
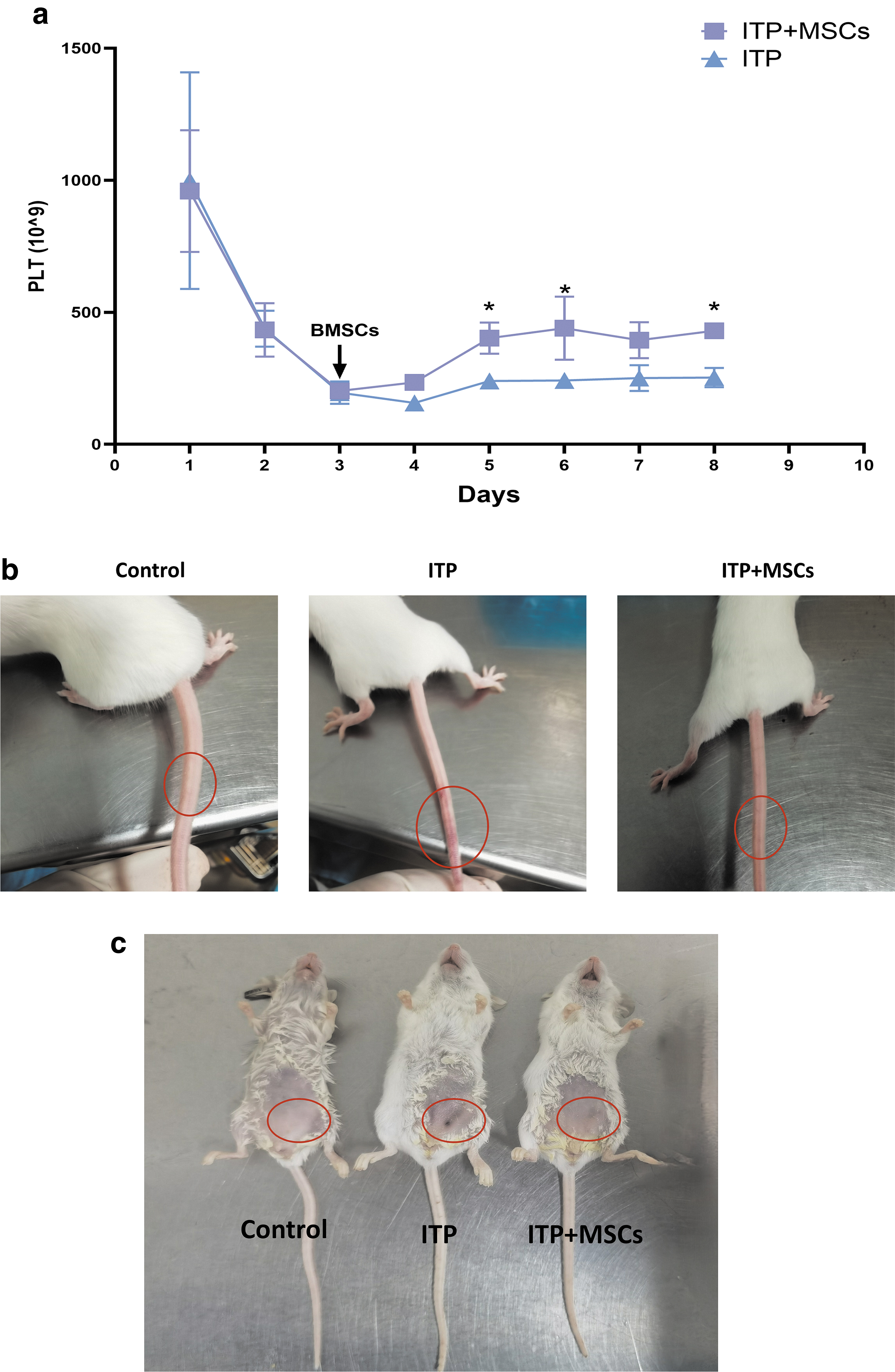
Recovery of platelets and petechiae in each group. The control group received daily intraperitoneal administration of IgG1 + 200 μL PBS, while the ITP group was used to establish a passive ITP mouse model. On the contrary, the ITP+MSCs group received a tail intravenous injection of 5 × 107/kg hBMSCs on the third day of model induction.
hBMSCs reverse inflammatory M1/M2 macrophage imbalance in vivo
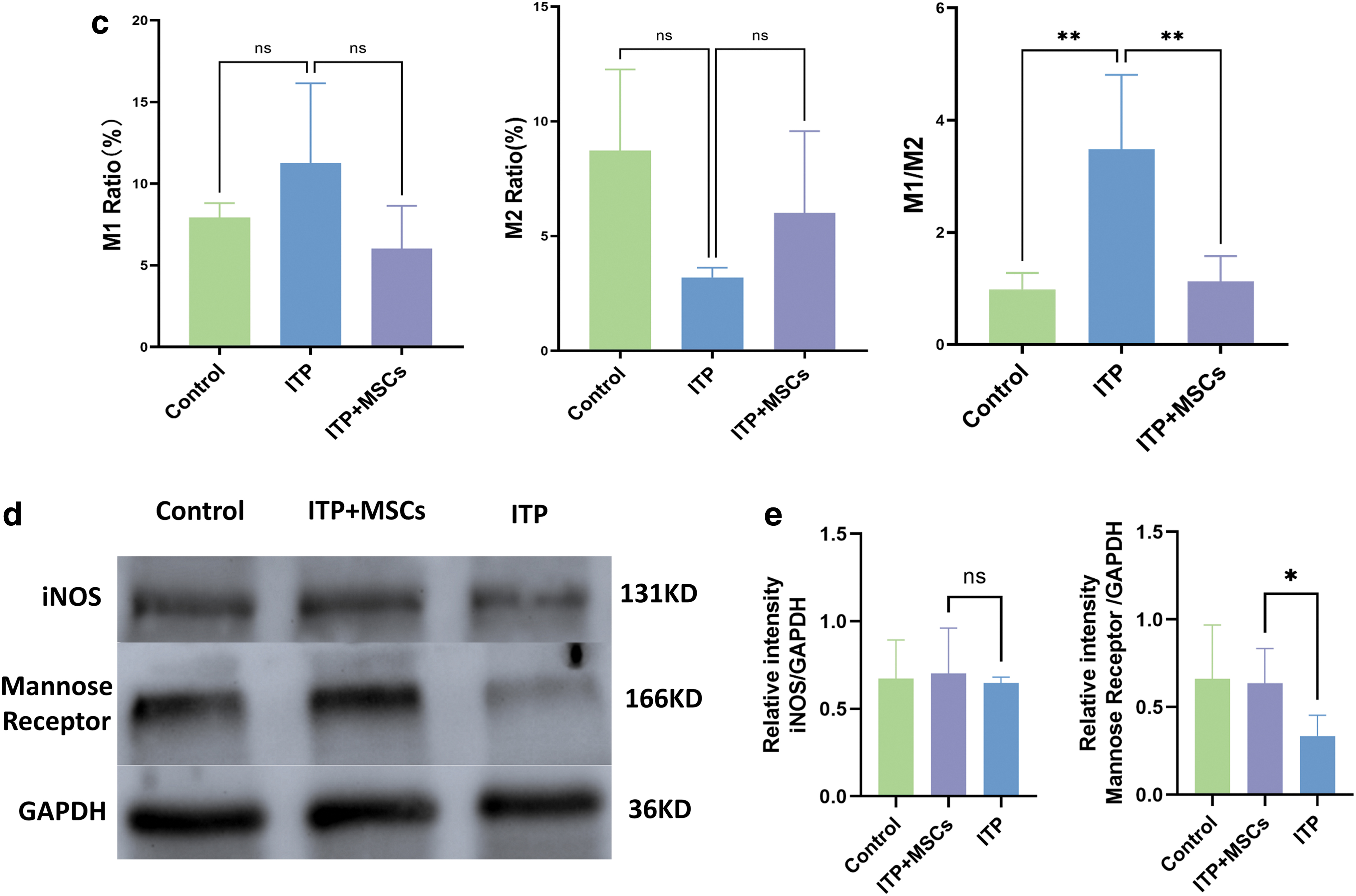
Imbalance in the M1/M2 macrophage ratio promotes inflammatory diseases. To verify whether hBMSCs treatment can improve the M1/M2 imbalance in ITP model mice, we assessed M1 and M2 cell levels in the bone marrow, spleen, and liver of mice by flow cytometry. As shown in the Figs. 3c, 4b, and 5c, the ITP group exhibited a significant upregulation of the M1/M2 ratio in the spleen, liver, and bone marrow, relative to the control group. Particularly, the dysregulation of the M1/M2 ratio was more pronounced in the liver and spleen compared to the bone marrow. We further analyzed the changes in M1 levels as well as M2 levels in each organ (bone marrow, liver, and spleen) after infusion of BMSCs.
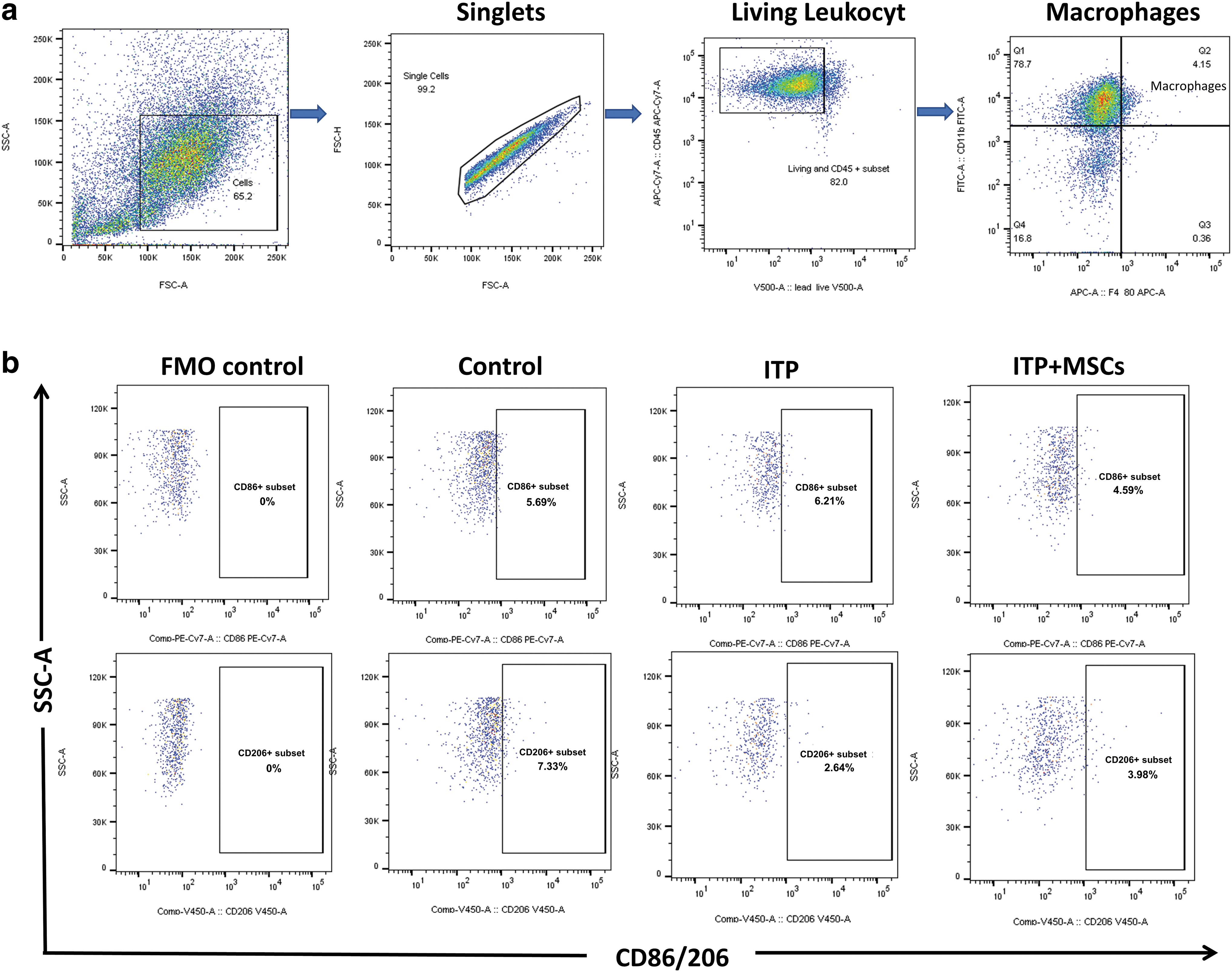
M1/M2 subtype ratio of bone marrow macrophages. The FMO was administered all antibodies except for CD86 or CD206, which were used specifically to establish flow cytometry gating. The control group received daily intraperitoneal administration of IgG1 + 200 μL PBS, while the ITP group was used to establish a passive ITP mouse model. On the contrary, the ITP+MSCs group received a tail intravenous injection of 5 × 107/kg hBMSCs on the third day of model induction.
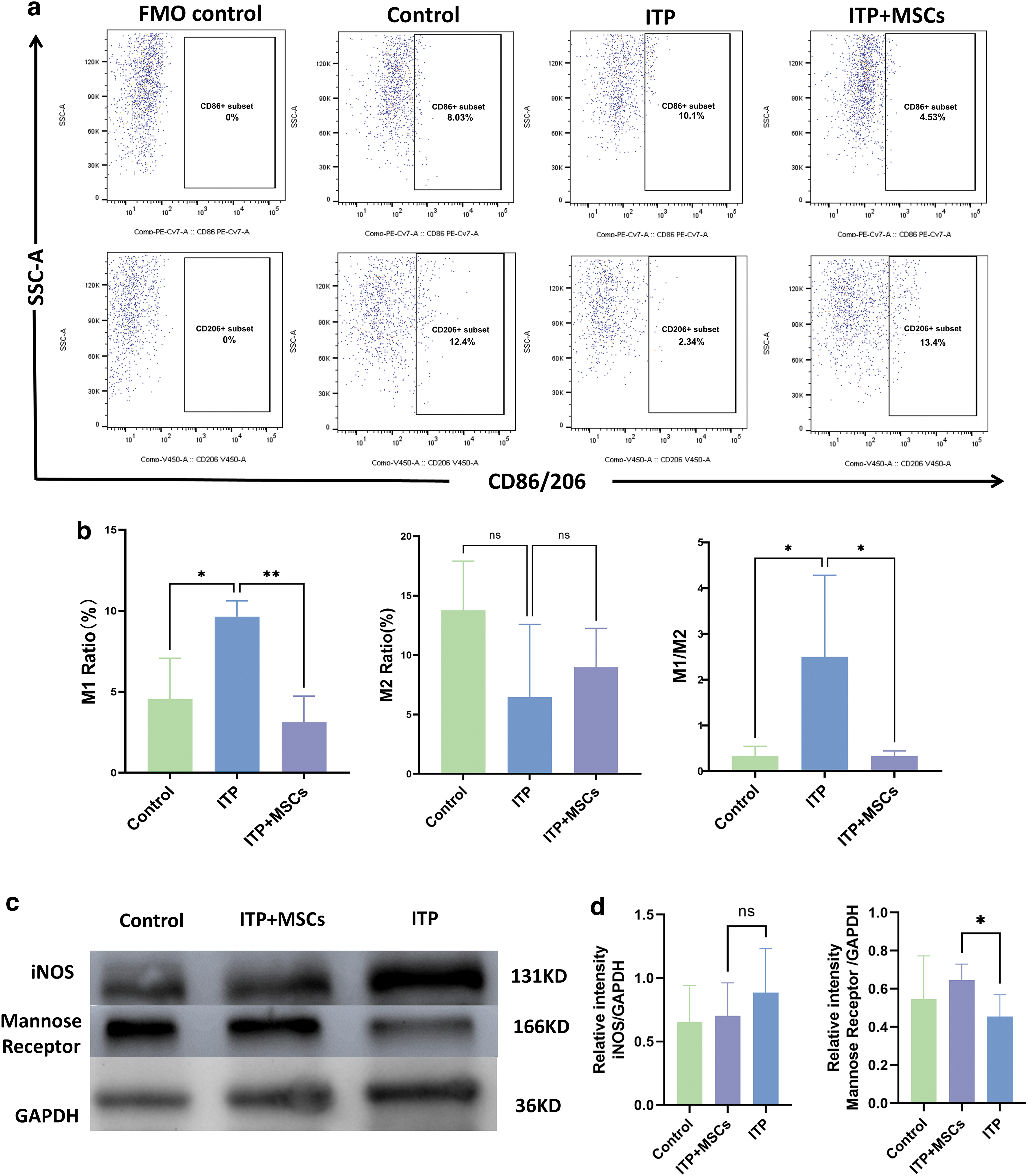
M1/M2 subtype ratio of spleen macrophages. The grouping and flow cytometry circle gating strategy used in this study resemble those used for analyzing bone marrow cells.
We observed no significant difference in the M1 levels in bone marrow among the three groups, but in the liver and spleen, the M1 levels were lower in the ITP+MSCs group than in the ITP group (bone marrow: 6.022% ± 2.626% vs. 11.26% ± 4.895%, P = 0.10; spleen: 3.140 ± 1.591 vs. 9.643 ± 0.9693, P < 0.01; liver: 6.843 ± 2.909 vs. 20.32 ± 2.510, P < 0.0001, respectively; Figs. 3c, 4b, 5c). While a similar trend was observed for the M2 levels in the three organs, treatment with hBMSCs significantly increased the M2 levels (bone marrow: 6.008% ± 3.567% vs. 3.195% ± 0.4266%, P = 0.36; spleen: 8.975 ± 3.271 vs. 6.453 ± 6.131, P = 0.74; liver: 12.77 ± 4.252 vs. 2.142 ± 0.9408, P < 0.0005; Figs. 3c, 4b, 5c).
M1/M2 subtype ratio of liver macrophages by flow cytometry and serum levels of inflammatory factors.
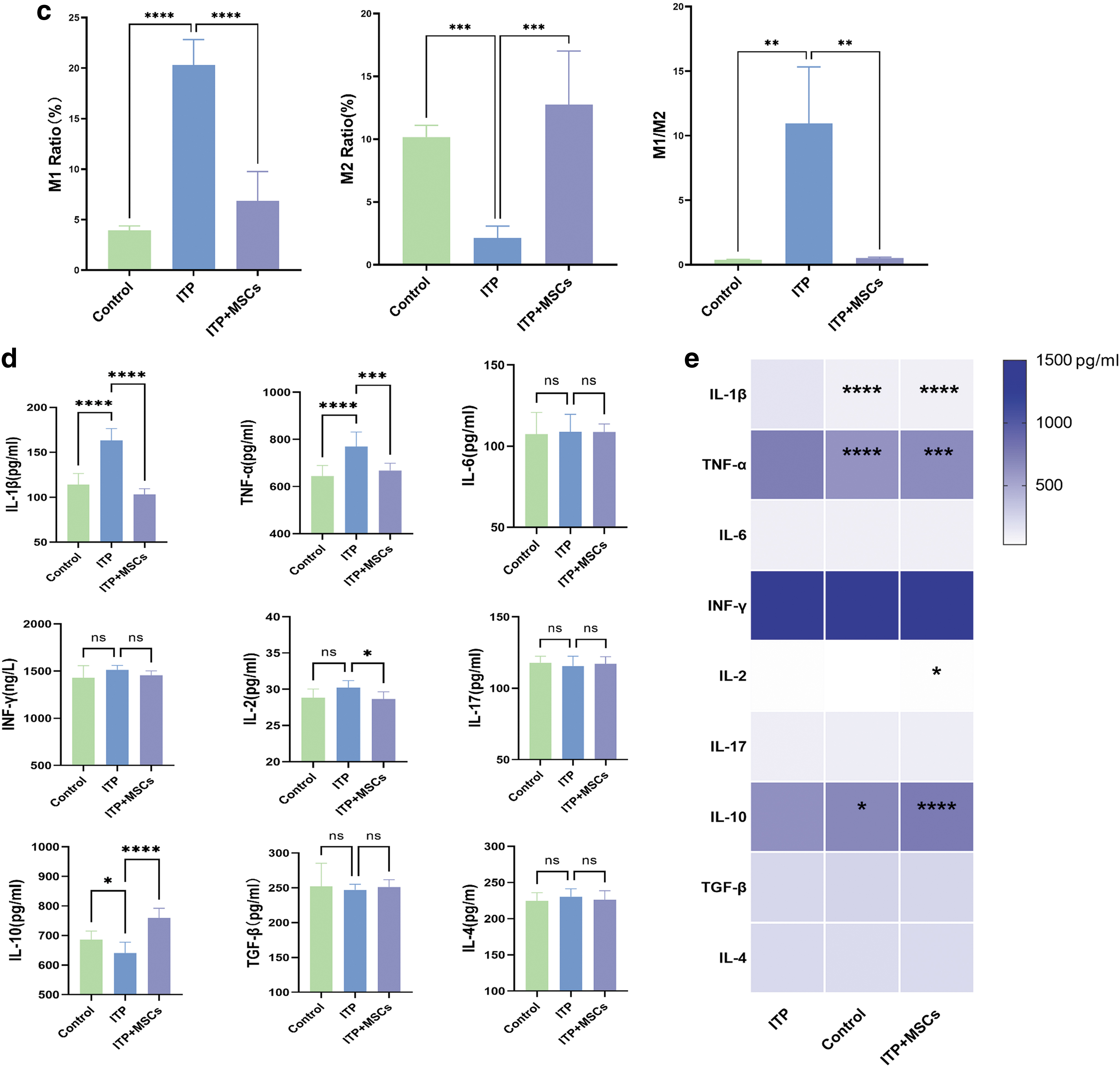
In addition, the M1/M2 macrophage ratio in the three organs in ITP+MSCs group was significantly lower than that in the ITP group. We also used western blotting to assess the expression levels of iNOS (an M1 macrophage marker) and mannose receptor (an M2 macrophage marker) in the bone marrow and spleen, and the results were consistent with the flow cytometry results (Figs. 3d and 4c; The Western blot full membrane image is provided in the Supplementary Materials), which further proved that the BMSCs treatment reversed the M1/M2 imbalance in ITP model mice.
We examined nine inflammatory factors associated with macrophages in the three groups of mice. The levels of the TNF-α and IL-1β proinflammatory cytokines, which are released by activated M1 macrophages, were significantly higher in the ITP group than in the control group, further suggesting that M1 macrophages may be involved in the pathogenesis of ITP. The administration of hBMSCs resulted in a significant decrease in TNF-α, IL-1β, and IL-2 proinflammatory cytokine expression. In contrast, as shown in Fig. 5d and e, the expression of the M2 macrophage-associated anti-inflammatory cytokine IL-10 was significantly upregulated in the ITP+MSCs group. Taken together, these findings suggest that hBMSCs can convert macrophages into an anti-inflammatory M2 phenotype, thereby alleviating the inflammatory state in vivo.
Discussion
In recent years, since the excellent immunomodulatory ability of MSCs was first demonstrated in 1998, they have been proved to be effective in the treatment of a variety of inflammatory diseases [29,30], and their various mechanisms of action can regulate immune imbalance via multiple pathways [31]. We demonstrated that in a passive ITP mouse model, there was a similar abnormality of M1/M2 polarization as in ITP patients. Disordered macrophages and the regulatory role of MSCs were described in three tissues and organs involved in the pathogenesis of ITP, including bone marrow, liver, and spleen. After treatment with hBMSCs, the platelet counts of ITP model mice were increased, M1/M2 polarization imbalance was alleviated, and plasma proinflammatory cytokine levels were significantly reduced.
Aberrant polarization of macrophages plays a significant role in promoting a variety of inflammatory diseases, and M1/M2 imbalances are found in SLE, RA, and IBD and correlated with disease severity [21,32,33]. In addition, in colitis as well as in diabetic nephropathy [34,35], disease progression was significantly alleviated by transplantation of M2 macrophages. This is solid evidence that M1/M2 imbalance plays a deleterious role in the development of inflammatory diseases, and targeting these imbalances may be a new approach for treatment. The M1/M2 imbalance is also observed in ITP patients, and it is alleviated after treatment [13,14]. We showed the same pathological changes in passive ITP model mice, with M1/M2 imbalance in the bone marrow, spleen, and liver, but the bone marrow showed predominantly reduced M2 macrophages, while the liver and spleen showed excessive activation of M1 macrophages and reduced M2 macrophage numbers. This is in accordance with the characteristics of macrophages from various tissues.
Zhao et al. [36] separated macrophages from bone marrow as well as spleen and found that splenic macrophages were polarized toward the M1 phenotype significantly more strongly than those from the bone marrow, while the potential for polarization toward the M2 phenotype was similar. This indicates that in vivo, under inflammatory conditions, bone marrow-derived macrophages and liver- and spleen-derived macrophages may respond differently. Macrophages in the liver and spleen are polarized toward the M1 phenotype in ITP patients, and they release large amounts of proinflammatory factors and engulf platelets in large numbers; the use of CD41-PLT-vincristine can effectively eliminate M1 macrophages and thus alleviate the disease [9].
Recently, Zhao et al. demonstrated the presence of excessively activated M1 macrophages among hepatosplenic macrophages in a passive mouse model according to several indexes, but they did not assess the level of M2 macrophages [37]. The application of anti-TNF-α therapy in ITP mice led to a decrease in the population of nonclassical monocytes and M1 macrophages improved platelet retention in the spleen and liver. This suggests that targeting the M1/M2 imbalance is an effective treatment for ITP.
Our team has already found that BMSCs can improve platelet levels in ITP model mice and can elevate the proportion of Treg cells [27]. Recent studies have also shown that exosomes from BMSCs can effectively elevate platelet levels and flatten and reverse the imbalance in the Th17/Treg ratio [38]. Although MSCs have been clinically used to treat a variety of diseases, the application of MSCs in ITP is still under exploration. Previous studies on the mechanism by which MSCs function in the treatment of ITP have mainly focused on acquired immune disorders such as Th17/Treg imbalances. Therefore, additional therapeutic mechanisms need to be explored to advance the use of these cells in clinical practice. The main pathological mechanism underlying ITP is the massive production of platelet antibodies; antiplatelet autoantibodies promote the phagocytosis of platelets by activated macrophages, mainly in the spleen and liver.
Therefore, macrophages contribute to the persistence of the autoimmune response as major antigen-presenting cells as well as effector cells [39,40]. Recently, there has been increasing evidence that MSCs can promote the conversion of M1 macrophages into M2 macrophages in vitro [41], which in turn exert immunomodulatory effects to alleviate diseases, and these cells have been found to have an excellent ability to regulate macrophage imbalance in autoimmune diseases such as RA, SLE, and IBD [42]. Similarly, we found that treatment of ITP model mice with hBMSCs promoted the polarization of anti-inflammatory M2 macrophages and reversed the M1/M2 imbalance, both in the liver and spleen as well as in the bone marrow; in addition, the platelet levels of the mice recovered.
We also measured M1 subtype- and M2 subtype-associated inflammatory factors; the M1-associated inflammatory factors IL-1β and TNF-α were significantly decreased, while the level of IL-10 was significantly increased, which further confirmed our suspicion.
In conclusion, hBMSCs treatment could effectively increase platelet counts, and the mechanism was related to the polarization of macrophages toward the anti-inflammatory M2 phenotype and the decrease in proinflammatory cytokine production; these phenomena ultimate ameliorate innate immune disorders. This study provides new insights into the mechanism by which MSCs function in the treatment of ITP.
Footnotes
Acknowledgments
We thank the Lim Por Yen Medical Research Center of Sun Yat-sen Memorial Hospital (Guangdong, China) for their equipment support and technical guidance, and also thank Saliai Stem Cell Science and Technology Co. LTD for providing hBMSCs.
Author Disclosure Statement
The authors declare that there is no conflict of interest to declare.
Funding Information
This work was supported by Science and Technology Program of Guangzhou (grant No. 201803010012).
Supplementary Material
WBCD206InFigure3d
WBCD206InFigure4c
WBgapdhInFigure3d
WBgapdhInFigure4c
WBinosInFigure3d
WBinosInFigure4c