
Abstract
With the postponement of the reproductive age of women, the difficulty of embryo implantation caused by uterine aging has become a key factor restricting fertility. However, there are few studies on protective interventions for naturally aging uteri. Although many factors cause uterine aging, such as oxidative stress (OS), inflammation, and fibrosis, their impact on uterine function manifests as reduced endometrial receptivity. This study aimed to use a combination of human umbilical cord mesenchymal stem cells (hUC-MSCs) and dehydroepiandrosterone (DHEA) to delay uterine aging. The results showed that the combined treatment of hUC-MSCs + DHEA increased the number of uterine glandular bodies and the thickness of the endometrium while inhibiting the senescence of endometrial epithelial cells. This combined treatment alleviates the expression of OS (reactive oxygen species, superoxide dismutase, and GSH-PX) and proinflammatory factors (interleukin [IL]-1, IL6, IL-18, and tumor necrosis factor-α) in the uterus, delaying the aging process. The combined treatment of hUC-MSCs + DHEA alleviated the abnormal hormone response of the endometrium, inhibited excessive accumulation and fibrosis of uterine collagen, and upregulated uterine estrogen and progesterone receptors through the PI3K/AKT/mTOR pathway. This study suggests that uterine aging can be delayed through hUC-MSCs + DHEA combination therapy, providing a new treatment method for uterine aging.
Introduction
In modern social culture, the delayed age of pregnancy for women makes maternal age the most important risk factor for successful childbirth. 1,2 Uterine aging is an important factor affecting fertility, and several age-related changes in the hypothalamus, pituitary gland, and ovaries lead to changes in the structure and function of the uterus. 3 –5 Uterine dysfunction is related to abnormal endometrial hormone response and decreased endometrial receptivity. 2,6 Multiple molecular activities may be involved in uterine aging, including oxidative stress (OS), inflammation, fibrosis, DNA damage response, and cellular aging. 7 –10 Studies have shown that changes in uterine structure are related to steroid synthesis damage. 11
Dehydroepiandrosterone (DHEA) is abundant in circulation and can be used as a precursor for endometrial cells. 12 DHEA and DHEAS levels peak in the 30s and then decrease with age, regardless of menopausal status. 13 DHEA is the main source of female androgens and research has shown its role in the immune system or improving immune response and aging. 14 Reijnen et al. found that DHEA is effective for postmenopausal women with sexual dysfunction. 15 Giannini et al. found that oral DHEA can alleviate female reproductive aging. 16 In the past decade, stem cell therapy and antiaging drugs have achieved therapeutic effects in the treatment of aging uterus but scattered evidence suggests it has not been well explored. 17 –20 In order to identify potential protective interventions for uterine aging, we used the combination therapy of human umbilical cord mesenchymal stem cells (hUC-MSCs) and DHEA in this study to explore its therapeutic effects on uterine function and structure as well as potential cellular and molecular mechanisms.
Materials and Methods
Cell culture
We purchased hUC-MSCs from Zhongsheng Traceability Biotechnology Co., Ltd (Supplementary Data S1). They were cultivated using MEM-Alpha medium (Gibco, USA) and 10% fetal bovine serum (FBS, Gibco, USA), and placed in a 37°C, 5% CO2 concentration incubator. Fibroblast-like growth was observed under a cell microscope. When the cells reached about 90% confluence, they were digested with 0.25% trypsin, separated and passaged in a 1:2 ratio, and cultured until the logarithmic growth phase is ready for injection. The cells (3 × 105) were inoculated into a 24-well plate for overnight cultivation. After treating hUC-MSCs with DHEA (1 µM) for 24 h, quantitative reverse-transcription polymerase chain reaction (qRT-PCR) was used to detect the expression level of transforming growth factor (TGF)-β and matrix metalloproteinase 9 (MMP9). Each experiment was repeated three times.
Experimental design
Eighteen-month-old C57BL/6J female mice were purchased from Beijing Huafukang Biotechnology Co., Ltd. Mice freely feed on water. Each group was randomly assigned 10 mice and the experimental groups were as follows: control, hUC-MSC, DHEA, and hUC-MSC + DHEA groups. The control group was raised normally; mice in the DHEA group freely consumed drinking water containing 0.15 mg/mL DHEA per day with a daily water intake of approximately 6–10 mL; mice in the hUC-MSC group received tail vein transplantation every 14 days (1 × 107/kg hUC-MSCs), three transplants in total; the mice in the hUC-MSC + DHEA group were injected with 1 × 107/kg hUC-MSCs in the tail vein every 14 days and transplanted three times, and freely consumed drinking water daily containing 0.15 mg/mL DHEA. All animals underwent neck dissection 60 days after treatment. The life status of mice every day was observed and recorded, and the mortality rates of each group were counted.
All procedures were approved by the Animal Protection and Use Committee of the National Family Planning Society of China. The use and operation of experimental animals comply with the relevant regulations of the Animal Experiment Management Committee of the Institute of Science and Technology of the National Health Commission.
Enzyme-linked immunosorbent assay detection of serum biochemical indicators
Mice fasted for 8 h were anesthetized with isoflurane inhalation and immediately underwent eye blood collection. Blood samples were placed at 4°C and centrifuged at 4000 rpm for 10 min to obtain serum. The level of estradiol (E2), superoxide dismutase (SOD), reactive oxygen species (ROS), and 8-hydroxydeoxyguanosine (8-OHdG), interferon-γ (IFN-γ), transforming growth factor-β (TGF-β), tumor necrosis factor-α (TNF-α), IL-1β, IL-6, and IL-18 were measured using Beijing Huabaitai Biotechnology Co., Ltd. Each experiment was repeated three times.
Immunohistochemical and immunofluorescence analysis
Multiple tumor suppressor 1 (p16) and Ki67 expression levels in the uterus along with the immunofluorescence detection of vimentin and pan-keratin expression levels were immunohistochemically detected. Rabbit anti-p16 monoclonal antibody (Abcam, Cambridge, UK; 1:200), rabbit anti-Ki67 polyclonal antibody (Abcam, Cambridge, UK; 1:200), rabbit antivimentin monoclonal antibody (Abcam, Cambridge, UK; 1:200), and rabbit anti-pan-keratin polyclonal antibody (Abcam, Cambridge, UK; 1:200) were mixed with 3% goat serum, and the mixture was added dropwise to uterine paraffin sections. The nonspecific antibody binding sites were blocked overnight at 4°C. They were incubated at room temperature for 1 h and horseradish peroxidase (HRP)-coupled goat antirabbit IgG (ZSJQ Bio, Beijing, China, 1:1000) was added. Immunoreactivity was observed using diaminobenzidine and hematoxylin staining. Image J software was used to analyze the expression level of immunohistochemistry. All immunohistochemistry and immunofluorescence experiments used the H-SCORE score. H-score = Σ (Pi × i), where Pi refers to the proportion of cells with a certain intensity in the positive cells, and i refers to the staining intensity, which usually takes values of 0, 1, 2, and 3. The staining intensity was divided into different grades: no staining (0), weak staining, medium staining, and strong staining, negative: no visible staining or only very weak background staining. Weak positive (1): the staining is light but the presence of target antigen can be clearly identified. Medium intensity (2): the staining is obvious and easy to observe. Strong positive (3): strong staining, dark and extensive color. Each experiment was repeated three times.
Mouse in vivo imaging
We added 10 μg/mL DiR with 1 × 106 hUC-MSCs and placed it at 37°C for 30 min. After washing the cells with phosphate-buffered saline, mice were injected with 1 × 106/kg hUC-MSCs in the tail vein. On the 7th day after injection, the mice were anesthetized and transferred to the NiR scanning system (Odyssey, LI-COR Biosciences) for observation. A DiR filter for excitation at 710 nm and emission at 760 nm was selected.
qRT-PCR
Extract uterine RNA using Trizol (Invitrogen, CA, USA) and reverse transcribe it into cDNA using a reverse transcription kit (Transgen Biotech, Beijing, China). The qRT-PCR detection kit (SYBR Green, Bio Rad, CA, USA) detects the expression level of the sample. Each experiment was repeated three times. qRT-PCR primer sequences:
IL-1β-Forward, 5′-ATGGGCAACCACTTACCTATTT-3′;
IL-1β-Reverse, 5′-GTTCTAGAGAGTGCTGCCTAATG-3′;
IL-6-Forward, 5′-TGTTCTCTGGGAAATCGTGG-3′;
IL-6-Reverse, 5′-CAAGTGCATCATCGTTGTTCATAC-3′;
IL-18-Forward, 5′-GTTCCCACAACGATGAGTACA-3′;
IL-18-Reverse, 5′-CTGAGGATTATAGCAGGCTTCC-3′;
insulin-like growth factor (IGF)-Forward, 5′-CTCTGCTTGCTCACCTTCAC-3′;
IGF-Reverse, 5′-CACTCATCCACAATGCCTGT-3′;
mammalian target of rapamycin (mTOR)-Forward, 5′-GGGAGAACAGAAGATGGGTAAC-3′;
mTOR-Reverse, 5′-GTTGAGAGGACCAACTGGATTAT-3′;
estrogen receptor (ER)-Forward, 5′-GCATGATGAAAGGCGGCATA-3′;
ER-Reverse, 5′-AAGGACAAGGCAGGGCTATT-3′;
progesterone receptor (PGR)-Forward, 5′-TTCATCCATGTGCCCATCCT-3′;
PGR-Reverse, 5′-GTAGGTGCAGTCTGGGAAGT-3′;
TGF-β-Forward, 5′-CAGAAGGTGGGTGGTCTTGA-3′;
TGF-β-Reverse, 5′-AGACTTTTCCCCAGACCTCG-3′;
MMP2-Forward, 5′-GCTCCACTCTTCTGGTTCTTC-3′;
MMP2-Reverse, 5′-CCCTCCTAAGCCAGTCTCTATTA-3′
Western blot analysis
The uterine protein was extracted by RIPA buffer, quantified with BCA protein detection kit (23225, Thermo Fisher Scientific, USA) and separated on 10% polyacrylamide gel. Protein transfer to the PVDF membrane was conducted using anti-p-p70S6K (Abcam, Cambridge, UK; 1:500), anti-βactin (Abcam, Cambridge, UK; 1:1000), anti-pS6K (Abcam, Cambridge, UK; 1:200), anti-IGF1 (Abcam, Cambridge, UK; 1:500), anti-ER (Biodragon, China; 1:200), anti-PGR (Abcam, Cambridge, UK; 1:200), anti-p21 (Abcam, Cambridge, UK; 1:200), anti-p53 (Abcam, Cambridge, UK; 1:200), and anti-GADPH (Biodragon, China; 1:1000) with overnight incubation at 4°C and bound to goat antirabbit IgG conjugated with HRP (ZSJQ Bio, Beijing, China; 1:2000). The protein expression was detected using an enhanced chemiluminescence assay kit (New Cell&Molecular Biotech Co., Suzhou, China). Each experiment was repeated three times. ImageJ was used to quantify the intensity of the bands. All data were based on three biological replicates.
Statistical analysis
The experimental data were analyzed using SPSS and Prism GraphPad 6.0, represented by mean ± standard deviation (mean ± SEM). The differences between the two groups of sample mean were compared using t-test. Multiple groups of data were analyzed using one-way ANOVA, with P < 0.05 being the significant difference.
Results
hUC-MSC + DHEA combined therapy improves the uterine appearance and survival rate in aging mice
We visually observed that the majority of mice uteri in the control group showed edema, suggesting that inflammation may cause edema in the uterus affecting uterine function (Fig. 1A). Weight gain is a common phenomenon in aging organisms. Therefore, we conducted a statistical analysis of the weight of mice and analyzed the changes in their weight before and after treatment (Fig. 1B and C). The results showed that compared with that in the control group (34.03 ± 0.6, n = 8; 1.18 ± 0.02, n = 8), the weight of mice in the hUC-MSC group (31.28 ± 1.4, n = 8; 1.09 ± 0.06, n = 8) and hUC-MSC + DHEA group (29.96 ± 1.1, n = 9; 1.06 ± 0.05, n = 8) decreased 60 days after treatment, with the DHEA group (33.53 ± 1.59, n = 8; 1.17 ± 0.16, n = 8) showing the most significant weight change compared with that before treatment. To test the efficacy of hUC-MSC transplantation in mice, we detected red fluorescence using in vivo imaging (Fig. 1D). The results showed that DHEA did not affect the effectiveness of hUC-MSCs. Red fluorescence of hUC-MSCs was observed in the mouse uterus, indicating that hUC-MSCs survived on a systemic scale 7 days after injection. The survival rate of aging mice showed that the lifespan of aging hUC-MSC+DHEA mice was significantly prolonged compared with that of the control; the survival rate of aging mice in the hUC-MSC group was greatly improved (Fig. 1E and F). The results showed that the combined treatment with hUC-MSCs + DHEA significantly improved uterine aging in mice and prolonged their lifespan.

The combined treatment of human umbilical cord mesenchymal stem cells (hUC-MSCs) and dehydroepiandrosterone (DHEA) improves uterine appearance and prolongs the lifespan of aging mice.
hUC-MSC + DHEA combination therapy increases the number of uterine glands and endometrial thickness
The endometrium is the site of implantation and reduced endometrial thickness can increase difficulty in pregnancy. At the same time, the number of uterine glands reflects the blood supply to the uterus. We performed hematoxylin-eosin (H&E) staining of the uterus of aging mice to observe the uterine structure and conducted a statistical analysis on the thickness of the endometrium and the number of uterine glands (Fig. 1G–1). Observation of the uterine structure revealed that the morphology of the mouse uterus remained intact. The thickness of the endometrium and the number of glands in the treatment group were greater than those in the control group. The hUC-MSC + DHEA group exhibited a more complete uterine cavity and endometrial epithelial structure. The results showed that the uterine cavity structure of the aging uterus was intact, and the number of uterine glands and the thickness of endometrium in the hUC-MSC group (21.00 ± 0.86, n = 8; 261.6 ± 13.18, n = 7), DHEA group (15.63 ± 1.03, n = 8; 242.3 ± 12.25, n = 6), and hUC-MSC + DHEA group (29.75 ± 1.52, n = 8; 303.2 ± 12.24, n = 7) were significantly increased compared with those in the control group (5.88 ± 0.69, n = 8; 196.2 ± 4.27, n = 6). The hUC-MSCs + DHEA group showed significant therapeutic effects compared with those in the control group. These results suggest that the hUC-MSC + DHEA group had an increased therapeutic effect on the structure of the aging uterus and protected endometrial receptivity.
hUC-MSC + DHEA combined therapy inhibits uterine aging
The expression of p16 and Ki67 in the uterus was detected using immunohistochemistry. The results showed that p16, which inhibits the cell cycle, was highly expressed in endometrial epithelial cells in the control (275.1, n = 8) and DHEA groups (206.3, n = 8), and positive brown staining was significantly increased compared with that in the hUC-MSC (184.9, n = 8) and hUC-MSC + DHEA groups (146.5, n = 8; Fig. 2A and B). Simultaneously, we observed that the expression of Ki67, which promotes cell proliferation, in endometrial epithelial cells in the hUC-MSC group (111.3, n = 8) and the hUC-MSC+DHEA group (123.8, n = 8) was significantly higher than that in the control group (101.4, n = 8; Fig. 2C and D). These results suggest that the vitality of endometrial epithelial cells in the aging uterus is restored by treatment with hUC-MSCs + DHEA, indicating that endometrial receptivity is protected.
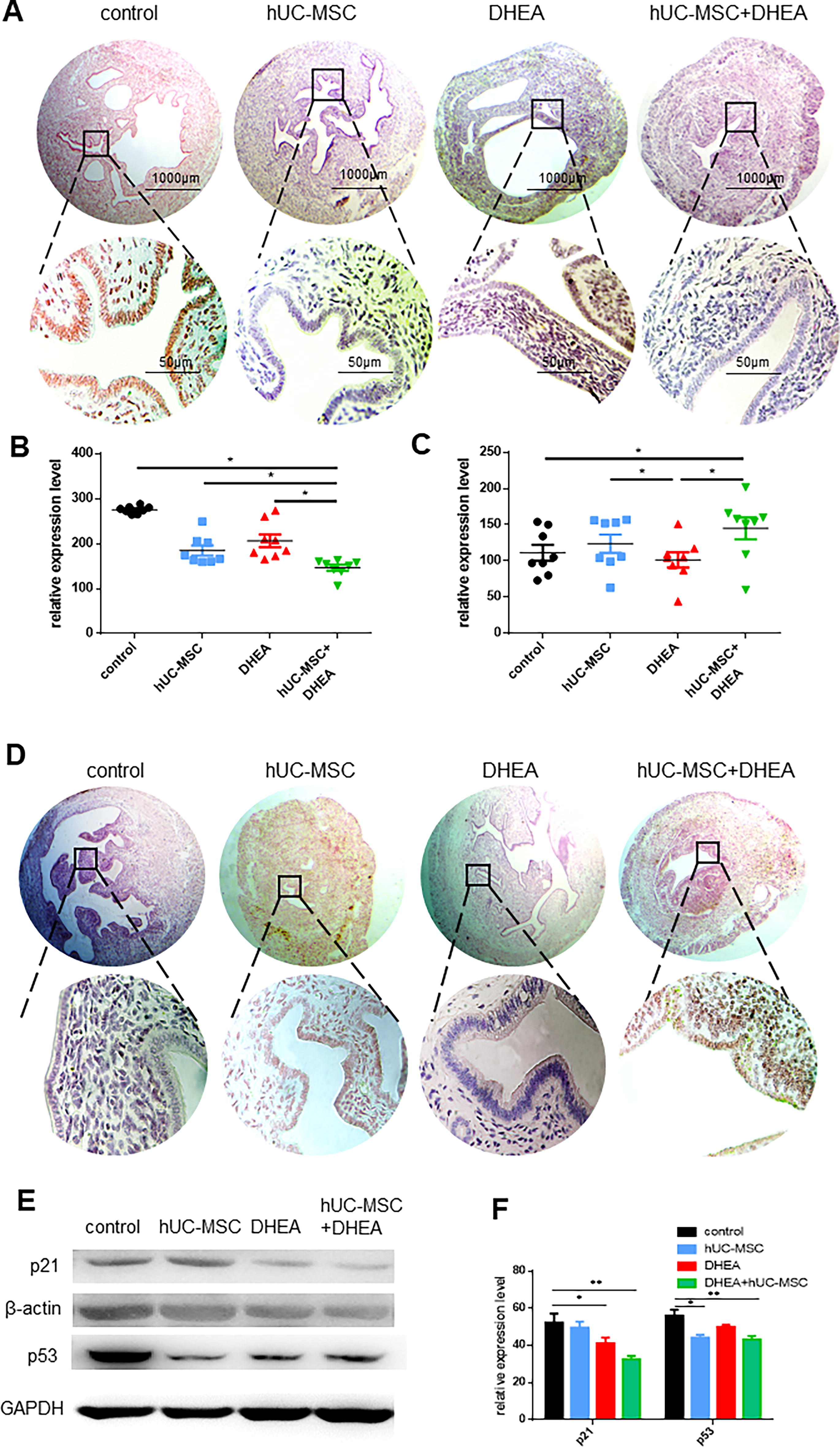
hUC-MSC + DHEA combined therapy improves uterine structure in aging mice.
We examined the expression of aging markers p16, p53, and p21 in the uterus to detect uterine aging. The results showed that the expression levels of p21 in uterine tissue were significantly reduced in the DHEA (41.33 ± 2.96, n = 3) and hUC-MSC + DHEA groups (32.67 ± 1.76, n = 3) compared with that in the control group (52.37 ± 4.81, n = 3; Fig. 2E and F). The expression level of p53 was significantly reduced in the hUC-MSC (44.32 ± 1.45, n = 3) and hUC-MSC + DHEA groups (43.43 ± 1.76, n = 3) compared with that in the control (56.5 ± 3.22, n = 3). The qRT-PCR results showed that the expression levels of p16 and p21 were significantly reduced in the hUC-MSC (0.54 ± 0.05, n = 8; 0.70 ± 0.04, n = 8) and hUC-MSC + DHEA groups (0.25 ± 0.03, n = 8; 0.56 ± 0.02, n = 8) compared with those in the control (1.00 ± 0.0, n = 8; Fig. 3A and B). These results suggest that treatment with hUC-MSC + DHEA inhibits uterine aging.
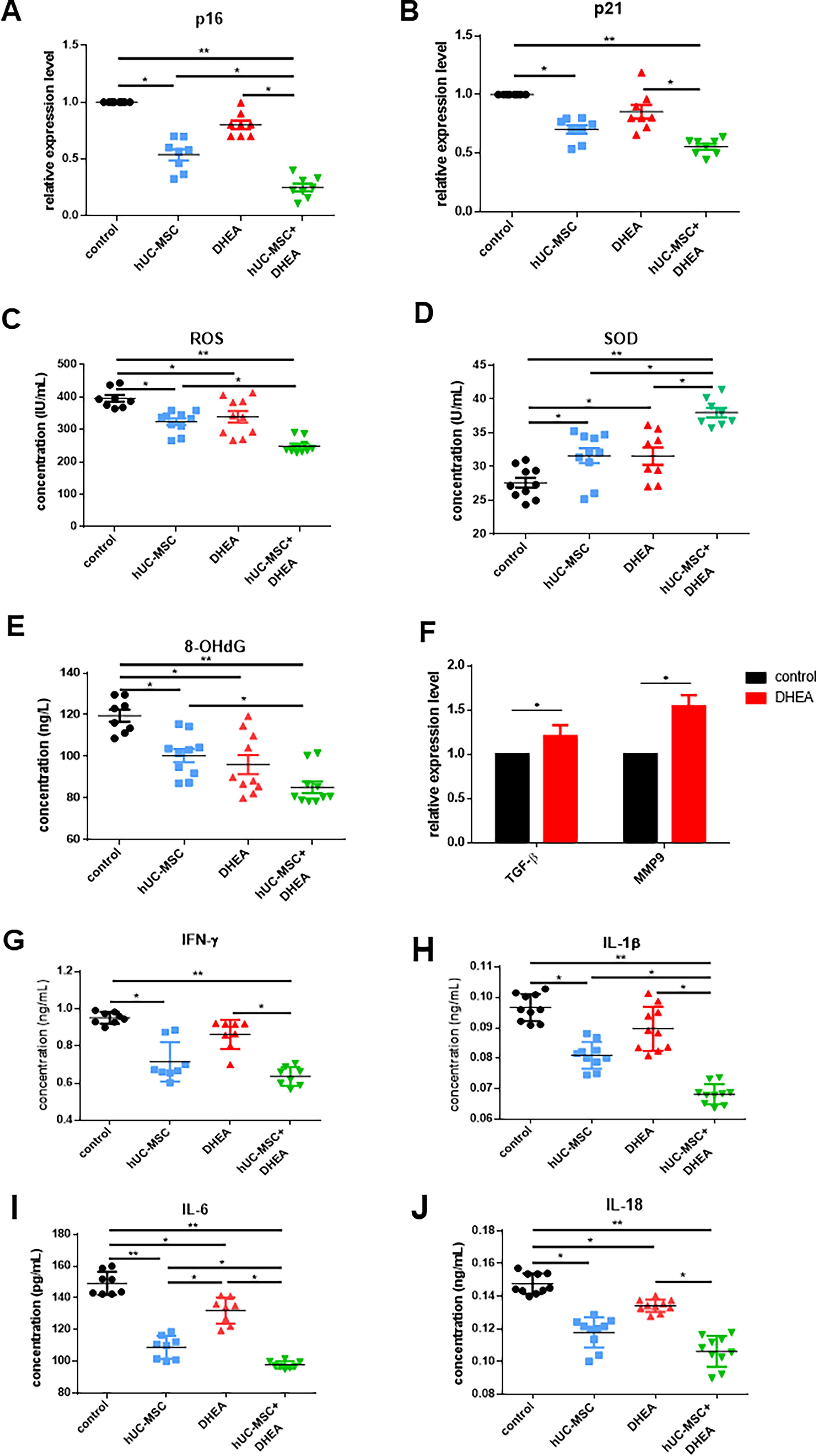
hUC-MSC + DHEA combined therapy inhibits uterine aging in mice. Detection of
hUC-MSC + DHEA combination therapy inhibits uterine proinflammatory factors and reduces OS
The accumulation of peroxides leads to body aging. We measured the OS levels in the serum of aging mice using the enzyme-linked immunosorbent assay (ELISA) (Fig. 3C–E). The results showed that the expression of ROS and 8-OHdG in the hUC-MSC + DHEA groups (248.9 ± 7.03, n = 10; 85.21 ± 2.76, n = 10; 38.53 ± 0.72, n = 8) was significantly lower than that in control (396.1 ± 10.57, n = 8; 119.5 ± 2.90, n = 8; 27.63 ± 0.73, n = 10) and hUC-MSC groups (324.1 ± 10.38, n = 10; 100.4 ± 3.18, n = 10; 31.63 ± 1.11, n = 10), whereas the expression of SOD was significantly increased. To verify the effect of DHEA on hUC-MSCs, we detected the expression levels of anti-inflammatory factor TGF-β and chemokine MMP9 in hUC-MSCs after DHEA treatment (1.21 ± 0.12, n = 3; 1.54 ± 0.13, n = 3; Fig. 3F). The results showed that DHEA increased the expression levels of TGFβ and MMP9 in hUC-MSCs, promoting hUC-MSC activity. Chronic inflammation cumulatively causes aging, an important cause of uterine collagen accumulation. We evaluated the therapeutic effect by detecting the expression levels of proinflammatory factors (IFN-γ, IL-1β, IL-6, IL-18, and TNF-α) and anti-inflammatory factors (TGF-β) in the body (Fig. 3G–J and Fig. 4A–F). The results showed that the expression of proinflammatory factors in serum and uterus in the hUC-MSC (0.71 ± 0.04, n = 8; 0.08 ± 0.00, n = 10; 108.9 ± 2.56, n = 8; 0.12 ± 0.00, n = 10; 0.56 ± 0.02, n = 8) and hUC-MSC + DHEA groups (0.64 ± 0.02, n = 8; 0.07 ± 0.00, n = 10; 97.88 ± 0.79, n = 8; 0.11 ± 0.00, n = 10; 0.50 ± 0.02, n = 8) was significantly lower than that in control (0.95 ± 0.01, n = 8; 0.09 ± 0.00, n = 10; 149.2 ± 2.59, n = 8; 0.15 ± 0.00, n = 10; 0.89 ± 0.01, n = 8); the expression level in the DHEA group (0.86 ± 0.03, n = 8; 0.09 ± 0.00, n = 10; 132.0 ± 2.92, n = 8; 0.13 ± 0.00, n = 10; 0.71 ± 0.02, n = 8) was decreased. The expression level of TGF-β was upregulated in the DHEA group (0.12 ± 0.00, n = 8) compared with that in the control (0.12 ± 0.00; n = 8) and significantly increased in the hUC-MSC (0.20 ± 0.00, n = 8) and hUC-MSC + DHEA groups (0.21 ± 0.00, n = 8). Moreover, it was observed in the hUC-MSC (0.70 ± 0.03, n = 10; 0.72 ± 0.03, n = 10; 0.80 ± 0.04, n = 10; 0.72 ± 0.02, n = 10) and hUC-MSC + DHEA (0.39 ± 0.03, n = 10; 0.41 ± 0.02, n = 10; 0.48 ± 0.04, n = 10; 0.38 ± 0.03, n = 10) groups that the expression levels of IL-1β, IL-6, IL-18, and TNF-α in serum and uterus decreased compared with that in the control (1.000 ± 0.0, n = 10); the expression level in the DHEA group (0.93 ± 0.03, n = 10; 0.97 ± 0.04, n = 10; 0.97 ± 0.07, n = 10; 0.90 ± 0.02, n = 10) decreased. These results suggest that DHEA and hUC-MSCs affect the expression of inflammatory factors and that the combined treatment with DHEA and hUC-MSCs has significant regulatory effects on the internal environment, alleviating the accumulation of collagen and the progression of fibrosis in the uterus by inhibiting pro- and enhancing anti-inflammatory factors in the uterus and serum.
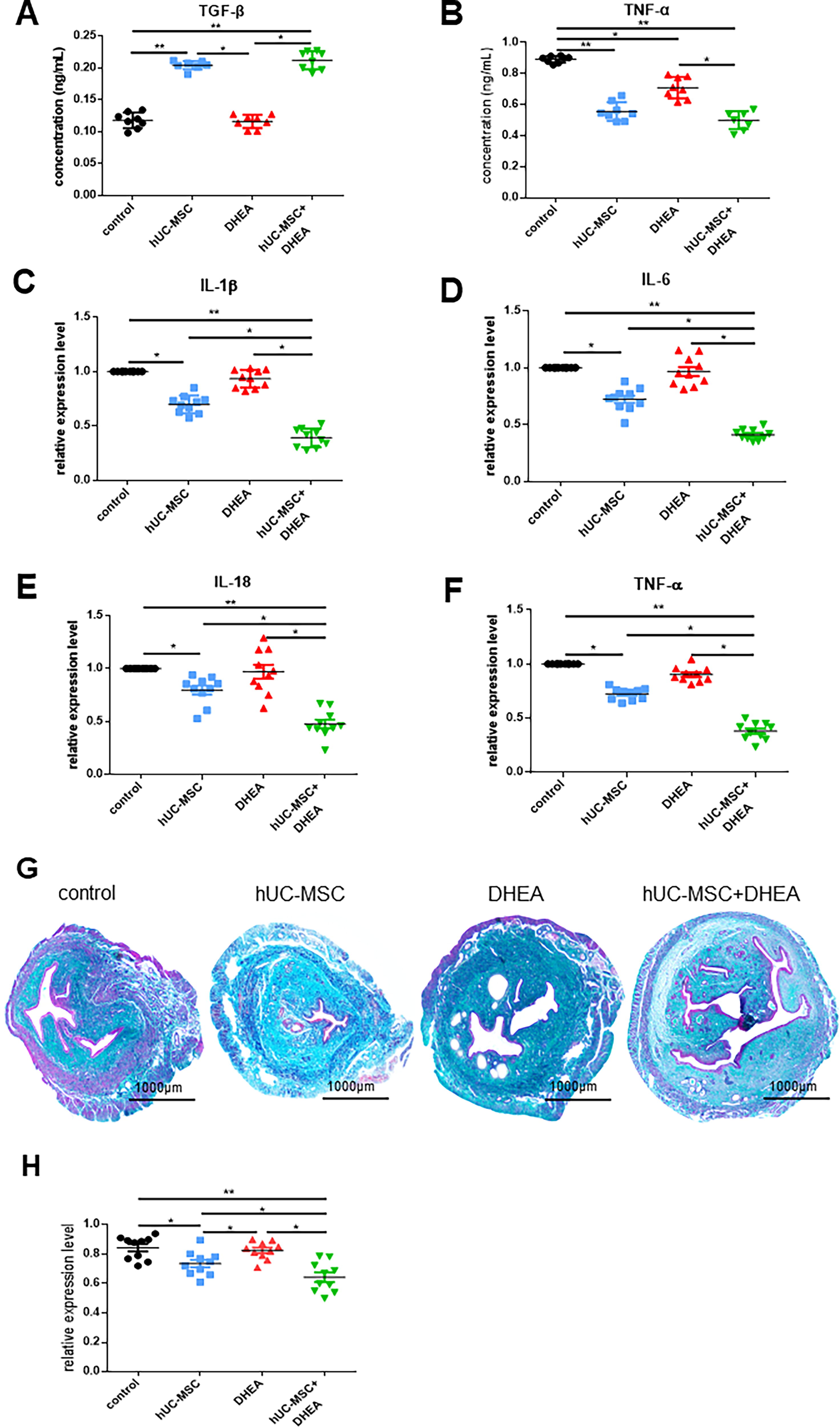
hUC-MSC + DHEA combination therapy reduces levels of inflammation. ELISA detection in mice of
hUC-MSC + DHEA combination therapy inhibits excessive collagen accumulation in the uterus
Collagen deposition is the most significant morphological change in the aging uterus. Masson’s trichrome staining was used to detect the area of green-stained collagen fibers in the uterus (Fig. 4G and H). The results showed that the amount of collagen fibers in the DHEA (0.82 ± 0.02, n = 10), hUC-MSC (0.74 ± 0.03, n = 10), and hUC-MSC + DHEA groups (0.64 ± 0.03, n = 10) was significantly lower than that in the control (0.84 ± 0.02, n = 10). Simultaneously, the quantity of collagen fibers in the hUC-MSCs + DHEA group was significantly lower than that in the hUC-MSCs and DHEA groups. These results suggest that hUC-MSCs + DHEA can inhibit the accumulation of uterine collagen fibers.
To further verify fibrosis in the aging uterus, we performed immunofluorescence staining for vimentin and keratin in the uterine tissue to observe their expression in the uterus (Fig. 5A and D). Compared with that in the control group (40.51 ± 2.65, n = 7; 64.02 ± 1.11, n = 7), the hUC-MSC (50.84 ± 1.45, n = 7; 50.71 ± 2.21, n = 7), DHEA (56.32 ± 0.88, n = 7; 55.63 ± 1.35, n = 7) and hUC-MSC + DHEA (58.49 ± 1.65, n = 7, 51.30 ± 1.09, n = 7) groups showed an increase in the expression of mesenchymal cell marker vimentin, whereas the expression of epithelial cell marker keratin decreased. The hUC-MSCs and hUC-MSCs + DHEA groups showed more significant changes compared with that in the control. These results suggest that the hUC-MSC + DHEA can maintain endometrial epithelial to mesenchymal transition.

hUC-MSC + DHEA combination therapy inhibits uterine fibrosis.
hUC-MSC + DHEA combined therapy inhibits aging through the PI3K/AKT/mTOR pathway
We evaluated uterine function recovery by measuring ER, PGR, and IGF expression levels in the uterus (Fig. 6A–C, E, and F). The qRT-PCR results showed that compared with those in the control group (1.00 ± 0.0, n = 8), the expression levels of ER, PGR, and IGF increased in the DHEA (1.57 ± 0.09, n = 8; 1.19 ± 0.10, n = 10; 1.42 ± 0.07, n = 10), hUC-MSC (1.54 ± 0.09, n = 8; 1.39 ± 0.08, n = 10; 0.95 ± 0.07, n = 10) and hUC-MSC + DHEA (1.85 ± 0.03, n = 8; 1.51 ± 0.05, n = 10; 1.25 ± 0.08, n = 10) groups, with the hUC-MSC + DHEA showing the most significant increase in expression levels. In the western blot analysis, it was shown that compared with those in the control group (43.75 ± 2.66, n = 4; 34.25 ± 2.18, n = 4; 49.75 ± 1.55, n = 4), the expression levels of PGR, IGF, and ER in the hUC-MSC + DHEA group (63.50 ± 1.71, n = 4; 58.25 ± 1.75, n = 4; 73.25 ± 1.43, n = 4) were significantly increased; the expression levels were upregulated in the hUC-MSC (41.65 ± 1.58, n = 4; 39.50 ± 1.66, n = 4; 63.25 ± 1.65, n = 4) and DHEA (48.67 ± 1.22, n = 4; 48.50 ± 3.59, n = 4; 72.75 ± 2.93, n = 4) groups. To verify the possible molecular mechanism of hUC-MSC + DHEA combination therapy in inhibiting uterine aging, we measured the expression levels of mTOR in the uterus (Fig. 6. D, G, and H). The results showed that compared with that in the control (1.00 ± 0.0, n = 10), the mTOR expression level was significantly reduced in the hUC-MSC (0.75 ± 0.03, n = 10) and the hUC-MSC + DHEA (0.52 ± 0.02, n = 10) groups. The expression levels of p-p70S6K and pS6K in the hUC-MSC + DHEA group (34.68 ± 2.67, n = 6; 13.33 ± 1.89, n = 6) were significantly reduced compared with those in the control (64.59 ± 1.45, n = 6; 47.76 ± 3.12, n = 6). The expression levels were downregulated in the hUC-MSC (50.59 ± 3.52, n = 6; 18.05 ± 1.16, n = 6) and DHEA (42.55 ± 1.01, n = 6; 25.33 ± 3.63, n = 6) groups. These results suggest that hUC-MSCs + DHEA inhibit uterine aging via the PI3K/AKT/mTOR pathway.

hUC-MSC + DHEA combined therapy inhibits the PI3K/AKT/mTOR pathway. Detection of
Discussion
The uterus is an important reproductive organ, covering the process from embryo to implantation and fetal development during pregnancy. 21 Uterine aging is associated with decreased sensitivity to hormones such as progesterone, decreased endometrial receptivity, and impaired uterine cell proliferation. 22 The effects of uterine factors on age-related infertility have attracted considerable attention. 23 Endometrial receptivity refers to the ability of an embryo to be accepted and implanted into the endometrium. 24 OS can hinder early embryo implantation and alter the normal immune function in the uterus. 25 The ways to alleviate uterine aging are still being explored. We chose to treat degenerative diseases with hUC-MSCs combined with DHEA hormone therapy, attempting to find an ideal treatment method.
In this study, we evaluated the effect of hUC-MSCs + DHEA on uterine aging by examining the appearance and structure of the uterus, the degree of uterine aging, the expression of inflammatory factors, and the degree of uterine fibrosis. The results showed that hUC-MSCs + DHEA treatment significantly improved the degree of uterine fibrosis and recovery of uterine function compared with that in hUC-MSCs or DHEA treatments alone. Through in vitro cell experiments, we demonstrated that DHEA promotes the secretion of anti-inflammatory factors, increases the secretion of chemokines in hUC-MSCs, and promotes the hUC-MSC effect. Naturally aging mice begin to experience natural death after 19 months of age. 26 We injected hUC-MSCs and tracked their distribution in aging mice using in vivo imaging. The results showed that injecting hUC-MSCs into the tail vein of mice was effective; mice treated with hUC-MSCs, DHEA, or hUC-MSCs + DHEA showed varying degrees of lifespan prolongation, with the most significant effect observed in mice treated with hUC-MSCs + DHEA. We observed the thickness of the endometrium and the number of glands through H&E staining and observed that hUC-MSC + DHEA treatment increased the thickness of the endometrium and the number of glands, indicating that the “soil” for uterine blood supply and therefore implantation will probably be restored. We used the aging markers p21 and p16 to detect the uterus of aging mice and observed that their expression levels were significantly reduced in the hUC-MSC+DHEA treatment group compared with that in the control. Immunohistochemical staining showed that the hUC-MSC + DHEA treatment significantly increased endometrial epithelial cell proliferation compared with that in the control group, reduced aging cells, and significantly increased endometrial receptive marker ER and PGR expression levels. MSCs are antioxidants that treat uterine aging and inhibit DNA damage in prematurely aging mouse models. 27 Liu et al. regenerated the cilia of the fallopian tubes in aging mice by injecting MSCs, restoring the function of the fallopian tubes and uterus. 17 Lv et al. regenerated the cilia of the fallopian tubes in aging mice by injecting MSCs, restoring the function of the fallopian tubes and uterus. 28 This suggests a combination of steroid drugs for ovarian recovery and MSCs may have better therapeutic effects on uterine aging. Our experimental results confirm this hypothesis.
During uterine aging, the accumulation of uterine collagen leads to uterine fibrosis. We used Masson’s staining to detect uterine fibrosis and an immunofluorescence assay to detect the expression of vimentin and keratin. Our results showed that hUC-MSCs + DHEA treatment significantly inhibited uterine fibrosis. Dai et al. reported that MSCs had a good therapeutic effect on uterine fibrosis in a rat injury model. 29 We detected a decrease in fibrosis in the uteri of aging mice in the hUC-MSC and hUC-MSC + DHEA groups. Through detecting inflammatory and anti-inflammatory factors, we show that hUC-MSC + DHEA treatment inhibited the accumulation of inflammatory factors in the bodies of aging mice, leading to their extended lifespans. To explore the molecular mechanism of aging, we examined the PI3K/AKT/mTOR pathway and observed that hUC-MSCs + DHEA affected aging by inhibiting the PI3K/AKT/mTOR pathway. We observed an increase in ovarian follicles in aging mice treated with DHEA, hUC-MSCs, and hUC-MSC + DHEA. This result suggests that the upregulation of hormone levels improves uterine function. Therefore, we measured ER and PGR expression levels in the mouse uterus and observed significant upregulation in the treatment group, resulting in improved uterine function in mice. These results indicate that hUC-MSCs and DHEA combination therapy provide a new therapeutic approach for delaying uterine aging.
Footnotes
Ethics Approval
The animal-related experiments, including the isolation of human umbilical cord mesenchymal stem cells and the mouse premature uterine modeling, were approved by the China of
Consent for Publication
All authors gave consent for publication.
Availability of Data and Materials
The datasets during and/or analyzed during the current study available from the corresponding author on reasonable request.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by Research fund of
Supplementary Material
Supplementary Data S1