
Abstract
Intrauterine growth restriction (IUGR) pathophysiology is driven by abnormal uterine natural killer cell (uNK) activity leading to placental dysfunction. Transamniotic stem cell therapy (TRASCET) with mesenchymal stem cells (MSCs) can improve experimental IUGR by mechanisms not fully understood. We sought to examine TRASCET’s effects in downstream products of uNKs in a model of IUGR: 15 Sprague–Dawley dams were exposed to alternating hypoxia (10.5% O2) from gestational day 15 (E15) until term (E21). Their fetuses (n = 189) were divided into four groups. One group remained untreated (n = 52), whereas three groups received volume-matched intraamniotic injections of either saline (sham, n = 44) or a suspension of amniotic fluid-derived MSCs, either in their native state (TRASCET, n = 50) or “primed” to an enhanced antiinflammatory phenotype (TRASCET-Primed, n = 43). Normal fetuses served as controls (n = 33). At term, various analyses were performed, including ELISA for surrogates of placental inflammation and uNK activity. Statistical comparisons included Bonferroni-adjusted criterion. Overall survival from hypoxia was 74% (140/189). Placental efficiency was lower in untreated and sham but normalized in both TRASCET groups (P < 0.01–0.47). Interleukin-17, a stimulator of uNKs, was elevated from normal in all groups (P < 0.01 for all). Interferon-gamma, released from activated uNKs, was elevated in all groups except sham but lower than the untreated in both TRASCET groups (P ≤ 0.01–0.06). Tumor necrosis factor-alpha, also produced by uNKs, was elevated in untreated and sham (P < 0.01 for both), but normalized by TRASCET (P = 0.05) and even lowered from normal in TRASCET-Primed (P < 0.01). Vascular endothelial growth factor, also released by uNKs, was elevated in untreated and sham but lower than normal in both TRASCET groups (P < 0.01 for all). We conclude that TRASCET with MSCs modulates the activity of placental uNKs in experimental IUGR, with distinct effects on their downstream products. This mechanistic insight may inform the development of novel strategies for the management of this disease.
Introduction
Intrauterine growth restriction (IUGR) is a prevalent disease, affecting up to 8% of pregnancies and still a leading cause of perinatal mortality, second only to prematurity. 1 Survivors carry an increased risk of many long-term if not permanent consequences such as cognitive delay, obesity, and cardiopulmonary deficits. 2 –5 Currently there is no treatment for IUGR other than early delivery, which often has a negative compounding affect with prematurity.
We have recently shown that transamniotic stem cell therapy (TRASCET) with mesenchymal stem cells (MSCs) derived from the amniotic fluid (afMSCs) or placenta (pMSCs) ameliorates several manifestations of experimental IUGR, constituting an entirely novel strategy to manage this disease. 6 –9 We have also shown that, after TRASCET, donor MSCs home robustly to the placenta, which is the focal point of disease in IUGR. 6,10 There has been some indication that the effects of MSCs in IUGR relate to these cells’ known anti-inflammatory properties. 6 –9,11 However, much remains to be understood about the mechanisms behind the beneficial impact of MSC-based TRASCET in IUGR.
In this study, we sought to continue to unveil some of these mechanisms. Although we have previously uncovered the involvement of select markers of placental inflammation in this setting, 8 here we examine for the first time the chief upstream modulator of uterine natural killer cells (uNKs) while also adding comprehensive comparisons between the effects of primed versus non-primed donor MSCs.
Materials and Methods
Donor cell procurement and processing
Before injection, donor amniotic fluid-derived MSCs (afMSCs) were obtained from healthy syngeneic Lewis rat dams and banked as we have previously described. 12 –15 In brief, a midline laparotomy was performed just before term on gestational day 21 (E21, term = E21–23), exposing the bicornate uterus. Appropriate amniotic fluid pockets were identified, usually on the ventral aspect of the fetus, and amniotic fluid samples were procured via a 30G needle on a 1 mL syringe (both from Becton Dickinson, Franklin Lakes, NJ). Isolated afMSCs were expanded in culture and their mesenchymal progenitor identity was confirmed by fluorescence-activated cell sorting analysis using a Vantage SE cell sorter (Becton Dickinson). Unconjugated mouse monoclonal antibodies validated for use in rats were used, as follows (listed with their respective secondary antibodies): CD29-APC/Cy7 anti-mouse/rat CD29 antibody (catalogue # 102226, Biolegend, San Diego, CA); CD44-monoclonal anti-rat CD44-phycoerythrin (catalogue # FAB6577P, R&D Systems, Minneapolis, MN); CD45-PE/Cy7 anti-rat CD45 antibody (catalogue # 202214, Biolegend); CD90-mouse anti-rat-FITC (catalogue # 554894, BD Biosciences, Franklin Lakes, NJ); and CD73-mouse anti-rat CD73 (catalogue # 551123, BD Pharmingen, Franklin Lakes, NJ) conjugated with a biotin anti-mouse IgG1 (catalogue # 406604, Biolegend). Nonspecific cell staining was excluded with mouse isotype immunoglobulin controls. All donor cells were labeled with green fluorescent protein (GFP) using plasmid pMIG #9044 (Addgene, Cambridge, MA) and Lipofectamine LTX with PLUS Reagent (Invitrogen, Grand Island NY). This was done by transfection in antibiotic-free mesenchymal media [450 mL DMEM (Thermo Fisher Scientific, Waltham, MA) + 50 mL FBS (Life Technologies, Chicago, IL)] for 6 h at 37°C, and then refed with fresh M-20 media during expansion thereafter. An EVOS® FL Color Imaging System microscope (Life Technologies) was used to confirm GFP positivity. Of note, as mentioned above, the donor cells used had been previously banked. Before storage, they were labeled as so described; therefore, the said labeling details are described here in the name of accuracy. However, donor cells were not traced in the present study as such analysis has already been performed in a previous study from our group on this same rat model. 6
Donor cell priming
A portion of the donor afMSCs were primed toward an enhanced antiinflammatory phenotype by exposure to a modified culture media consisting of the standard media added with 30 ng/mL of interferon-gamma (IFNγ) and 5 ng/mL of interleukin-1beta (IL-1β) (both from PeproTech, Cranbury, NJ) for 24 h before injection as we and others have previously described, with dosing variations. 6,16,17 Cells were washed twice after cytokine exposure before in vivo injection to prevent inadvertent transfer of the said cytokines to the amniotic fluid.
IUGR model and intraamniotic injections
Fifteen age-matched time-dated pregnant Sprague–Dawley dams (Charles River Laboratories, Inc., Wilmington, MA) were submitted to an IUGR model using hypoxia based on methods as previously described. 6,18 Briefly, in their final week of pregnancy starting on E15 (term: E21–22), dams were placed in a hypoxia chamber (Pro Ox P360, Biospherix, Parish, NY) configured to alternating cycles of 10.5% O2 every 12 h. Hypoxia was scheduled for daylight hours, whereas normoxia occurred overnight during their more active period. Dams were subsequently divided into four groups on E17: an untreated group had no further manipulations (n = 52 fetuses), whereas three additional groups received volume-matched (50 μL) intraamniotic injections of either saline (sham; n = 44) or a suspension of either regular afMSCs (TRASCET; n = 50) or primed afMSCs (TRASCET-Primed; n = 43). All injected cells were administered as a suspension of 2 × 106 cells/mL in PBS and all fetuses originating from each dam received the same treatment. Both the cell concentration and volume of the suspension injected as described had been previously optimized. 19
To perform the intra-amniotic injections, dams were removed from the hypoxia chamber for less than 1 h, as we have previously described. 6 In brief, chamber inhaled isoflurane was used to induce anesthesia (Abbott, Abbott Park, IL) and maintained via nose cone at 2–4% in 100% O2. Dams underwent a midline laparotomy, allowing exteriorization of the bicornate uterus and inspection of the number of fetuses. Injections were then performed under direct visualization in order to prevent damage to the fetus, placenta, or umbilical cord. A 33G noncoring needle fitted to a 100-μL syringe (both from Hamilton Company, Reno, NV) was used for each injection. After completion of the injections, the uterus was returned to the abdomen and the laparotomy was closed in two layers with 3-0 Vicryl (Ethicon, Somerville, NJ) and 5-0 Monocryl (Ethicon) running sutures. Animals were allowed to briefly recover and observed for signs of distress before returning to the hypoxia chamber. Postoperative analgesia was provided with sustained release buprenorphine (Zoopharm, Windsor, CO).
Gross measurements and uteroplacental tissue extraction
Just before term, on E21, all dams underwent euthanasia via CO2 inhalation. The previous midline laparotomy was reopened and the uterus eviscerated. All subsequent dissection was done under 2.5× loupe magnification as previously described. 20 Briefly, a uterine myotomy was performed and fetuses were delivered, the amniotic sac was then disrupted and removed, and the fetuses were dried. Their umbilical cords were clamped and cut to prevent bleeding, and each fetus was weighed. The placenta was trimmed from all chorionic attachments and subsequently weighed. Both fetal and placental weights were adjusted to maternal weight to account for the variance between dam sizes. 7 –9 Placental efficiency (PE), defined as the ratio between fetal and placental weight, was then calculated.
An additional 33 normal fetuses from three Sprague–Dawley dams not exposed to hypoxia were procured on E21 to serve as normal controls and undergoing the same measurements described above.
Enzyme-linked immunosorbent assay
The placentas were subsequently homogenized, standardized by weight, and submitted to enzyme-linked immunosorbent assay (ELISA) for markers of placental pathology previously described to be altered in models of IUGR, specifically IL-17, known to activate uNKs in physiological IUGR; IFN-γ and tumor necrosis factor-alpha (TNF-α), both pro-inflammatory products of cytolytic uNKs; and vascular endothelial growth factor (VEGF), a pro-angiogenic byproduct of activated uNKs. 21 –23 All ELISA kits (Abcam, Cambridge, MA) were used as per manufacturer recommendations under similar temperature and light conditions.
Statistical analysis
Fisher exact test was used to compare survival between groups. Medians and interquartile ranges were calculated for the gross morphometric data, and all ELISA biomarker data were analyzed by nonparametric Wilcoxon rank sum test. Significance was set at P ≤ 0.05 and included Bonferroni-adjusted criterion. All analyses were done using IBM SPSS software (SPSS version 21.0, IBM, Armonk, NY).
Results
Overall survival from hypoxia was 74% (140/189), but significantly lower in all three injection groups than in the untreated group (P < 0.01 for all). There was no difference in survival between sham and either TRASCET group (P = 0.99 for both), or between both TRASCET groups (P = 0.81).
Gross morphometrics
Placental efficiency (PE) was significantly lower than normal in untreated and sham groups (P < 0.02 and <0.01, respectively). However, PE normalized in both the TRASCET and TRASCET-Primed groups (P = 0.47 and 0.28, respectively, compared with normal controls) (Table 1). Both TRASCET groups had significantly improved PE compared with both sham and untreated groups (P < 0.01-0.03). There was no difference in PE between both TRASCET groups (P = 0.58).
Maternal Adjusted Placental Weight, Maternal Adjusted Fetal Weight, Placental Efficiency (All Ratios) and IFN-γ, TNF-α, VEGF, and IL-17 Levels (All pg/mg)
TRASCET, transamniotic stem cell therapy; IFN-γ, interferon-gamma; VEGF, vascular endothelial growth factor; IL, interleukin.
Maternal adjusted fetal weight was significantly lower in all hypoxic groups (untreated, sham, and both TRASCET groups) compared with normal controls (P < 0.01 for all), confirming successful replication of the IUGR model. However, both TRASCET and TRASCET-Primed groups had significantly higher maternal adjusted fetal weight than the sham group (P < 0.01 for all) (Table 1). The TRASCET-Primed group had significantly higher maternal adjusted fetal weight than the TRASCET group (P < 0.02). Maternal adjusted placental weights were comparable with normal in the untreated, sham, TRASCET, and TRASCET-Primed groups (P = 0.40–0.93).
ELISA data
Levels of IL-17 were significantly elevated in all hypoxic groups (untreated, sham, TRASCET, and TRASCET-Primed; P < 0.01 for all) (Fig. 1). There was no significant difference in IL-17 levels across the hypoxic groups (P = 0.07–0.83).
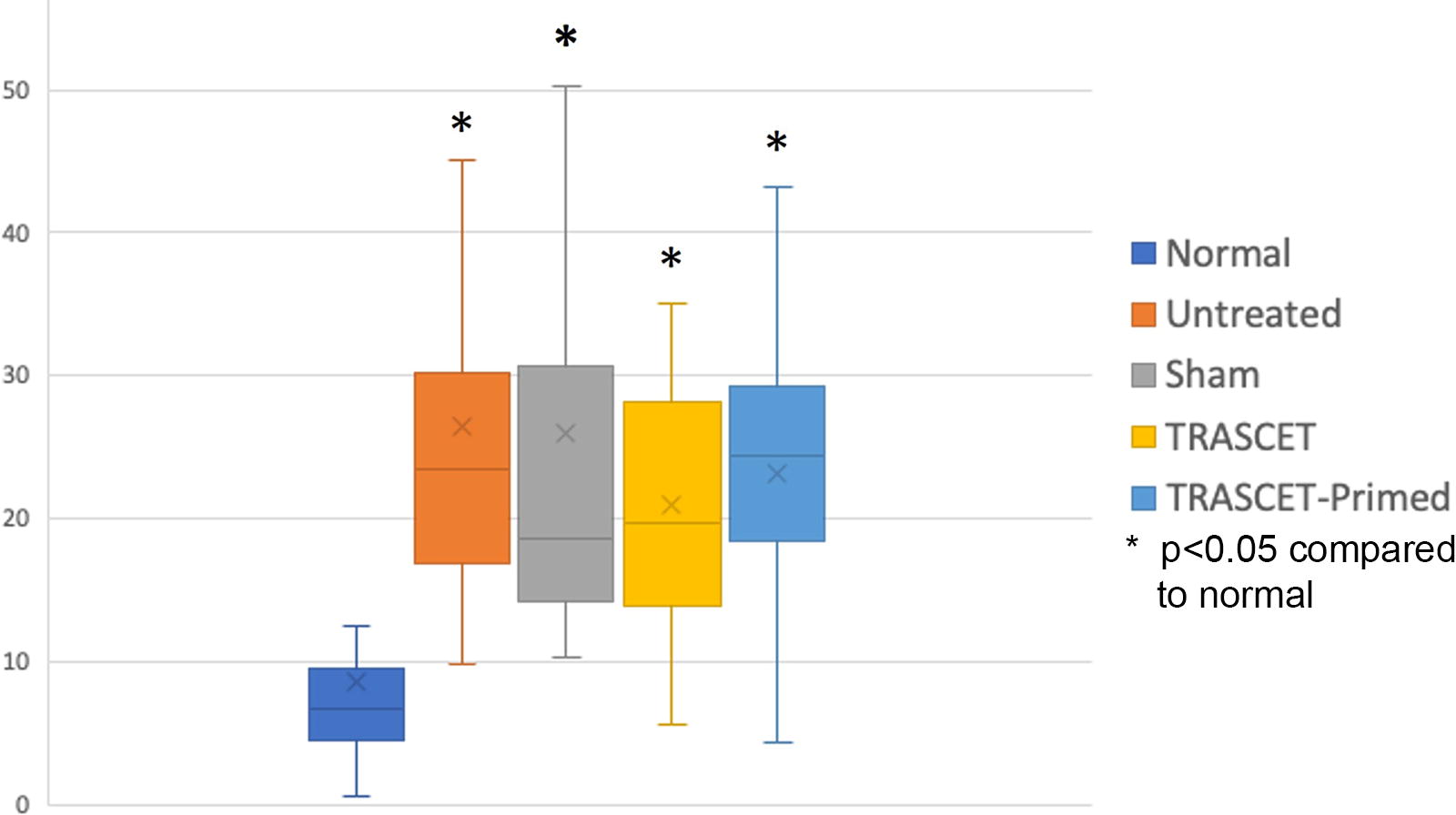
Placental IL-17 levels (pg/mg) for all groups. IL, interleukin; TRASCET, transamniotic stem cell therapy.
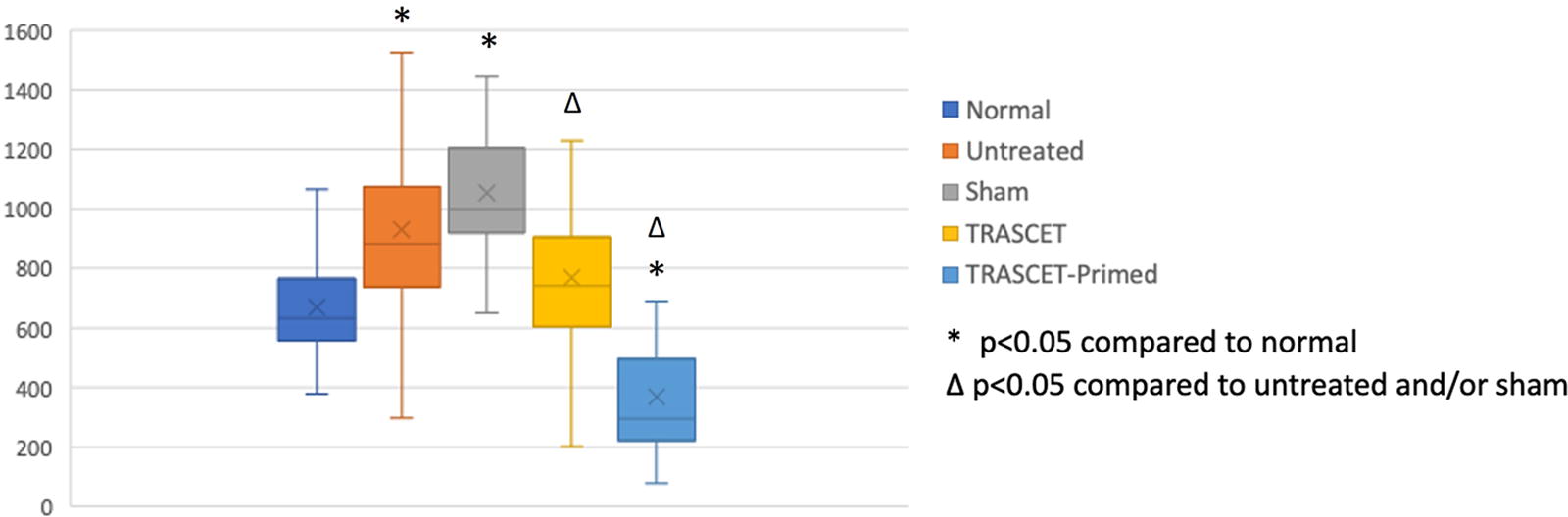
Levels of IFN-γ were significantly elevated in all hypoxic groups compared with normal controls (P < 0.01–0.02) except for the TRASCET group, in which they were normalized (P = 0.06) (Fig. 2). IFN-γ was significantly elevated in the untreated group compared with both TRASCET and TRASCET-Primed (P < 0.01 for both) and also significantly decreased in TRASCET and TRASCET-Primed group compared with sham (<0.05, 0.01). There was no difference in IFN-γ levels between the TRASCET groups (P = 0.09).
Placental IFN-γ levels (pg/mg) for all groups. IFN-γ, interferon-gamma; TRASCET, transamniotic stem cell therapy.
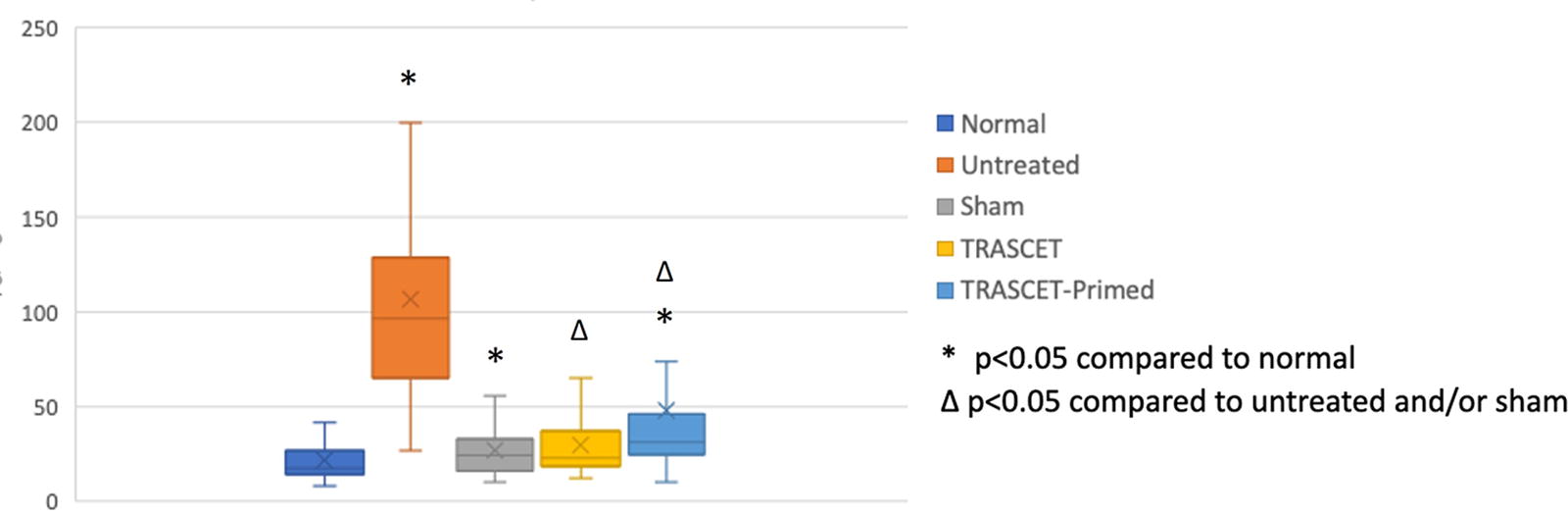
Levels of TNF-α were significantly elevated in both the untreated and sham groups (P < 0.01 for both) and normalized in the two TRASCET groups (P = 0.05) (Fig. 3). TNF-α levels were significantly below normal in the TRASCET-Primed group (P < 0.01). TRASCET-primed also had significantly lower TNF-α levels than the TRASCET group (P < 0.01).
Placental TNF-α levels (pg/mg) for all groups. TNF-α, tumor necrosis factor-alpha; TRASCET, transamniotic stem cell therapy.
Levels of VEGF were significantly elevated in the untreated and sham groups compared with normal controls (P < 0.01 for both) (Fig. 4). VEGF was actually lower than normal controls in both TRASCET and TRASCET-Primed groups (P < 0.01 for both), and TRASCET-Primed had lower VEGF levels than TRASCET (P < 0.01).

Placental VEGF levels (pg/mg) for all groups. TRASCET, transamniotic stem cell therapy.
Discussion
Although most causes of IUGR are not known and there are variations in its timing and severity, it is well established that IUGR is a placental disease. It is thought to begin with placental distress associated with hypoxia, which signals T-helper 17 (Th17) cells in the placenta to release IL-17. 23,24 IL-17 then acts as an upstream effector triggering uNKs to a proinflammatory and cytolytic state (Fig. 5). In normal pregnancies, uNKs are necessary for healthy placental remodeling and function. However, elevated numbers of these cells, especially in a cytolytic state, can damage the placenta and are associated with increased risks of spontaneous abortion and the development of IUGR. 21,23 This is because cytolytic uNKs release proinflammatory cytokines such as IFN-γ and TNF-α. 21,22 They also release increased levels of VEGF, which can lead to aberrant spiral artery remodeling, leading to poor placental circulation and growth. 24,25 The present data shows that MSC-based TRASCET impacts these downstream processes for the better.

Summarized pathophysiology of upstream signaling (IL-17) and downstream products (IFN-γ, TNF-α, and VEGF) of uterine natural killer cells (uNKs) in intrauterine growth restriction. IL, interleukin; IFN-γ, interferon-gamma; TNF-α, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor.
Our results showed that the rodent model of IUGR is associated with elevated levels of IL-17 in all hypoxia groups, a marker of placental dysfunction, and stimulator of uNK. After TRASCET, however, especially with donor afMSCs primed toward an enhanced antiinflammatory phenotype, we are able to partially reverse the fetal growth restriction ubiquitous to this model. Additionally, select markers of uNK activity known to be altered in IUGR, such as IFN-γ and TNF-α, were normalized by TRASCET. In the TRASCET-Primed group, TNF-α even reached below normal levels. This would indicate that afMSCs delivered via TRASCET appear to be modulating uNK function, given the lack of differences in the levels of their stimulator-IL-17, but significant reductions in their downstream products IFN-γ and TNF-α. This is further supported by the reduction in VEGF with TRASCET, which is also a byproduct of uNK activity. At first it may seem counterintuitive that VEGF is reduced by TRASCET, as it is associated with vascular remodeling and one might expect increased angiogenesis in the setting of placental dysfunction as an attempt to improve vascularity. 26,27 However, it is known that there is abnormal spiral artery remodeling in IUGR, and uNKs actually release VEGF in that setting. Therefore, decreases in VEGF are actually further indicative of uNK modulation.
Certainly, given the essentially unknown etiology of IUGR and the complexities of both placental function/remodeling and MSC activity in pathological sites/states, much remains to be understood about this disease and how TRASCET improves it. Just as a more immediate example, the present results could possibly have been driven, in part, also by an eventual impact of TRASCET on macrophage activity within the placenta. Macrophages can also produce IFN-γ, TNF-α, and VEGF and have been shown to play a role in IUGR. 28,29 Our main focus on uNK stemmed from the widely accepted more prominent role of these cells in IUGR, as more extensively described previously, as well as on the upstream modulatory role of IL-17 on them which, to our knowledge, has yet to be described for macrophages in this setting. At the same time, IL-17 can also activate T-cells and neutrophils, which are cell phenotypes also previously implicated in the pathophysiology of IUGR that could be playing a role in our findings on downstream products as well. 30,31 Analyses of uNK, macrophage, T-cell, and neutrophil recruitment were beyond the scope of this study. Yet, this early mechanistic inquiry into this novel treatment strategy provides some valuable insight that can guide further development, including eventual noncell-based methods of treatment. In summary, we can conclude that afMSCs delivered via TRASCET seem to modulate uNKs, which are the central drivers of placental dysfunction in IUGR, improving fetal outcomes. Such insights are also seminal to the path toward clinic translation of this novel therapy for this prevalent disease.
Footnotes
Author Disclosure Statement
No competing financial interests exist from any of the authors.
Funding Information
This work was funded by the Kevin and Kate McCarey Fund for Surgical Research at