
Abstract
The aim of this article was to investigate whether exosomes derived from bone marrow mesenchymal stem cells repair damaged endometrial stromal cells (EnSCs) through the miR-99b-5p/PCSK9 axis. Exosomes derived from bone marrow mesenchymal stem cells (BMSC-exos) were isolated by ultracentrifugation and characterized using transmission electron microscopy and nanoflow cytometry. A mifepristone-induced EnSC injury model was established in vitro, and the uptake of BMSC-exos was assessed. EnSCs were divided into three groups: the normal group (ctrl), EnSC injury group (model), and BMSC-exo treatment group. The effects of BMSC-exos on EnSC proliferation, apoptosis, and vascular endothelial growth factor (VEGF) expression were assessed by coculturing MSC-exos with endometrial cells. Furthermore, high-throughput sequencing was used to identify differentially expressed genes (DEGs). Through bioinformatics analysis, reverse transcription-quantitative polymerase chain reaction, western blotting, the CCK8 assay, immunohistochemistry, and dual-luciferase experiments, the potential mechanism by which BMSC-exos-derived miRNAs repair EnSC injury was studied. BMSC-exos expressed the marker proteins CD9 and CD63. Laser confocal microscopy showed that BMSC-exos could enter damaged EnSCs. In the BMSC-exos–EnSC coculture group compared with the model group, BMSC-exos significantly increased the proliferation of damaged EnSCs and inhibited cell apoptosis in a dose-dependent manner. The expression levels of Caspase-3, Caspase-9, Bax, and VEGF mRNA were significantly downregulated in the BMSC-exos–EnSC coculture group, whereas Bcl-2 expression was upregulated. We identified 28 overlapping DEGs between the model and ctrl groups and between the BMSC-exo and model groups. Transfection with miR-99b-5p mimics significantly decreased PCSK9 gene expression and inhibited the expression of the autophagy-related proteins Beclin-1 and LC3-II/I and apoptosis, thereby promoting EnSC proliferation. Transfection with a miR-99b-5p inhibitor showed the opposite effects. Beclin-1, LC3-II/I, and PCSK9 expression in the thin endometrium was significantly increased. miR-99b-5p promoted cell proliferation by targeting PCSK9. BMSC-exos promoted endometrial proliferation, and miR-99b-5p inhibited cell apoptosis and promoted EnSC proliferation by targeting PCSK9, providing a new target for the treatment of thin endometrium.
Introduction
Successful embryo implantation depends on the perfect synchrony of normal embryonic development and good endometrial receptivity. The endometrium is a highly regenerative tissue that undergoes cyclical morphological and biochemical changes essential for reproduction each month, primarily in response to sequential patterns of ovarian sex steroid concentrations, which are crucial to the regulation of growth and differentiation of the endometrium. 1–2 These cyclical changes represent the self-repair process of the endometrium, which involves stromal cell proliferation and angiogenesis. Abnormal repair of the endometrium can cause amenorrhea, infertility, miscarriage, or other symptoms. 3 Various therapies for endometrial repair have been developed over the past years, such as estrogen (E2), hysteroscopy for intrauterine adhesions, local scratching, aspirin administration, vaginal sildenafil citrate, and intrauterine infusion, which have been used to promote endometrial regeneration. 4 –6 However, the effectiveness of these methods is controversial from the view of evidence-based medicine. 7–8 Therefore, it is necessary to develop novel methods for endometrial repair.
In recent years, mesenchymal stem cells (MSCs), which have self-renewal, differentiation, and immunomodulatory abilities, have been widely used to treat many diseases. 9 –12 Research shows that MSCs, including bone marrow mesenchymal stem cells (BMSCs), umbilical cord mesenchymal stem cells (UMSCs), and menstrual blood-derived stem cells (MenSCs), can effectively repair endometrial damage and promote fertility recovery. 13 –16 Increasing evidence suggests that BMSCs can not only replace damaged cells through self-proliferation and differentiation but also secrete important regulatory factors, such as vascular endothelial growth factor (VEGF), insulin-like growth factor-1 (IGF-1), epidermal growth factor (EGF), and fibroblast growth factor (FGF), through the paracrine pathway, which can directly or indirectly affect the microenvironment of damaged tissue to stimulate angiogenesis and the proliferation of endogenous cells. 17
Exosomes, a major class of extracellular vesicles (EVs) secreted by various cells, mediate intercellular communication by transporting bioactive molecules, such as mRNAs, noncoding RNAs, lipids, and proteins. 18 –20 Notably, the use of exosomes derived from MSCs (MSC-exos) has been proposed as an alternative to MSC-based therapy for several diseases. 21 Numerous studies have proven that the specific miRNAs enriched in MSC-exos might account for the ability of these exosomes to repair the damaged endometrium. 22 –24 UMSC-exos can promote the proliferation of endometrial stromal cells (EnSCs) in a dose-dependent manner through the transport of miR-21-5p. 25 Wang et al. further proposed that BMSCs can increase the endometrial thickness and normalize protein expression in a rat model of thin endometrium, possibly through the negative regulation of HOXA10 by miR-196b, to further promote the expression of VEGF, leukemia inhibitory factor (LIF), and integrin. 26 In addition, some studies have found that BMSCs can ameliorate endometrial fibrosis by transporting exosomal miR-340 to EnSCs and promote the proliferation of vascular endothelial cells to repair the damaged endometrium, 27 but the specific mechanism underlying this effect has not been clarified.
Mifepristone is a progesterone receptor antagonist, which has potential therapeutic effects on many diseases, such as hysteromyoma, endometriosis, and functional uterine bleeding. 28 For many years, research has found 29 –31 that mifepristone has a significant antiproliferative effect and is often used in research on endometrial cell damage. In this study, to further explore the mechanisms by which BMSC-exos exert a therapeutic effect, BMSC-exos were isolated and assessed to ameliorate endometrial injury on the basis of a mifepristone-induced hEnSC injury model. The results showed that overexpression of miR-99b-5p in BMSC-exos promoted endometrial cell proliferation. In addition, our results demonstrated that miR-99b-5p exerted its effect by directly inhibiting PCSK9-mediated apoptosis pathway, identifying a novel therapeutic strategy for endometrial injury.
Methods
BMSC culture
BMSCs were provided by the American Type Culture Collection (Cat. No. CP-H166) and cultured in complete mesenchymal stem cell medium (Procell, CA) supplemented with 1% penicillin–streptomycin (Solarbio, CA) in a 37°C incubator with 5% CO2. The medium was changed every 3 days, and when the confluency reached 80%, the adherent cells were passaged with 0.25% trypsin (Solarbio). Cells from the third passage were used for subsequent experiments.
BMSCs were identified by fluorescence staining with anti-CD29PE (BioLegend, USA), anti-CD34PE (BioLegend), anti-CD44PE-CY7 (eBioscience, USA), and anti-CD45Percp (BioLegend) antibodies at the dilution recommended by the manufacturer. The cells were incubated with each antibody for 20 min at room temperature in the dark. Then, the cells were washed twice to remove the unbound antibodies by adding 1 mL of phosphate-buffered saline (PBS), centrifuging them at 1,500 rpm for 5 min, and discarding the supernatant. The cell pellet was gently resuspended in 500 μL of PBS and then analyzed by flow cytometry (NovoCyte 2060R, Agilent, USA).
BMSC differentiation experiment
When the cells reached 100% confluency, the complete medium was replaced with adipogenic induction medium, and the cells were cultured for 3 weeks. The hBMSC lipogenic differentiation kit (Cyagen, USA) was used according to the manufacturer’s instructions. Then, Oil Red O staining was performed.
Isolation and purification of BMSC-exos
BMSCs were cultured with complete MSC medium. When the cell confluency reached 90%, the medium was changed to serum-free culture medium (Dulbecco’s modified Eagle’s medium [DMEM] + 2% exocrine-free fetal bovine serum [FBS]) for 48 h, and the supernatant was subsequently collected for the extraction of exosomes and stored at −80°C.
BMSC-exos were isolated by ultracentrifugation as previously described. In brief, the supernatant was centrifuged at 2,000g for 30 min at 4°C and 10,000g for 45 min at 4°C, filtered with a 0.45-μm filter (Millipore, USA), and ultracentrifuged twice at 100,000g for 70 min at 4°C. The supernatant was discarded, and the purified exosomes were resuspended in precooled PBS and stored at −80°C for subsequent experiments.
Characterization of BMSC-exos
The morphology of BMSC-exos was observed by transmission electron microscopy (TEM, HT-7700, Hitachi, Japan). The particle size distribution of the exosomes and the expression of surface markers (CD63, CD9) were analyzed by nanoflow cytometry (NanoFCM, N30E, CA). BMSC-exos were labeled with PKH-26 (Sigma–Aldrich, USA) according to the manufacturer’s instructions. Subsequently, PKH26-labeled exosomes were added to cultured human EnSCs and incubated for 24 h. The medium was discarded, and the cells were washed three times. The cells were fixed with 4% paraformaldehyde at room temperature for 30 min. Images were captured with a laser scanning confocal microscope (Nikon TE300, Japan).
Establishment of a human EnSC injury model in vitro
A human EnSC line was obtained from the American Type Culture Collection (Cat. No. CP-H208) and cultured in 5.5 mM glucose DMEM supplemented with 10% FBS (Gibco™, USA) and 1% penicillin–streptomycin for 48 h. Then, the medium was changed to serum-free medium for 24 h, and the cells were treated with 40 μmol/L mifepristone (Sigma, USA) for 48 h. After the treatment, the medium was changed to DMEM, and the mifepristone was removed. The treated EnSCs were used as an in vitro injury model. In the BMSC-exo treatment group, EnSC injury was modeled as described earlier, and different concentrations of exos (0, 50, 100, 150, and 200 μg/mL) were added for 48 h. Cell viability was measured using the CCK-8 assay, and apoptosis was assessed with a flow cytometer.
Analysis of cell viability by the CCK8 assay
The CCK8 assay was used to measure cell viability following the manufacturer’s instructions. The cells were seeded in 96-well plates overnight and then treated for 48 h. Cells in the control group were cultured in normal medium. The supernatant was discarded, 10 μL of CCK8 reagent was added, and the cells were incubated for 2 h. The absorbance at 450 nm was measured by a microplate reader (Thermo Fisher, USA).
Flow cytometric analysis of cell apoptosis
Cells were harvested, washed twice in PBS, and resuspended in 300 μL of precooled Annexin V-FITC binding buffer (Multiscience, CA). Then, 5 μL of Annexin V-FITC and 10 μL of PI were added to each well and incubated at room temperature for 10 min away from light. The results were analyzed with flow cytometry.
miRNA/mRNA extraction and target gene prediction
miRNA/mRNA was extracted from BMSC-exos by using an RNeasy Mini kit (Qiagen, Germany) according to the manufacturer’s protocol. The integrity and quality of the miRNA/mRNA were determined with an Agilent 2100 Bioanalyzer System (Agilent Technologies GmbH, Germany). After cluster generation, the prepared libraries were sequenced on an Illumina HiSeq platform. The sequence reads were normalized, and read quality was assessed with fastp. The reads were mapped to reference genomes using miRdeep2 software. Then, the number of miRNA/mRNA reads in each sample was counted. The DEseq2 package was used to identify differentially expressed genes (DEGs) with criteria of fold change > 2 and adjusted P value < 0.05. Gene Ontology (GO) functional enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of the target genes were performed.
Transient transfection and luciferase reporter assays
293T cells were cultured in 5.5 mM DMEM supplemented with 10% FBS and 1% penicillin–streptomycin, incubated in 5% CO2 at 37°C, and seeded at a density of 7 × 104 cells per well in 12-well plates. When the cell confluency reached 50–60%, the medium was changed to 5% FBS medium before transfection. The cells were transfected with pRL-SV40 plasmids via Lipofectamine 3000 reagent (Life Technologies) according to the manufacturer’s instructions. The miR-99b-5p and PCSK9 interaction was confirmed by luciferase reporter assay. Forty-eight hours after transfection, all transiently transfected cells were harvested, and dual-luciferase activity was measured by a fluorescence spectrophotometer. Transfection efficiency was analyzed by reverse transcription-quantitative polymerase chain reaction (RT-qPCR).
Cellular RNA extraction and RT-qPCR
The expression of Caspase-3, Caspase-9, Bax, Bcl-2, and VEGF mRNA was analyzed using RT-qPCR. Total RNA was isolated from cells from the different groups using an RNeasy Mini kit (Qiagen) according to the manufacturer’s protocol. The concentration and quality of the RNA were measured with a NanoPhotometer 80 spectrophotometer (Thermo Fisher Scientific), and the total RNA was eluted in 12 µL of RNase-free water for reverse transcription into cDNA with HiScript II qRT SuperMix (Vazyme, USA). mRNA expression was quantified by RT-qPCR using SYBR qPCR Master Mix (Vazyme). Primers for Caspase-3, Caspase-9, Bax, Bcl-2, and VEGF were obtained from General Biol, Inc. Each sample was analyzed in triplicate in three independent experiments. Predenaturation was performed at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 10 s, annealing for 30 s at 58°C, and extension for 30 s at 72°C. The primer sequences have been listed in Table 1.
The Primer Sequences used for RT-PCR
Relative gene expression levels are presented as the mean fold change in normalized gene expression relative to a calibrator sample and were analyzed according to the 2−△△Ct method.
Western blotting
For western blot analysis, proteins were extracted from EnSCs using RIPA lysis buffer supplemented with protease and phosphatase inhibitors (Applygen, CA). After centrifugation at 12,000 rpm for 10 min, the supernatant was collected for analysis. Protein concentrations were determined with a BCA protein assay kit (Elabscience, USA), and the proteins were then electrophoresed on 10% SDS–PAGE gels and transferred to polyvinylidene difluoride membranes. The membranes were blocked with 3% nonfat milk for 1 h at room temperature; incubated overnight with rabbit anti-Bax (Proteintech, USA, 1:8,000), anti-Bcl-2 (Abcam, England, 1:1,000), anti-caspase3 (Abcam, 1:2,000), anti-caspase9 (Proteintech, 1:1,000), anti-VEGF (Proteintech, 1:2,000), anti-PCSK9 (Affinity, USA, 1:1,000), anti-proliferating cell nuclear antigen (PCNA, Proteintech, 1:1,000), anti-LC3 (CST, USA, 1:1,000), and anti-Beclin-1 (Bioss, USA, 1:1,000) antibodies in blocking buffer; washed three times; and incubated with a 1:2,000 dilution of a goat anti-rabbit IgG secondary antibody (Servicebio, CA) for 2 h at room temperature. β-actin (TransGen Biotech, CA, 1:2,000) was used as the loading control. All antibodies were diluted as recommended. Protein bands were detected using the electrochemiluminescence method. Three replicates were performed for each group.
Immunohistochemistry
For immunohistochemistry, endometrial tissue was routinely obtained following standard procedures, including control group (n = 11) and thin endometrium (n = 12). The thin endometrium was diagnosed as endometrial thickness <7 mm on the mid-secretion of endometrium or the day of injection of human chorionic gonadotropin according to Chinese Expert Consensus in 2018, 32 and patients voluntarily sign an informed consent form to retain endometrial tissue samples during hysteroscopy. The content involved has been discussed by the hospital’s ethics committee. All steps are carried out according to the operation process of the immunohistochemistry kit. Briefly, fresh endometrial tissue samples were fixed in 10% formalin and embedded in paraffin. The samples were sliced into 4–5-μm sections, permeabilized with 0.5% Triton X-100, and blocked with 5% BSA. The samples were incubated with anti-Beclin-1 (Bioss), anti-LC3 (Affinity), and anti-PCSK9 (Affinity) antibodies at a 1:100 dilution at 4°C overnight. After washing with PBS, the sections were incubated with 1:100 HRP-conjugated secondary antibody (Zhongshan Jinqiao, CA) for 30 min at room temperature and were briefly counterstained for 3 min with hematoxylin solution. Images were obtained using a microscope (BX43, Olympus, Japan) at a magnification of 200×.
Statistical analysis
All experimental data are presented as the means ± standard deviations. Analysis of variance (ANOVA) followed by Student’s t test was applied for comparisons between two groups. One-way ANOVA was used to determine the significance of differences among three or more groups. All analyses were conducted with SPSS 22.0. P < 0.05 was considered to indicate a significant difference.
Results
Identification of BMSCs
Primary BMSCs were seeded in culture dishes, and the medium was changed every 48 h. When the BMSCs adhered to the walls, they were observed to exhibit a uniform fibroblast-like morphology and to be orderly arranged under phase-contrast microscopy (Fig. 1A). As time passed, the cells exhibited a more uniform and orderly radial or spiral arrangement. To evaluate the multipotency of the BMSCs, we incubated them in adipogenic induction medium. Oil red O staining showed that the cytoplasm of the cells was filled with orange–red lipid droplets of different sizes, and some lipid droplets fused to form bead-like structures (Fig. 1B). Furthermore, flow cytometric analysis showed that the cells had high expression of CD29 (99.92%) and CD44 (99.69%) and low expression of CD34 (1.08%) and CD45 (0.14%) (Fig. 1C). All of these characteristics were consistent with those of BMSCs reported in the literature.
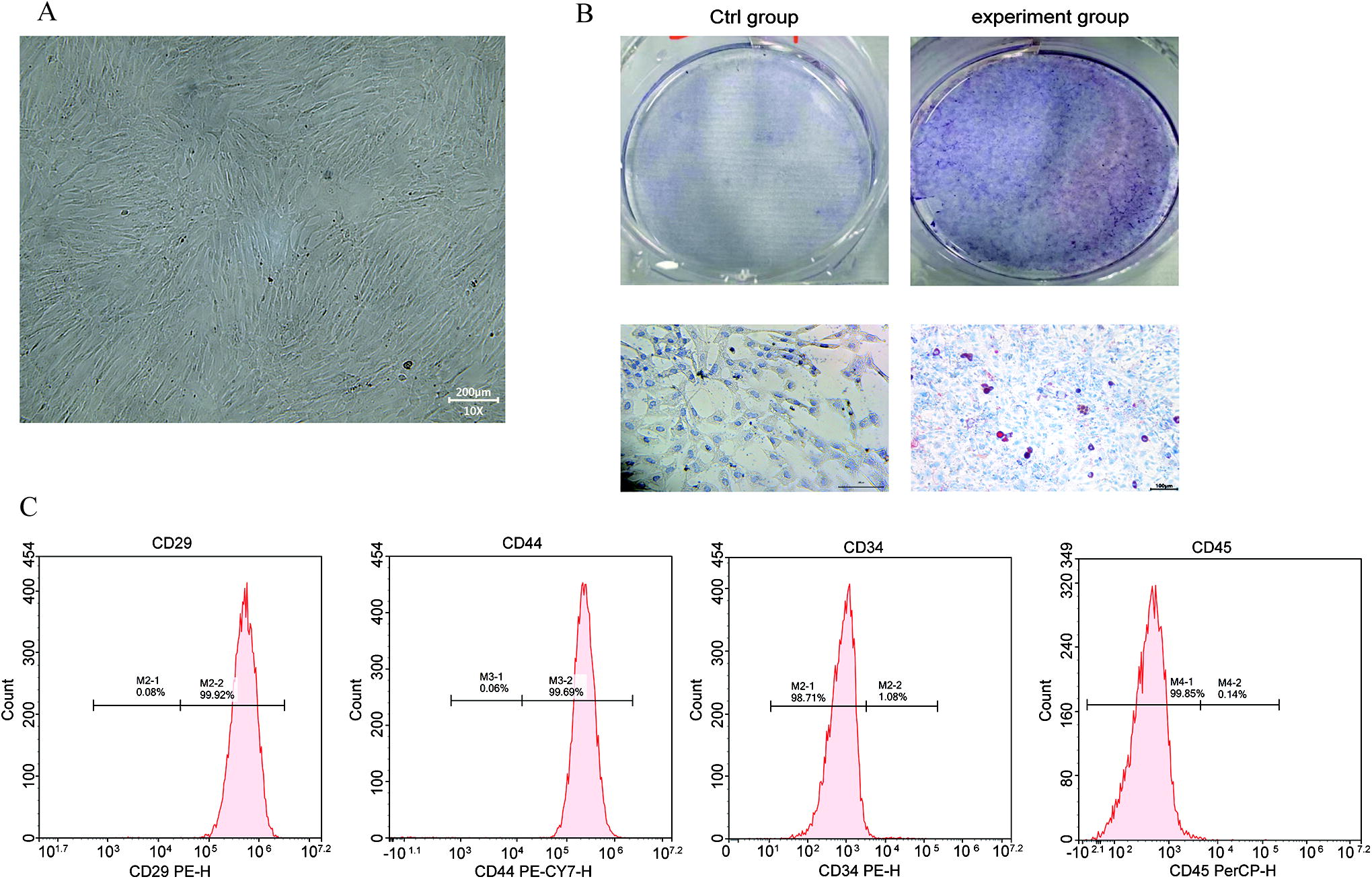
BMSC culture and characteristics.
Isolation and identification of BMSC-exos
BMSC-exos were extracted using ultracentrifugation and observed by TEM. As expected, TEM showed typical bilayer membrane vesicles containing low-electron density substances (Fig. 2A). The results of flow cytometry showed that the average diameter of exosomes was 71.63 nm (Fig. 2B), and the exosome concentration was 2.76E + 10 particles/mL (Fig. 2C). Subsequently, we found that the exosomes expressed the specific marker proteins CD9 and CD63 (Fig. 2D). To test whether the obtained exosomes exhibited biological activity, we labeled them with PKH26 and added the labeled exosomes to EnSCs. The results showed that BMSC-exos labeled with PKH26 were incorporated into EnSCs after incubation for 24 h and thus could exert their effect on EnSCs (Fig. 2E).
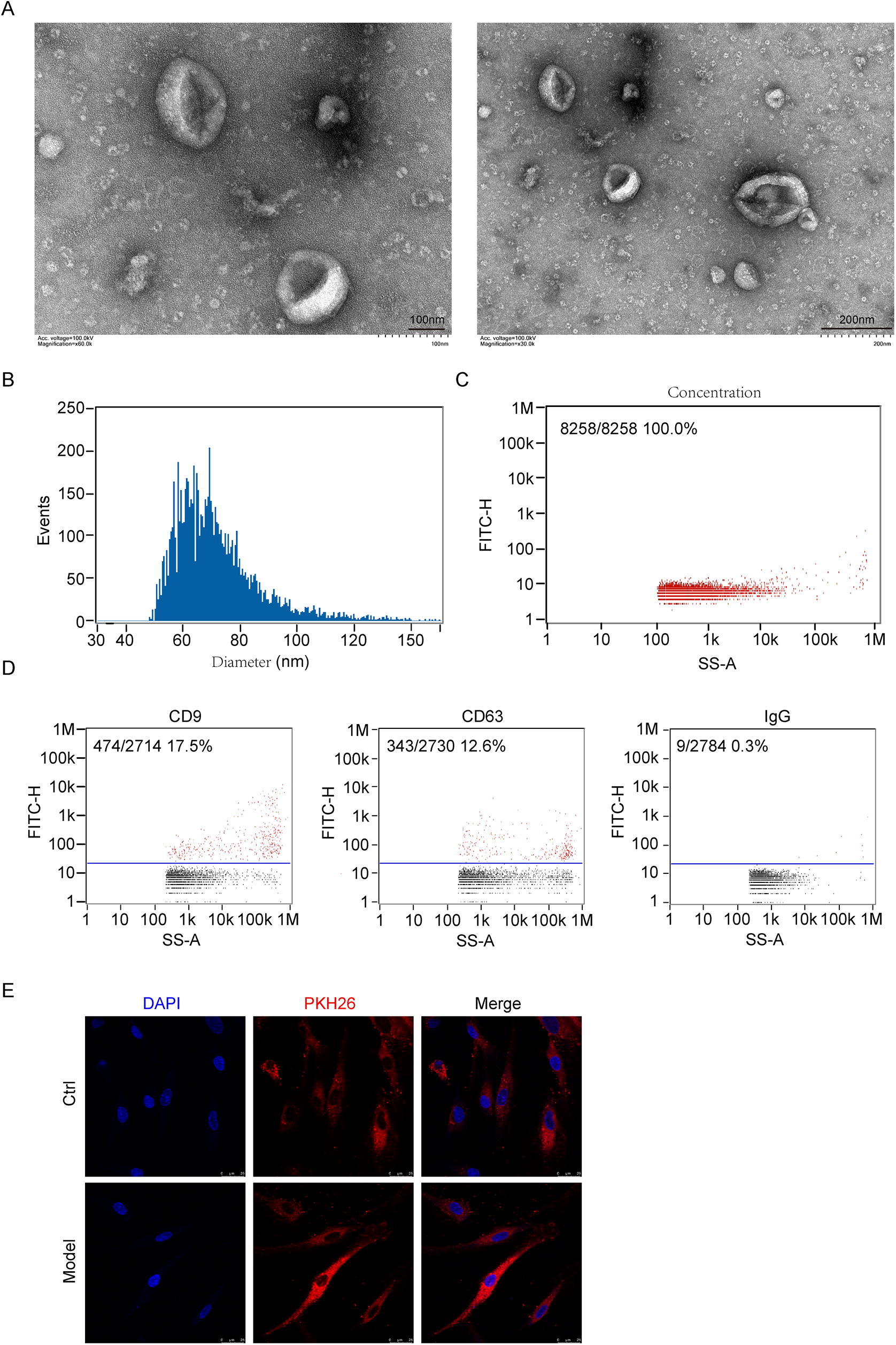
Identification of BMSC-exos.
BMSC-exos promote the proliferation of injured EnSCs
To investigate the effect of BMSC-exos on injured EnSCs, cells were divided into four groups: a group of normal EnSCs (control group), a group of EnSCs with mifepristone-induced injury (model group), a group of EnSCs with mifepristone-induced injury cocultured with BMSCs, and a group of EnSCs with mifepristone-induced injury cocultured with BMSC-exos (exosome group). CCK8 assays were performed, and the data showed that a large proportion of the cells treated with mifepristone died and that the viability was reduced by approximately 40% in the model group compared with the control group (Fig. 3A). After injured cells were cocultured with BMSCs (Fig. 3B), their proliferation significantly increased (P < 0.05). Similarly, EnSC proliferation was higher in the exos groups (86.24 ± 3.22%) than in the model group (P < 0.05) and in a dose-dependent manner (50, 100, 150, and 200 μg/mL) (Fig. 3C, P < 0.05). Furthermore, we examined whether BMSC-exos could ameliorate the apoptosis of injured EnSCs. As shown in Figure 3D, the apoptosis of EnSCs in the model group was markedly reduced after coculture with BMSCs and BMSC-exos. The mRNA expression of Caspase-3, Caspase-9, Bax, and VEGF was markedly downregulated in the exos group, and the anti-apoptosis gene Bcl-2 was upregulated (Fig. 4). The data indicated that BMSC-exos could promote the proliferation of injured EnSCs through the inhibition of apoptosis.
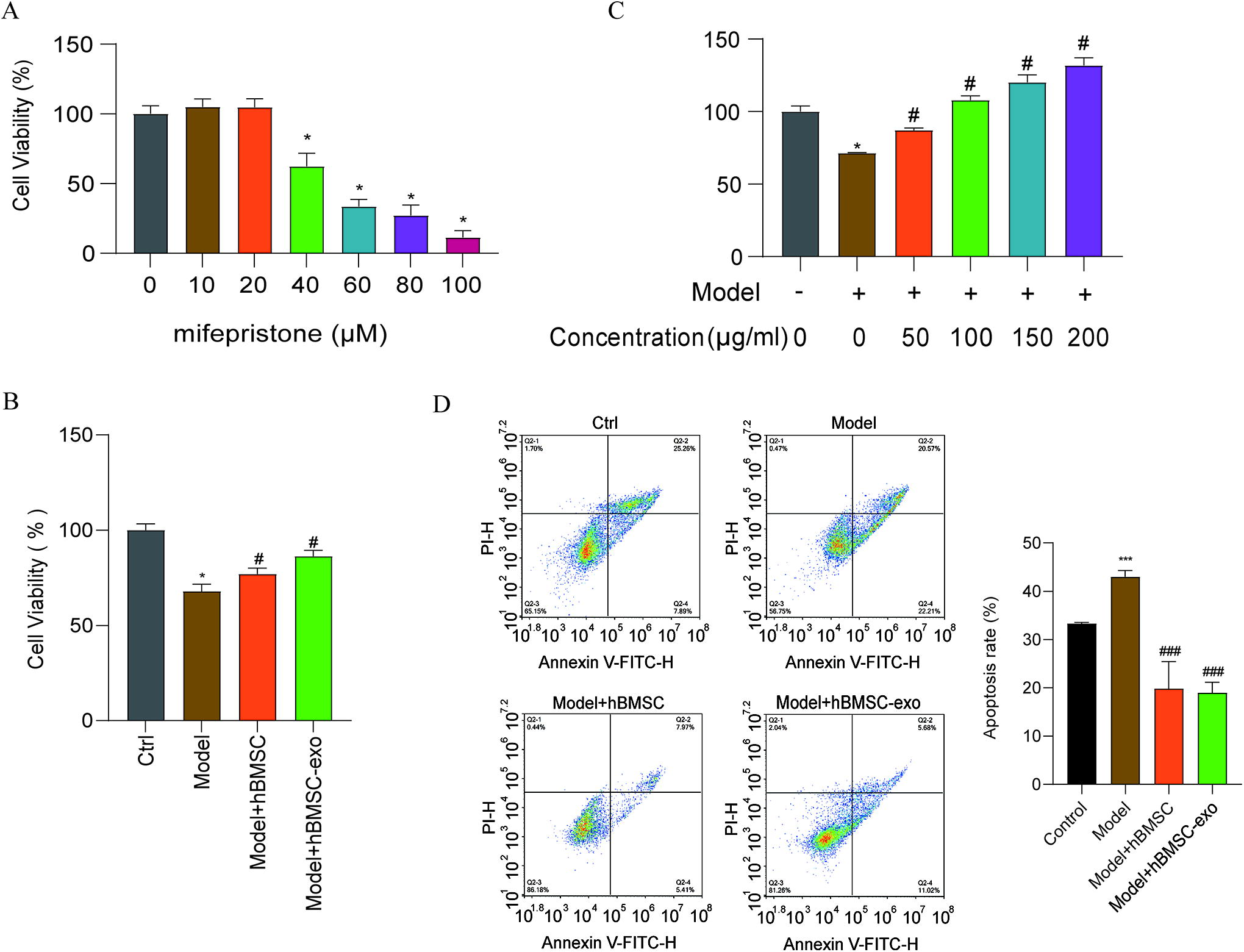
BMSC-exos promote the proliferation of injured EnSCs.

Quantitative analysis of Caspase-3, Caspase-9, Bax, Bcl-2, and VEGF mRNA expression in EnSCs. The mRNA expression of Caspase-3, Caspase-9, Bax, and VEGF was markedly downregulated in the exosome group, and the anti-apoptosis gene Bcl-2 was upregulated. *P < 0.05 compared with the ctrl group; #P < 0.05 compared with the model group. VEGF, vascular endothelial growth factor.
Differentially expressed mRNAs among the groups
To analyze mRNA expression profiles and identify the mechanism responsible for the therapeutic effects of MSC-exos, mRNAs differentially expressed in the control, model, and exos groups were identified by high-throughput sequencing. mRNAs with a |log2(fold-change) |>1 and adjusted P value < 0.05 were selected for analysis. The results showed that there were 3,749 downregulated mRNAs and 2,803 upregulated mRNAs in the model group compared with the control group (Fig. 5A). Interestingly, there were 71 downregulated mRNAs and 198 upregulated mRNAs in the exos group compared with the model group (Fig. 5B). In total, there were 144 overlapping DEGs among the three groups (Fig. 5C), and 28 of these genes (Table 2) were upregulated in the model group and downregulated after BMSC-exo treatment. Notably, some of the DEGs, including SPHK1, PCSK9, IL1RL1, MAD1L1, MT1G, and PTPRH, are associated with cell apoptosis. From the results of the analysis and a review of the literature, we selected PCSK9 and SPHK1, which are regulators of the apoptosis signaling pathway, for subsequent analysis.

High-throughput sequencing analysis assessing the role of the exosome regulatory network in endometrial repair.
Twenty-Eight DEGs for Upregulation of Model vs. Ctrl Group and Downregulation of BMSC-Exo vs. Model Group
BMSC-exo, exosomes derived from bone marrow mesenchymal stem cells; DEGs, differentially expressed genes.
According to miRNA-sequencing of exosomes derived from BMSCs, the miRNA families most strongly represented in the top 20 differentially expressed miRNAs were the hsa-let-7 family, followed by the miR-10 and miR-99 families (Fig. 5D). Further analysis showed that hsa-miR-99b-5p, hsa-let-7b-5p, hsa-miR-100-5p, and hsa-miR-423-5p interact with PCSK9, but there was no interaction between the miRNAs and SPHK1. The interaction between miR-99b-5p and PCSK9 was confirmed by miRTarBase, and the changes in the expression of these genes was validated by RT-qPCR (Fig. 5E). However, the exact mechanism by which miR-99b-5p and PCSK9 mediate the effects of BMSC-exos on EnSCs is unclear.
Exosomal miR-99b-5p promotes the proliferation of EnSCs
Mimics and an inhibitor of miR-99b-5p were generated and transfected into EnSCs; the results are shown in Figure 6A. The viability of EnSCs was significantly increased and the apoptosis of cells was decreased in the group transfected with miR-99b-5p mimics compared with the NC group. These changes were reversed after miR-99b-5p inhibitor transfection (Fig. 6B, C). In addition, the cell proliferation protein indicator PCNA and the apoptosis markers were measured with western blotting; the results revealed that PCNA was highly expressed in EnSCs transfected with miR-99b-5p mimics than in that of the mimics NC group, and Bcl-2 was upregulated, whereas Caspase-3, Caspase-9, and Bax were downregulated. The opposite changes were observed in the group transfected with the miR-99b-5p inhibitor (Fig. 7).
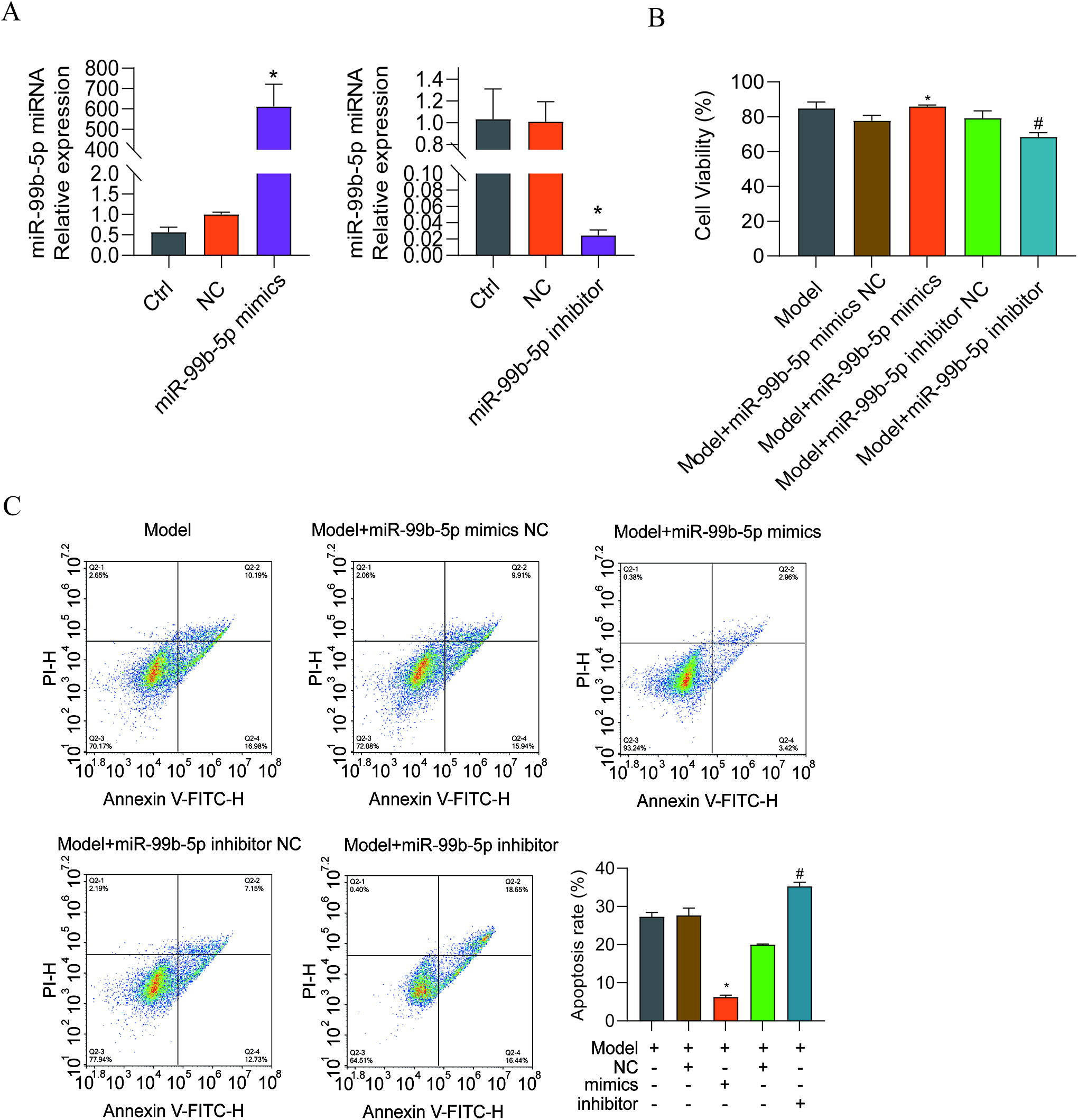
miR-99b-5p mimic and inhibitor transfection into EnSCs.

Expression of PCNA and apoptosis-related proteins after miR-99b-5p mimic and inhibitor transfection. *P < 0.05 compared with the NC mimic group; #P < 0.05 compared with the NC inhibitor group. PCNA, proliferating cell nuclear antigen.
PCSK9 promotes cell proliferation of injured EnSCs
Subsequently, to determine the function of PCSK9 in the repair of damaged EnSCs, PCSK9 was overexpressed or knocked down with small interfering RNAs (siRNAs), and the effects were assessed by RT-qPCR and western blotting (Fig. 8A). Overexpression of PCSK9 significantly increased the apoptosis of EnSCs; in contrast, apoptosis of EnSCs was decreased when the cells were transfected with PCSK9-siRNA (Fig. 8B, C). The expression of apoptosis-related proteins was also measured. The results suggested that when PCSK9 was knocked down; Bcl-2 was upregulated; and Caspase-3, Caspase-9, and Bax were downregulated (Fig. 9A). Moreover, the protein expression of LC3I/II and Beclin-1 was downregulated after PCSK9-siRNA transfection (Fig. 9B). However, the opposite changes were observed when PCSK9 was overexpressed.
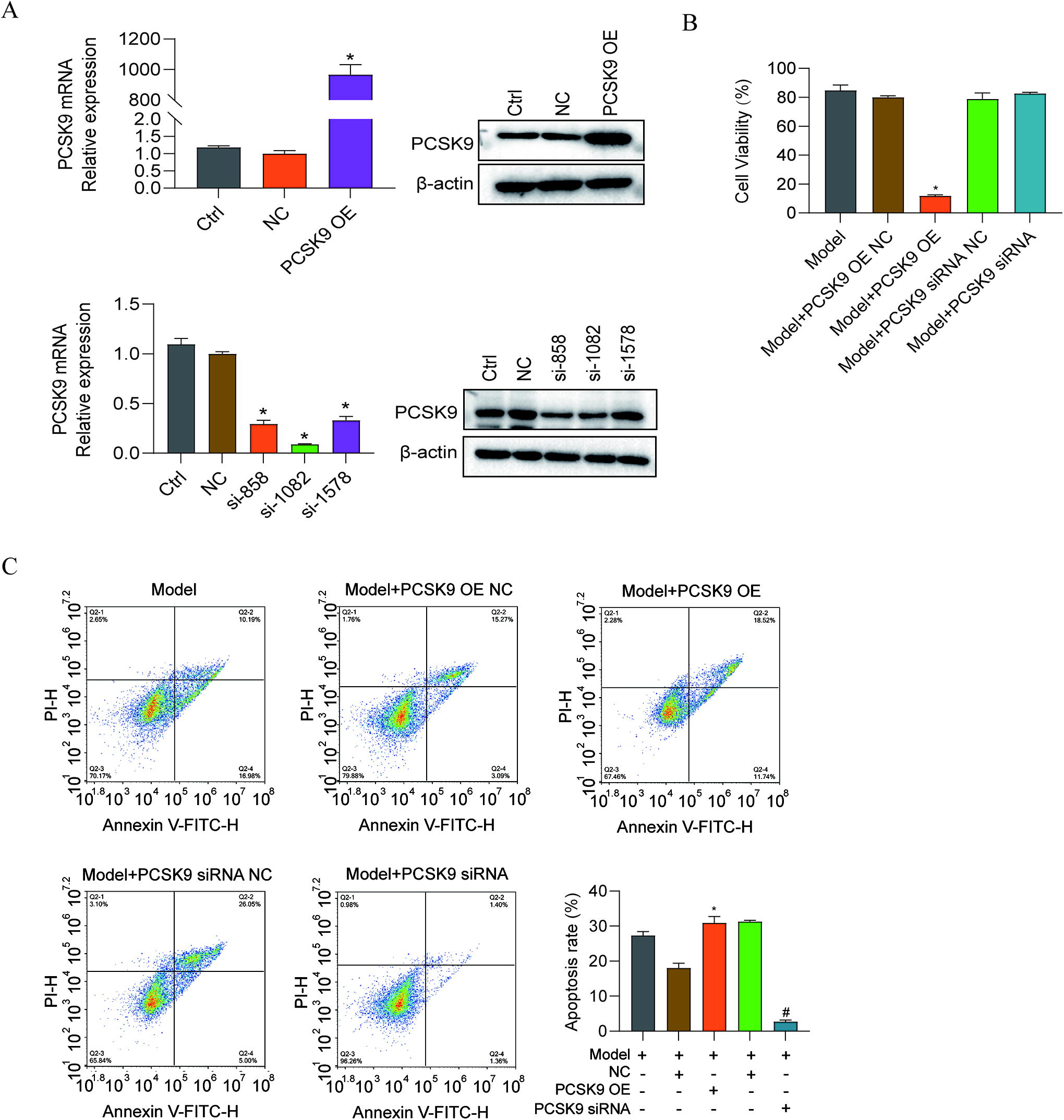
PCSK9 OE plasmid and siRNA transfection into EnSCs.

In addition, as shown in Figure 10, immunohistochemical staining revealed that endometrial injury increased the expression of LC3, Beclin-1, and PCSK9 in thin endometrium. The aforementioned results suggested that PCSK9 may act as an important promoter of cell proliferation by regulating apoptosis during injured EnSC repair.

Immunohistochemical staining was used to assess the expression of PCSK9, Beclin-1, and LC3 in the thin endometrium. Hematoxylin-stained nuclei were blue, whereas DAB staining appeared brownish yellow. From this, we found that there was stronger staining of thin endometrium than in normal cases.
Exosomal miR-99b-5p regulates the proliferation of endometrial cells by targeting PCSK9
To determine whether exosomal miR-99b-5p regulates the expression of PCSK9, a luciferase reporter assay was performed to confirm the direct interaction of miR-99b-5p and PCSK9. The 3’UTR of PCSK9, which contains the predicted miR-99b-5p binding site, was cloned into the pcDNA3.1(+) vector immediately downstream of the firefly luciferase reporter gene (PCSK9 WT). To generate a PCSK9 MUT reporter vector, the predicted miR-99b-5p binding site was mutated (Fig. 11A). The PCSK9 WT or MUT reporter vector was cotransfected with miR-99b-5p mimics, inhibitor, or NC into 293T cells. The results showed that miR-99b-5p mimics caused a significant decrease in luciferase activity, and mutation of the miR-99b-5p binding site inhibited this knockdown effect (Fig. 11B). The expression of PCSK9 was examined by RT-qPCR and western blotting after transfection with miR-99b-5p mimics or inhibitor to further verify this interaction (Fig. 11C, D).
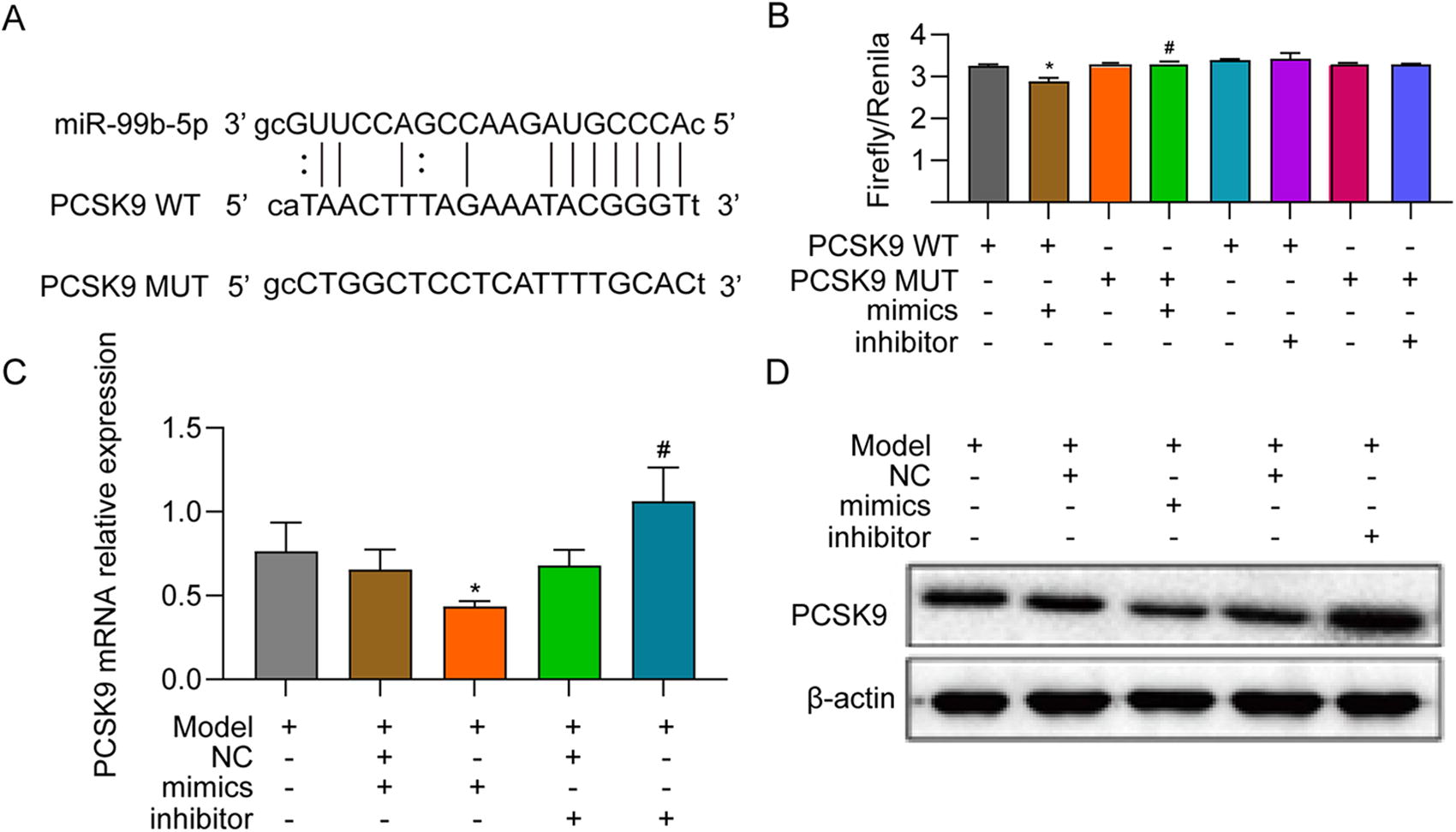
PCSK9 directly targets miR-99b-5p.
Discussion
Endometrial injury, which has a high incidence, is caused by trauma, endometritis, or congenital factors and leads to intrauterine adhesions, thin endometrium, and lower implantation and pregnancy rates. 3 The basal layer of the endometrium is an important source of cells for repair and regeneration. 1,33,34 It is widely accepted that reduced blood flow is one of the causes of endometrial injury and affects endometrial proliferation. 35 Therefore, we administered mifepristone to establish an endometrial cell injury model. Our results demonstrated, for the first time, a novel role for the miR-99b-5p/PCSK9 axis in promoting the proliferation of damaged EnSC via BMSC-exos.
Over the past decade, MSCs have been proposed to be an ideal source for cell-based therapy to promote the scar-free healing of the damaged endometrium. 36 Initially, MSCs were thought to exert their effects by their multipotent differentiation capacity and direct intercellular interactions. 12 However, emerging studies have indicated that MSCs release many EVs, which participate in tissue regeneration by transmitting information to damaged cells or tissues and exhibit biological activities similar to those of MSCs. 37
Exosomes are cell-derived microvesicles with a lipid bilayer membrane that are released from cells into the extracellular space, where they play key roles in intercellular signaling. The diameter of exosomes is approximately 50–100 nm. According to the cell source, exosomes can contain a variety of cell components, including mRNAs, miRNAs, lipids, metabolites, and cell surface proteins. 18 Since the therapeutic potential of exosomes secreted by MSCs was first described in a mouse myocardial ischemia/reperfusion injury model, 38 the therapeutic effects of MSC-exos have been further researched. In our study, after mifepristone administration, EnSC proliferation was inhibited and apoptosis was induced in vitro, and the expression of cleaved Ccaspase-3, Caspase 9, and Bax increased. In contrast, BMSC-exos increased damaged EnSC viability and growth in a dose-dependent manner and inhibited apoptosis by upregulating Bcl-2, which was consistent with the findings of several previous studies. 39
Numerous studies have confirmed that exosomal miRNAs play an important role in the pathogenesis of endometrial injury and can be used as targets for the diagnosis and treatment of this condition. 22 In the present study, through bioinformatics analysis and luciferase reporter assays, we found that miR-99b-5p directly interacts with PCSK9, which was originally found to be associated with neuronal apoptosis. 40 In addition, PCSK9 was subsequently found to be related to autophagy. Consistent with this, we demonstrated that when PCSK9 was knocked down, the expression of autophagy-related biomarkers, such as Beclin1 and LC3, and apoptosis of cells were decreased, while cell proliferation was induced. In addition, we obtained similar results with an inhibitor of miR-99b-5p. Overall, our study suggests that BMSC-exos increase PCSK9 expression and promote the proliferation of damaged EnSCs. Furthermore, these changes are accompanied by the activation of the PCSK9/miR-99b-5p axis, which inhibits damaged EnSC apoptosis and autophagy.
However, our present study does not exclude the possible influence of other substances in BMSC-exos, such as other miRNAs, lncRNAs, and proteins, in promoting endometrial cell proliferation. According to GO and KEGG analyses, the target genes of the identified miRNAs are associated with many pathways related to the occurrence and development of endometrial injury and repair. Therefore, more studies are needed to further explore the exact mechanism by which BMSC-exos promote endometrial cell proliferation.
In conclusion, BMSC-derived exosomes effectively increase the viability of injured EnSCs and inhibit apoptosis induced by mifepristone. The potential mechanism involves the regulation of the PCSK9/miR-99b-5p axis. Our results imply that the application of BMSC-derived exosomes may be a novel cell-free therapeutic strategy to promote endometrial function in patients with endometrium-related diseases.
Conclusions
In conclusion, the current study demonstrated that BMSC-exos promote EnSC proliferation and inhibit cell apoptosis. Furthermore, miR-99b-5p inhibits the expression of the PCSK9 gene, cell apoptosis, and autophagy (Fig. 12). Our results provide a target for the development of EV-related drug delivery systems for the treatment of endometrial injury-related diseases.

Schematic illustration of the mechanism of BMSC-exos regulating the proliferation of endometrial cells.
Footnotes
Acknowledgments
The authors thank the study group for their assistance with collecting patient data and endometrial biopsies. The authors express their gratitude to anonymous reviewers for the suggestive comments, which help improve this presentation.
Ethical Approval
This research study was approved by the Institutional Ethics Committee of The First Hospital of Lanzhou University (Grant No. LDYYLL2021-316).
Consent to Participate
Written informed consent was obtained from all individual participants included in the study.
Consent for Publication
Patients signed informed consent forms allowing the publication of photographs.
Author Disclosure Statement
None of the authors have relevant financial or nonfinancial interests to disclose.
Funding Information
The study was supported by the Natural Science Foundation of Gansu Province (No. 21JR7RA366), Gansu Provincial Administration of Traditional Chinese Medicine (GZKP-2021-29), and Lanzhou Chengguan District Science and Technology Project (No. 2021-9-8).