
Abstract
The Hedgehog (Hh) signaling pathway orchestrates its influence through a dynamic interplay of Hh proteins, the cell surface receptor Ptch1, Smo, and Gli transcription factors, contributing to a myriad of developmental events. Indian Hedgehog (Ihh) and Gli zinc finger transcription factor 1 (Gli1) play crucial roles in developmental regulation within the Hh signaling pathway. Ihh regulates chondrocyte proliferation, differentiation, and bone formation, impacting the development of cranial bones, cartilage, and the temporomandibular joint (TMJ). Losing Ihh results in cranial bone malformation and decreased ossification and affects the formation of cranial base cartilage unions, TMJ condyles, and joint discs. Gli1 is predominantly expressed during early craniofacial development, and Gli1+ cells are identified as the primary mesenchymal stem cells (MSCs) for craniofacial bones, crucial for cell differentiation and morphogenesis. In addition, a complex mutual regulatory mechanism exists between Gli1 and Ihh, ensuring the normal function of the Hh signaling pathway by directly or indirectly regulating each other’s expression levels. And the interaction between Ihh and Gli1 significantly impacts the normal development of craniofacial tissues. This review summarizes the pivotal roles of Gli1 and Ihh in the intricate landscape of mammalian craniofacial development and outlines the molecular regulatory mechanisms and intricate interactions governing the growth of bone and cartilage exhibited by Gli1 and Ihh, which provides new insights into potential therapeutic strategies for related diseases or researches of tissue regeneration.
Introduction
The Hedgehog signaling pathway
The inception of the Hedgehog (Hh) gene traces back to its discovery in the fruit fly, where mutations in the gene induced a transformative shift from hairless to hairy body parts in larvae, hence its apt nomenclature as the “hedgehog” gene. 1 The Hh signaling pathway involves the Hh protein and two cell surface transmembrane receptors: patched1 (Ptch1) and smoothened (Smo), which collectively mediate the signaling cascade. Ptch1 and Smo interact with primary cilia in target cells, facilitating signal transduction mediated by the Gli zinc finger transcription factors (Gli). 2 –5 With the lack of Hh ligands, Ptch1 inhibits Smo, repressing downstream factor expression through the inhibition of Gli zinc finger transcription factors (Gli). 6 Upon Hh expression, the Hh ligand binds to Ptch1, catalyzing the liberation of Smo. Activated Gli transcription factors then ingress into the cell nucleus, thereby initiating the transcription of downstream factors. 7,8 Following their entrance into the nucleus, GLI transcription factors start to express downstream target genes, including transcriptional loop control of Ptch1, Gli1, and Hhip, which are components of the Hh pathway. 9 Members of the Hh family are involved in a variety of cellular functions, such as proliferation, migration, differentiation, and cell fate determination. 10 The Hh signaling pathway significantly regulates mammalian development and homeostasis. The Hh protein comprises three subtypes: Indian Hh (Ihh), Sonic Hh (Shh), and Desert Hh (Dhh), serving as a family of secreted signaling molecules. 11 Various studies have highlighted the essential role of Hh signal transduction in the development and maintenance of skeletal tissue. This signaling pathway exerts significant regulatory effects across various developmental stages, affecting intrinsic processes in both embryonic and postnatal skeletal growth and development. 12
Among the three Hh ligands in mammals, Ihh is pivotal in bone and cartilage development both prenatally and postnatally, and in bone metabolism and cartilage homeostasis maintenance, also participates in intramembranous ossification, chondrocyte proliferation, maturation, and ossification, making it a central regulatory factor in skeletal development and the primary initiator of biological effects in Hh signaling. 13 –19 Meanwhile, osteoblast development is regulated by all Gli factors (Gli1, Gli2, and Gli3) in response to Hh activation, 20 with the expression of Gli1 indicating Hh signal activity 21 and facilitating its activation. 22 In addition, during endochondral ossification, Hh-mediated differentiation of osteoblast lineages depends on the presence of Gli1 protein. 23
Ihh and Gli1 play key roles in developmental regulation
Hh gene subtype Ihh is mostly expressed in hypertrophic chondrocytes and the prehypertrophic layer of the growth plate. 19 Its expression is more pronounced in prehypertrophic chondrocytes near the proliferative zone but weaker in hypertrophic chondrocytes within the growth plate. 20 Ihh binds to Ptch1, prompting Ptch1 to move from the primary cilium on the cell surface, resulting in Smo phosphorylation and activation of Gli proteins, such as Gli1, Gli2, and Gli3. 24 Ihh expression is verified in the ossifying skeleton and at the osteogenic front during ossification. 25 Noteworthy insights from Ihh−/− mice skulls and the misexpression of Ihh in the embryonic chicken head, as Abzhanov et al. 26 propose, are that Ihh, produced by mature osteoblasts in the cranial periosteum, exerts a feedback inhibitory effect on the transformation of preosteoblasts into chondrocyte-like osteoblasts. The phenotype observed in Ihh-null mutant mice unequivocally illustrates Ihh’s role in three Hh signaling-dependent processes during endochondral ossification: proliferation and differentiation of chondrocyte within cartilage and the osteoblasts’ specialization into osteoblasts that form bone. 20
Gli1 was initially isolated from human brain gliomas. 27 As a transcription factor within the Hh signaling pathway, Gli1 plays a pivotal role in the conserved Hh-Gli signaling pathway. It regulates the expression of Hh target genes 8 and is crucial for tissue homeostasis. 9 In bone, Gli1 promotes osteoblast differentiation while concurrently inhibiting their maturation. Upon activation, Gli1 translocates into the cell nucleus, where it induces the expression of various context-specific genes. 28 Genetic labeling studies have confirmed Gli1 as a general marker for mesenchymal stem cells (MSCs) in vivo across various organs. Gli1+ MSCs are primarily found in the cardiovascular system, intestines, liver, kidneys, teeth, and skeleton. Gli1+ MSCs exhibit predominant distribution in the teeth, liver, intestines, kidneys, cardiovascular system, and bones. 29 –34 Studies utilizing genetically modified mouse models by Park et al. revealed that Gli1−/− mice experienced a significantly reduced survival rate within the first 10 days postbirth. In addition, surviving Gli1−/− mice demonstrated lower growth rates and body weights compared to wild-type (WT) and Gli1−/+ mice. 35 Findings from Hojo et al. 13,23 also indicated impaired bone formation in Gli1−/− mice. Within Gli1−/− mitochondria cells, the expression of Col2a1 and Col10a1 was observed, while Runx2 or Sp7 was not expressed. Gli1 activates the transcription of early and mid-stage osteoblast markers by directly binding to the 5ʹ regulatory regions of these genes. It disrupts the activation of chondrocyte marker genes by Sox9 through inhibition of Sox9's DNA binding. Researchers 36,37 have consolidated insights from investigations into the expression and functionality of Gli1+ MSCs, indicating their pivotal involvement in maintaining tissue equilibrium, facilitating mesenchymal rejuvenation, and notably contributing to craniofacial bone osteogenesis. The fate of these cells is set by a signaling-shaped gene regulation network that actively participates in vital biological processes for the formation and regeneration of the skeleton and teeth.
Craniofacial bones, crucial for facial structure and chewing function, originate from neural crest cells and mesenchymal cells originating from the axial mesoderm. Neural crest cells have a major impact on the cranial section of the skull, including the frontal bone, midface bones, and mandible. 38 However, the specific regulatory mechanisms underlying craniofacial bone and cartilage development are complex, and related research findings are scattered. The Hh signaling pathway plays a crucial regulatory role in bone and cartilage development. Based on the pivotal effects of Gli1 and Ihh in the Hh signaling pathway mentioned above, understanding their roles in craniofacial bone and cartilage development is essential for comprehending their functions in tissue injury repair and regeneration. This article aims to review their contributions to craniofacial tissue development. Specifically, it focuses on summarizing and discussing their expression patterns, molecular regulatory mechanisms, and interactions in bone and cartilage formation within the context of existing research.
Expression Patterns of Ihh and Gli1 in Craniofacial Bone and Cartilage Development
Ihh is a member of a highly secreted signaling protein family that orchestrates fundamental processes in embryonic development. 39 It is notably present during the development of cranial osteoblasts and is expressed by these osteoblasts. Interference with Ihh signaling has been shown to impair osteoblast generation. 40 Research findings indicate Ihh expression during the osteoblast proliferation process in murine osteoprogenitor cells. 41 Literature reports consistently highlight the involvement of Ihh in various facets of cranial bone development, tissue structure, cranial base cartilage fusion, and temporomandibular joint (TMJ) growth. 26,38,42
Ihh is expressed in condensed mesenchymal cells during the early stages of cartilage formation. In the initial mesenchymal condensation process preceding the formation of the articular disc at E13.5 in mice, Ihh is present in the condyle of the TMJ. Its expression persists in early chondrocytes at E15.5, and by E16.5, Ihh becomes the sole detectable Hh signal in the TMJ. It is prominently expressed in the early chondrocytes during the condylar enlargement stage and continues in enlarged and hypertrophic chondrocytes at E18.5. 43,44 Ihh expression was identified in the mesenchyme between sagittal sutures of E17.5 mice, albeit at a lower level than in ossified bone and osteogenic fronts. 25 During palatine bone development, the expression of Ihh experienced a notable increase from E13.5 to E17.5, reaching its peak at E15.5. 45
A pivotal developmental process associated with Ihh is endochondral ossification, a fundamental bone development mechanism observed in most axial and appendicular skeletons, along with much of the cranial base. This process primarily unfolds when mesenchymal cells differentiate into chondrocytes. 46 In the structure of TMJ, the mandibular condyle, developed through endochondral ossification, represents a critical region for mandibular growth with significant implications for chewing and speech. 47 Ihh expression has been verified in the condylar cartilage of mice at E15.5. 39 Another critical structure related to TMJ development is the TMJ disc. Liang et al.’s histological study 48 suggests that cells may separate from the condyle’s fibrous surface, forming the disc. The top compartment of the TMJ is produced by cellular death of loose mesenchymal cells that exist between the disc and the fossa at E15.5, while the lower compartment is formed by the complete separation of the disc cell layer from the condyle at E17.5. By E18.5, the TMJ exhibits a well-defined upper joint cavity, a distinct boundary of the articular disc, and the early formation of the lower joint cavity. Ihh is expressed in the mesenchymal cells between the condylar fibers before the formation of the TMJ disc. 49,50 Table 1 summarizes the earlier discoveries regarding the involvement of Ihh in the formation of craniofacial bones and cartilage.
Expression Patterns of Ihh in Craniofacial Bone and Cartilage Development
E, embryo; Ihh, Indian Hedgehog.
Research on GLI signaling in craniofacial development remains relatively limited, with Du et al. 51 among the pioneering contributors in studying the expression patterns of Gli1-3 in the embryonic mouse face. Their findings reveal that Gli1 is highly expressed in the maxillofacial area and mandibular prominences between E11 and E14.5, alongside other early phases of craniofacial development. Notably, at E11-12.5, Gli1 expression is more robust in Meckel’s cartilage compared to surrounding tissues. Post the ossification of Meckel’s cartilage, Gli1 becomes predominantly expressed in the mandible. Li J et al. 52 found that during the development of the cranial vault in WT mice, Gli1 is widely expressed in the cranial neural crest (CNC) derived at E11.5. The expression of Gli1 is significantly downregulated in the dense mesenchyme of the cranial rudiment at E12.5. The transcripts of Gli1 are distinctly detected in preosteoblasts at E13.5, suggesting that the regulatory levels of Hh activity may be crucial for the fate determination of condensed mesenchyme toward preosteoblasts. The relevant expression of Gli1 in palatine bone development is significantly upregulated between E11.5 and 17.5 days, reaching its peak at E15.5. Levi et al. 45 detected a significant upregulation of Gli1 expression related to palatine bone development between E11.5 and 17.5 days, reaching its peak at E15.5. Atsushi et al. 53 identified widespread expression of Gli1 in the mandibular mesenchyme of wild-type mice at E11.5. By E12.5, Gli1 expression spans throughout the mesenchyme in the anterior part of the mandible but becomes restricted in the posterior part, particularly in the endogenous bone region. Importantly, Gli1 expression is not detected in the mesenchyme on the tongue side of Meckel’s cartilage. Furthermore, Gli1 expression undergoes significant downregulation in most of the mesenchyme from the anterior to the posterior part of the mandible. The findings regarding the expression pattern of Gli1 in craniofacial development are consolidated in Table 2.
Expression Patterns of Gli1 in Craniofacial Bone and Cartilage Development
Gli1, gli zinc finger transcription factor 1; E, embryo; CNC, cranial neural crest.
Key Roles and Functional Effects of Ihh and Gli1 in Craniofacial Bone and Cartilage Development
MSCs, known for their ability to self-renew and differentiate, play an important role in tissue homeostasis. 54,55 The Hh signaling pathway emerges as a crucial regulator in embryonic bone development and postnatal bone reconstruction by orchestrating MSC-mediated osteogenesis. 36 The Gli1+ cells have been demonstrated by multiple studies to be the main MSCs of craniofacial bones. 32,36,56 –58 In the following, we will review the significant functional roles of Ihh and Gli1+ MSCs in the two main ossification processes of the craniofacial bones.
Ihh and Gli1+ MSCs are two key influencing factors in intramembranous ossification
Craniofacial bones primarily undergo intramembranous ossification, in which Ihh serves as a key upstream factor in the Hh signaling pathway critical for craniofacial development. As indicated by previous research, the absence of Ihh leads to a distinct phenotype in mice, characterized by impaired bone development, reduced cranial bone size, diminished ossification, and notably widened cranial sutures. 59 –61 The diminished ossification in Ihh−/− mice correlated with an overall reduction in osteogenic markers and decreased expression of BMP2/4, affirming Ihh’s pivotal role in cranial bone ossification. 25 At E16.5 and E17, mice with an Ihh gene defect (Ihh−/− ) displayed diminished ossification in the palatine bones and a notable increase in clefts at the midline of the palate. This observation shows that Ihh may help with the osteogenic differentiation of the secondary palatal shelves. 45
Sutures, crucial structures influencing development within craniofacial bones, fibrous connections that link the bones of the skull, act as dynamic habitats in which mesenchymal stem cells manage skull developmental and growth processes. 62 Zhao et al. 32 using lineage tracing identified Gli1+ cells in cranial sutures as the principal MSCs for craniofacial bones, with Gli1+ MSCs in the cranial bone marrow exhibiting notably inferior functionality. Ablating Gli1+ MSCs resulted in impaired cranial bone growth, osteoporosis, and compromised damage repair. Pietro et al. 56 observed Gli1+ MSCs in bone trabeculae of human cranial tissue samples, further supporting their role in craniofacial bone development. Xu et al. 57 analyzed the single-cell RNA sequencing (scRNA-seq) dataset of E18.5 cranial sutures and found that osteoblasts that were Gli1+ and Ptch1+ were more numerous than chondrocytes. This suggests that during the development of cranial bones, intramembranous ossification osteoblasts exhibit a substantial activation of the Hh signaling pathway. In addition, Gli1 also plays an indispensable and crucial role in this process.
As summarized in Table 3, it can be inferred that Gli1+ MSCs and Ihh are two key influencing factors in intramembranous ossification during craniofacial bone and cartilage development, playing important roles in promoting osteogenic differentiation of craniofacial bones.
The Vital Roles of Ihh and Gli1+ MSC in Intramembranous Ossification during Craniofacial Development
E, embryo; Ihh, Indian Hedgehog; BMP, bone morphogenetic protein; Gli1, Gli zinc finger transcription factor 1; MSCs, mesenchymal stem cells.
Ihh and Gli1+ MSCs are essential prerequisites for endochondral ossification
For structures like TMJ and cartilage junction, ossification occurs through endochondral ossification. Regarding the development of the TMJ, generalized Ihh knockout or conditional knockout of Hh signaling during embryogenesis causes aberrant disc and synovial cavity formation, as well as underdeveloped condylar cartilage. 39 Inactivating or modulating Ihh signaling in the cartilage during postnatal life leads to delayed condylar cartilage development and disc fusion. 5,12,43,49 Kurio et al. 7 discovered that conditional Ihh loss in Ihhf/f ;Aggrecantm(IRES-creERT2) knockout (KO) mice reduced the proliferation and chondrogenesis of progenitor cells. Conditional Ihh deletion in the condylar cartilage is associated with severe disorganization of the multiplicity/progenitor cell layer tissue, accompanied by significantly reduced proliferation of cartilage progenitor cells and decreased expression of Tn-C/Sox9 in this layer. Chondrocyte progenitors in the condylar cartilage are highly sensitive to Hh signaling for normal postnatal function, and Ihh signaling plays a crucial role in maintaining the characteristics and functions of chondrocyte progenitors in the condylar cartilage during both juvenile and adult stages.
Phenotypically, Ihh−/− mice exhibit the presence of chondrocytes in the condyle at E15.5, but the primordium of the articular disc is either absent or indistinguishable. At both E17.5 and E18.5, there is no articular disc or joint cavity; therefore, the condyle is immediately facing the temporal bone. Gene expression analysis reveals the absence of Ihh transcripts at E15.5, accompanied by the lack of detectable RNA encoding patched, Gli1, and Gli2. In addition, the expression of Sox9 decreases significantly at E15.5 and E18.5, and PTHrP expression is undetectable. 39 In the study by Ochiai et al., 49 partial adhesion between the articular disc and the condyle surface was observed in Ihh(−/−) mice at both P4 and 8 weeks of age, indicating that the observed articular disc defects during embryonic development may persist into postnatal stages. Therefore, Ihh may play an upstream and crucial role in the mechanism of articular disc formation. If Ihh expression is lacking in the condyle, 63 the TMJ disc cannot form, indicating that Ihh signaling in the condyle directly induces disc formation early on, and the disc formation is independent of Ihh expression in the condyle.
When Ihh is absent, the tissue in the condylar growth plate–like region of the TMJ condyle is irritated. Mice missing Ihh lack the articular disc and have a smaller joint cavity, resulting in severe joint abnormalities at birth. And Ihh-mediated signaling is considered crucial for the creation and maintenance of the TMJ disc. 39,43,49,64
As a crucial downstream activator of the Hh signaling pathway, Gli1 has been found to undergo significant expansion in a lineage tracing study spanning 12 months. This expansion occurred in the population of Gli1+ MSCs located at the junction of condylar cartilage and mandibular bone tissue, and they were found to be distributed across almost all bone trabecular surfaces. 36 These Gli1+ MSCs, serving as progenitor cells of osteoblasts, contribute to the formation and homeostasis of the mandibular condyle. 58 Researchers detected local activation of Gli1+ MSCs at the fracture site in a model imitating condylar bone fractures to evaluate the response of Gli1+ MSCs to external triggers during the healing process. These cells migrated to the callus and differentiated into osteoblasts, actively participating in the regeneration of damaged bone. Zhang et al. 65 also noted that Gli1+ cells are selectively distributed in specific areas of the periosteum and condyle, with their differentiation ability significantly constrained to distinct regions. Notably, periosteal cells exclusively showed osteogenic potential, but Gli1+ cells in the condyle showed both osteogenic and chondrogenic potential.
Considering the limited survival rate of Ihh-deficient mice postnatally, studies on Ihh’s impact on postnatal development are constrained. Amano et al. 66 established a Wnt1-Cre;Ihhfl/fl ;Tomatofl/+ mouse model, allowing mice to survive up to two weeks postnatally. These mice displayed skeletal deformities characterized by simultaneous incomplete or poorly developed nasal and maxillary complexes and mandibles. According to these findings, endochondral ossification is initially delayed by cartilage mutations, and the preservation of the intersphenoid synchondrosis (ISS) is progressively impacted by deficiencies in chondrocyte differentiation and proliferation caused by conditional Ihh loss.
Ihh−/− mice, characterized by ectopic calcification in the cartilage junction between the cranial base synchondrosis, exhibit a delayed differentiation of hypertrophic chondrocytes. 59 This delay further results in disrupted synchronized growth plates in Ihh−/− mice, leading to defects between prehypertrophic chondrocytes expressing Ihh and proliferating chondrocytes expressing PTHrP. 42 These findings underscore Ihh’s indispensable role in cranial base cartilage fusion and overall cartilage formation. Koyama et al. 4 observed that the conditional deletion of Ihh in chondrocytes, utilizing Col2aCreER in neonatal mice, resulted in a shorter cranial base, disorganized cartilage fusion, and early fusion of cartilage junctions by day 15 postnatally. Ihh collaborates with other proteins, such as Ellis-van Creveld protein (EVC) and Kif3a, to coregulate endochondral ossification, stimulating chondrocyte proliferation and regulating maturation, thus promoting cranial base elongation.
Therefore, Ihh affects chondrocyte differentiation and proliferation during endochondral ossification in craniofacial cartilage, whereas Gli1+ cells support both osteogenic and chondrogenic potential. These processes are crucial for the growth of bones and cartilage in the craniofacial region. Research findings related to Ihh and Gli1+ MSC in endochondral ossification during craniofacial development are outlined in Table 4.
The Vital Roles of Ihh and Gli1+ MSC in Endochondral Ossification during Craniofacial Development
E, embryo; P, postnatal; Ihh, Indian Hedgehog; TMJ, temporomandibular joint; ISS, intersphenoid synchondrosis; Gli1, Gli zinc finger transcription factor 1; MSCs, mesenchymal stem cells.
Gli1 and Ihh Interaction
Molecular mechanisms of mutual regulation
The interaction between Gli1 and Ihh encompasses intricate molecular mechanisms. Under normal conditions, the active Ihh signal intricately regulates the expression level of Gli1. Ptch1 and Gli1, as direct transcriptional target genes of the Hh signal, are both upregulated by Ihh. 15 This regulation may manifest through direct action on the Gli1 gene’s promoter region or by controlling the stability of Gli1. Concurrently, Gli1 also participates in regulating Ihh expression, forming a mutual regulatory feedback loop. This precise regulatory mechanism ensures the normal functioning of the Hh signaling pathway during development.
Ihh must bind to Ptch1 in articular cartilage for the Hh pathway to be activated. Ptch1 then reduces Smo’s inhibition, which causes Smo to be activated and enriched in the primary cilium, an organelle made of microtubules that is found in a variety of cells, including articular chondrocytes. 2,67,68 Hh target genes like Ptch1 and Gli1 are expressed when Smo subtly triggers downstream Gli transcription factors, leading them to translocate to the nucleus. 2,67,69,70 Zhao et al.’s research 32 demonstrated the impact of the Hh signaling pathway on Gli1-CreERT2; Smofl/fl mouse cranial suture MSCs. Notably, after 8 months of tamoxifen inference, they saw a significant loss in bone mass and serious osteoporosis in cranial bones. This condition was successfully corrected by Ihh agonist treatment, leading to the upregulation of Gli1 activity.
Role in cellular differentiation
The interplay between Gli1 and Ihh assumes a pivotal role in cellular differentiation and tissue morphogenesis in craniofacial cells. Various signaling pathways may orchestrate this interaction, particularly in skeletal and dental tissues, where normal cell differentiation is indispensable for the proper formation of these tissue structures. Among these pathways, bone morphogenetic proteins (BMPs), integral members of the transforming growth factor-β (TGF-β) superfamily, emerge as key regulators, exerting control over the size, shape, and positioning of teeth during craniofacial development. 71 Insights from studies propose that BMP signaling not only profoundly influences the differentiation of odontoblasts but also holds significance in providing feedback to sustain the Gli1+ MSC population. 72,73
Moreover, findings from Guo et al. indicate that Gli1+ MSCs possess the capability to generate osteoprogenitors, exhibiting active BMP signaling in cranial sutures. Conditional inactivation of Bmpr1a in Gli1+ MSCs, resulting in an imbalance between osteogenic and osteoclastic activities, leads to weakened Hh signaling and a narrowing of sutural lines. In an injury model, the loss of Bmpr1a in Gli1+MSCs;Bmpr1afl/fl mice diminishes cranial bone injury healing. Conversely, the upregulation of Hh signaling in Gli1-CreERT2 promotes cranial bone healing. This underscores that cranial suture Gli1+ MSCs depend on BMP-mediated Ihh signaling to balance osteogenic and osteoclastic activities in cranial bone defect repair. Such intricate interactions contribute significantly to the maintenance of cranial tissue homeostasis and repair. 74
Interaction in the functions of bone and cartilage cells
The interplay between Gli1 and Ihh exerts a direct impact on the functions of bone and cartilage cells. The reciprocal regulation of Gli1 and Ihh signaling significantly influences the proliferation, differentiation, and matrix synthesis of both bone cells and chondrocytes. Investigations on senescence-accelerated (SAMP) mice, serving as a model for rapid aging, alterations in Ihh signaling activity, are linked to the thickening and thinning of the epiphyseal cartilage. This reduction is manifested by reduced expression of Gli1, Gli2, and/or Ptch1 in Ihh target cells, indicating that disrupted Ihh signaling indirectly contributes to the epiphyseal cartilage thickening through abnormal matrix transformation and accumulation. 12 In vitro cell experiments indicate that Ihh can directly promote chondrocyte hypertrophy and matrix mineralization, inducing the expression of Runx2 in reactive chondrocytes. 75 Notably, the Gli1 transcription factor in the Hh pathway facilitates the direct activation of Runx2 and Runx3 transcription, with Runx2 further regulating the maturation and proliferation of chondrocytes through inducing Ihh expression. 76 –78
However, specific studies focusing on the craniofacial region are limited in literature. Ihh signaling, crucial for normal trabecular bone growth in long bones, faces disruption upon genetic intervention, impairing the proliferation and osteogenic differentiation of Gli1+ mesenchymal progenitor cells (MMPs) in the metaphysis. 33 In the context of a bone injury study, 79 it was observed that SAG, a specific Hh signal activator, may heighten Ihh expression in chondrocytes through the GLI1-RUNX2-IHH signaling axis. This activation has been reported to promote bone healing in a bone defect model. Conversely, dysregulation of the Gli1 and Ihh interaction may lead to abnormal development of bone and cartilage tissues, potentially triggering certain diseases.
This parallels findings from cell culture experiments, where the interaction between Gli1 and Ihh directly or indirectly influences the expression of Runx2, or one of them affects the expression of the other through Runx2, thereby regulating cellular functions. Therefore, despite limited research on the craniofacial region, based on existing findings, we might speculate that Gli1 and Ihh have similar mechanisms of interaction in regulating cellular functions during the development of craniofacial bones and cartilage.
Summary
In conclusion, it is evident that Ihh and Gli1 contribute pivotally to the regulation of craniofacial development (as shown in Fig. 1). Ihh engages in multiple processes, including chondrocyte differentiation, proliferation, and bone formation, exerting significant influence on the development of cranial bones, cartilage, mandibular condyle, and the formation of TMJ. Loss of Ihh results in cranial bone malformation, reduced cranial ossification, and impacts the growth of the cranial base cartilage union, protrusion of the TMJ condyle, and the formation of the joint disc. Gli1, predominantly expressed in early craniofacial development, has been identified as the main MSCs in the cranial sutures, and its potential for osteogenesis and chondrogenesis in different locations plays a crucial role in cranial growth, development, and repair. Gli1 and Ihh play important regulatory roles in different modes of ossification (intramembranous ossification and endochondral ossification) during craniofacial development. There exist complex regulatory mechanisms between the two, where they maintain the normal functioning of the Hh signaling pathway by directly or indirectly regulating each other’s expression levels. The interaction between Gli1 and Ihh holds paramount importance for the normal development of craniofacial tissues, influencing cell differentiation and cellular functional transitions. Dysregulation of the regulatory pathways of Gli1 and Ihh may lead to abnormal development of bone and cartilage tissues.
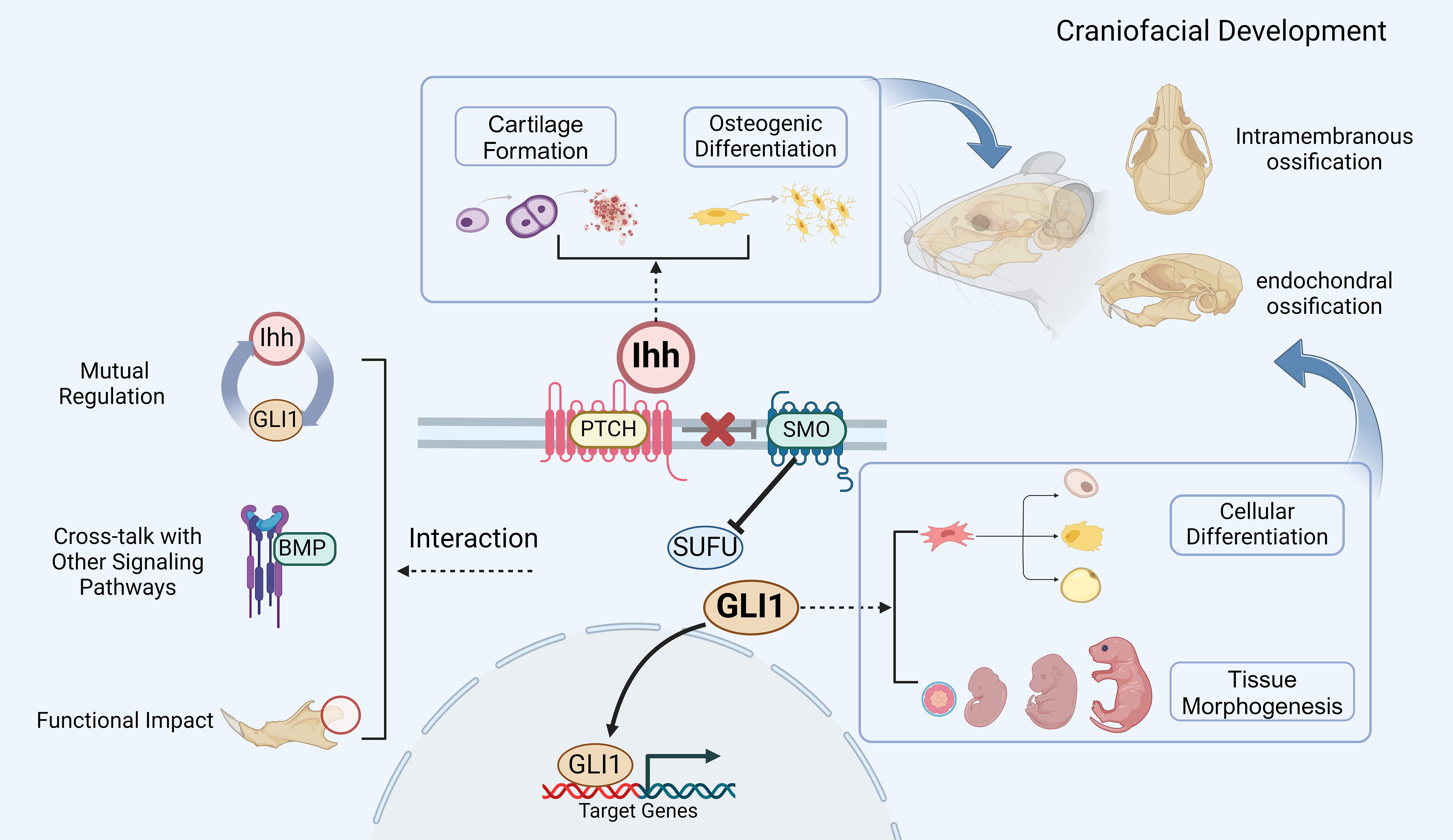
This diagram provides a brief overview of the roles of Ihh and Gli1 in craniofacial development. The diagram illustrates the main regulatory processes of Gli1 in the Ihh signaling pathway. In this context, Ihh plays a crucial role in craniofacial ossification and cartilage formation, while Gli1 has significant effects on the differentiation of MSCs and tissue morphology. The interaction between Ihh and Gli1 is mainly manifested through molecular mechanisms of feedback loops, crosstalk with other signaling pathways (e.g., BMP), and their effects on the functions of bone and cartilage cells. The image was created using BioRender.com (accessed on February 14, 2024). Ihh, Indian Hedgehog; PTCH, patched transmembrane receptors; SMO, smoothened transmembrane receptors; SUFU, suppressor of fused; Gli1, Gli zinc finger transcription factor 1; BMP, bone morphogenetic protein.
As previously mentioned, Gli1 is primarily expressed in the mesenchyme, and the Hh signaling pathway is vital for MSC-mediated osteogenic activity during embryonic and postnatal bone development and reconstruction. Nevertheless, several aspects still require further investigation. First, the specific expression patterns of Gli1 in various craniofacial regions at different developmental stages, and its relevance to cell differentiation and morphogenesis fate determination, remain inadequately explored. Second, the specific signaling transduction modes, molecular regulatory mechanisms, and the underlying gene regulatory networks involving Gli1 and Ihh in craniofacial MSCs remain unclear. Further in-depth analysis is needed to understand the expression and transformation patterns during development, necessitating additional basic and translational research. However, deeper investigation into the developmental mechanisms of Gli1 and Ihh will likely enhance our understanding of craniofacial skeletal development regulation, which will significantly contribute to addressing developmental defects and injury repair, offering new insights for tissue regeneration-related engineering research and providing novel therapeutic strategies for related disease.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key Clinical Specialty Development Project of Pediatric Dentistry Division of China and Kunming Medical University Innovation Team Project (CXTD202213).