
Abstract
Adipose stem cells are considered one of the primary drivers of autologous fat graft biological activity and survival. We have previously demonstrated that hormonally active VD3 improved adipose stem cell viability in ex vivo and in vivo fat grafting models. In this study, we evaluated the inactive form of VD3 (cholecalciferol) on adipose stromal cell (ASC) phenotype during hypoxia and the subsequent effect on human fat graft retention in the xenograft model. Lipoaspirate collected from six human donors was used for ex vivo particle culture studies and isolated ASC studies. Adipose particles were treated with increasing doses of VD3 to determine impact on ASC survival. Expanded stromal cells were treated with VD3 during hypoxic culture and assessed for viability, apoptosis, mitochondrial activity, and nitric oxide (NO) release via caspase, DAF-FM, or TMRM. Finally, 40 Nu/J mice receiving bilateral dorsal human lipoaspirate were treated thrice weekly with (1) vehicle control, (2) 50 ng calcitriol, (3) 50 ng VD3, (4) 500 ng VD3, and (5) 5,000 ng VD3 for 12 weeks, n = 8 per group. Graft weight, volume, and architecture were analyzed. Adipose particles treated with dose-escalating VD3 had significantly increased ASC viability compared with control (P < 0.01). Under hypoxia, ASCs treated with 1 nM VD3 had significantly greater viability than untreated and pretreated cells (P < 0.01, P < 0.01) and significantly lower apoptosis-to-viability ratio (P < 0.01). ASCs pretreated with 1 nM VD3 had significantly lower NO release (P < 0.05) and lower mitochondrial polarization (P < 0.05) compared with controls. In vivo results showed mice receiving 5,000 ng VD3 had significantly greater graft weight (P < 0.05) and volume (P < 0.05) after 12 weeks of treatment compared with controls. Grafts had enhanced neovascularization, intact adipocyte architecture, and absence of oil cysts. VD3 is an over-the-counter nutritional supplement with a known safety profile in humans. Our xenograft model suggests administering VD3 at the time of surgery may significantly improve fat graft retention.
Introduction
Autologous fat grafting is a surgical technique for replacing soft tissue volume in reconstructive and cosmetic surgery and restoring tissue barrier function in otolaryngology 1,2 and colorectal 3,4 indications. Because adipose tissue also serves a critical role in protecting the body from mechanical forces, fat grafting has been successful in cushioning prosthetic attachment sites 5 and replacing pedal fat pads. 6 Although fat grafting has been used to improve the quality of life for tens of thousands of patients a year, hundreds of thousands more potential patients currently elect not to receive fat grafts owing to known complications such as unpredictable graft failure, tissue necrosis, and calcification. 7
In fat grafting procedures, excess autologous adipose tissue is harvested and grafted to a new location with the goal of permanent soft tissue replacement. Following injection, recipient sites exhibit a brief period of edema followed by three months of volume loss, after which graft volume stabilizes, typically retaining between 40% and 60% of injected volume. 8 Clinicians compensate for expected volume loss by overfilling defect sites, often by 50% or more. 9 As graft take can vary widely, with outcomes ranging from 10% to 90%, 10 physicians must carefully manage patient expectations by forewarning that multiple procedures may be necessary to achieve desired results. Although limitations in outcome predictability have constrained technique adoption, fat grafting has enormous potential to transform patient care, and continued efforts to improve surgical outcomes are warranted.
Previously, we showed that calcitriol, an Food and Drug Administration (FDA)-approved analog of hormonally active VD3, was a potent lipoprotectant from ischemic stressors and significantly improved human fat graft survival in an established xenograft mouse model. 11 Mechanistically, calcitriol increased adipose stromal and stem cell survival, decreased postgrafting tissue hypoxia, reduced graft fibrosis, increased graft revascularization and adipocyte content, and improved volume retention overall. However, its continuous high-dose administration may induce hypercalcemia or vascular calcifications. 12 –14 In contrast, nutritional VD3 (cholecalciferol, hereafter referred to as VD3) is a widely used over-the-counter oral supplement with an established safety profile. The overall goal of this study was to determine if inactive VD3 is a feasible alternative to calcitriol for protecting adipose stromal cells (ASCs) during nutrient and oxygen deficiency and improving long-term fat grafting retention outcomes. To test this, a series of in vitro and in vivo survival studies were conducted using human primary adipose cells, tissue culture approaches, and the established immunocompromised mouse xenograft model.
Materials and Methods
Human adipose tissue collection
Lipoaspirate was collected from discarded abdominal resections from 6 unique human donors undergoing elective panniculectomy under University of Pittsburgh Institutional Review Board (IRB) exemption (STUDY23100134). Liposuction was performed on full-thickness human skin samples using a 60-mL syringe (Medline SYR110010) attached via luer-lock to a 1 mm blunt tip cannula (Mentor Worldwide LLC MER4155) in a biological safety cabinet according to our previous publication. 11 Lipoaspirate was collected, divided for use in particle or cell studies, and processed as described below. Tissue from individual donors was never mixed, but used independently to reproduce experimental results, after which data were aggregated for analysis.
Measurement of VD3 effect on whole adipose survival with ex vivo tissue culture
Ex vivo particulate culture was set up as previously described. 11 Briefly, for each adipose donor, 1 mL of lipoaspirate was added to 21 wells of multiple 6-well plates, fully submerged with a mesh filter, wholly covered with basal media, phenol-free McCoys 5A media (Gibco; Cat: 16600082) with 10% Fetal Bovine Serum (FBS) and 1% penicillin-streptomycin (Thermo Fisher) and cultured in a 5% CO2 humidified incubator. Experimental test groups included a vehicle control (0.3 mL Phosphate Buffered Saline (PBS) with 0.1% EtOH), calcitriol (15.6 nM, 62.5 nM, or 250 nM), or cholecalciferol (15.6 nM, 62.5 nM, or 250 nM), n = 3 replicates per group. Particles were cultured for 7 days with one intermittent media change. At culture completion, samples were weighed and digested with collagenase type II, as previously described. 11 Initial stromal cell viability was determined via trypan blue exclusion with confirmation of cell survival and size via calcein AM and propidium iodide (Cellometer, Nexelom) as previously described. 11
Measurement of VD3 metabolic conversion during ex vivo tissue culture
To determine if cultured adipose tissue metabolically converted VD3 to the active metabolite, 1α,25-dihydroxyVitaminD3 (DH-VD3), and if tissue particles were saturated with VD3 at tested concentrations, cultured adipose particles were flash frozen after 7 days of culture and the active metabolite, DH-VD3, was quantified by ELISA according to the manufacturer’s instructions (Cloud-Clone Corp, Katy, TX #CEA467Ge). DH-VD3 was extracted from tissue samples according to a modified protocol by Farhangi et al. 15
In vitro analysis of VD3 effects on ASC survival and activation
ASC isolation and culture
Lipoaspirate samples were mixed 1:2 with collagenase type II solution (1 mg/mL, Worthington Biochemical Corp, Lakewood, NJ, in Hanks Balanced Salt Solution with 3.5% bovine serum albumin) and digested as previously described. 11 Nucleated cells were collected with centrifugation (300g for 5 min), after which the aqueous layer was decanted, resuspended in an erythrocyte lysis buffer, and collected again with centrifugation. The remaining stromal vascular cells were resuspended in EGM-2MV™ media (Lonza, Walkersville MD, CC-3156 & CC-4147) and culture expanded until passage 2–3 in tissue culture flasks.
ASC survival in hypoxia
Culture-expanded ASCs were plated in triplicate in 96-well plates at confluence in growth media and allowed to adhere overnight. VD3 (1 nM) was added to cells 0 or 12 h before hypoxia induction with 100 µM cobalt chloride (CoCl2), mimicking cellular hypoxic exposure. 16 Briefly, CoCl2 was dissolved directly into EBM media at 1 mg/mL (4.203 mM), sterile filtered, and diluted as needed. VD3 or vehicle controls were added to growth media, ensuring less than 0.1% ethanol concentration in the final volume. Viability was measured at 6- and 12-hour posthypoxia by propidium iodide exclusion and fluorescent quantification via ApoLive-Glo multiplex Assay (Promega #G6410) with a Tecan M200 Infinite plate reader, normalizing each well internally with 4′,6-diamidino-2-phenylindole (DAPI). Apoptosis was similarly measured at 6 and 12 h with the ApoLive-Glo multiplex Assay (Promega #G6410) following the manufacturer’s instructions and read by the Tecan plate reader with fluorescence (400Ex/505Em) and luminescence specifications.
ASC mitochondrial polarization and nitric oxide synthesis
We additionally assessed (1) mitochondrial polarization [tetramethylrhodamine, methyl ester (TMRM)]; (2) nitric oxide (NO) production [4-amino-5-methylamino-2′, 7′-difluorofluorescein (DAF-FM)]; and (3) Adenosine Triphosphate (ATP) Determination Kit (Thermo Fisher; Cat: A22066). In each assay, culture-expanded ASCs were plated in triplicate in 96-well plates as described above, with VD3 added 0, 6, and 24 h before hypoxia induction with CoCl2.
TMRM was added to each well after culture at a final concentration of 10 nM in media by diluting a 10 mM stock in Dimethyl sulfoxide (DMSO). Cells were incubated in TMRM for 1 h, washed thoroughly with PBS, and then measured for fluorescence with the Tecan plate reader, with at least four measurements performed per well (548Ex/573Em). Data were normalized to DAPI (360Ex/465Em).
DAF-FM diacetate was used to measure cell production of NO, as modified from D’Amico Oblak et al. 17 Briefly, after the described culture treatment, cells were washed once, incubated in a solution of 10 µM DAF-FM diacetate in Hank's balanced salt solution (HBSS) for 1 h at 37°C, and washed three times in HBSS. At 37°C after a 3-sec medium shake, fluorescence (485Ex/516Em) was measured in intervals of 15 min up to 2 h via TECAN. A background control with no DAF-FM and activation controls whereby cells were treated with 100 µM diethylamine (DEA) and 100 µM diethylamine NONOate (DEA/NO) were used.
Extracellular ATP was assessed via quantitative luciferin–luciferase bioluminescence assay as per the manufacturer’s specification. An ATP standard curve was generated using the included standards, and aliquots of media from incubated cells described above were assessed for ATP content by luciferase activity. Luminescence was assessed via TECAN.
In vivo analysis of systemic VD3 effect on human adipose graft survival
Experimental design
All procedures were performed under the University of Pittsburgh IACUC approval #17121915, as previously described. 11 Mice were housed in pathogen-free cubicles and cared for under the supervision of the Division of Laboratory Animal Resources staff. Forty athymic/Foxn1nu female mice (Charles River Laboratories, Inc. 002019) were randomly assigned to five experimental groups: (1) vehicle control, (2) 50 ng calcitriol (historically effective dose), (3) 50 ng VD3, (4) 500 ng VD3, and (5) 5,000 ng VD3, n = 8 animals per group. After isoflurane anesthetization, a volume of 300 µL of processed lipoaspirate, with an estimated weight of 0.270–0.276 grams based on adipose density of 0.90–0.92, was bilaterally injected using a 16G blunt-tip cannula (Beacon Medical Ltd. BE-STD215) to murine dorsal flanks, totaling 16 experimental replicates per group and two unique human adipose donors. Mice received 300 µL intraperitoneal injections of VD3 or calcitriol diluted in 0.1% ethanol on Monday, Wednesday, Friday (MWF) for 12 weeks, whereas control animals received 0.1% ethanol in saline. Animals were sacrificed at 12 weeks by CO2 asphyxiation and cervical dislocation, after which grafts were excised, weighed, and measured for volume using an AccuPyc II 1340 (Micromeritics Instrument Corp., Norcross, GA). Grafts were then either snap-frozen or fixed in 10% neutral buffered saline for further analysis.
Histology
After fixation, samples were embedded in paraffin, cut into two equal halves at the midline, and embedded with the graft center on the block cutting surface. Representative slides from each mouse were obtained with 6 µm tissue sections by Pitt Biospecimen Core. General tissue architecture was obtained by Hemotoxylin and Eosin (H&E). Adipocytes were visualized with immunohistochemistry using antibodies for perilipin (Novus Biologicals NB110-40760, Vector Laboratories BA-1000) and semiquantified as previously described with ImageJ (National Institutes of Health, Bethesda, MD). 11 Graft vascularity was semiquantified by blinded counting of CD31+ (Abcam #ab28364, Cambridge, MA) blood vessel structures from a minimum of 10 images per slide, taken at 10× magnification. Viable adipocytes were similarly semiquantified as perilipin1+ cells (Invitrogen #PA5-72921).
Statistical analysis
Statistical analysis was conducted in GraphPad Prism 10. Tests of normality and heteroscedasticity were conducted, then ordinary one-way analysis of variance (ANOVA) or one-way ANOVA with Brown–Forsythe and Welch tests were performed when appropriate.
Results
VD3 increased ASC survival in whole tissue culture
Adipose particles cultured for one week had significantly increased stromal cell viability with dose-escalating calcitriol and VD3 (Fig. 1A). Calcitriol media supplementation with 15.6, 62.5, and 250 nM significantly increased in vitro stromal cell viability by 4.5%, 6.8%, and 8.1%, respectively (P < 0.01 for all doses). Addition of VD3 to the culture media significantly increased ASC viability with dose escalation for in vitro cultures compared with control (P < 0.01 for all doses). To determine the potential benefit of increased calcitriol or VD3 addition, tissue concentration of the active metabolite was measured to determine saturation levels, which occurred at 62.5 nM for both calcitriol and VD3 (Fig. 1B). Furthermore, adipose particles metabolized inactive VD3 to the active metabolite as indicated by no significant measurable difference in DH-VD3 concentration for calcitriol and VD3-treated particles (P = 0.92).
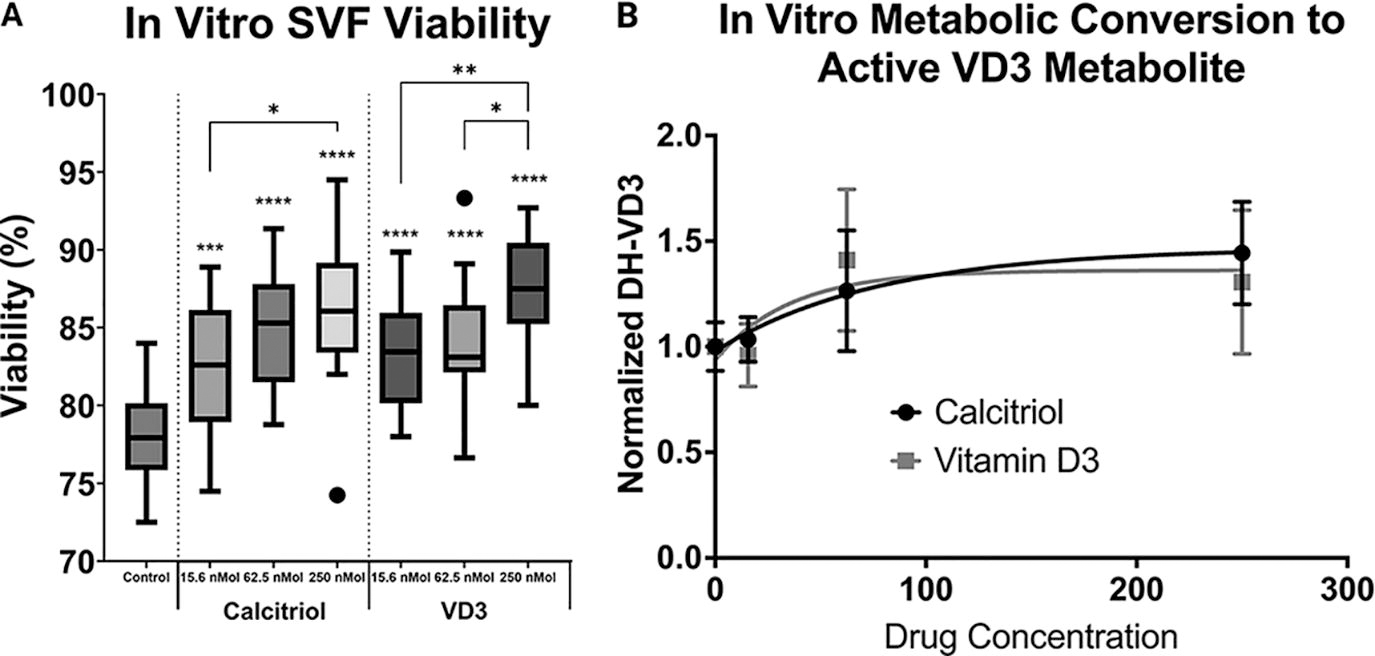
VD3 increased ASC viability under hypoxia
Cell cultures from six unique donors were either pretreated with VD3, treated with VD3, or untreated, with assays performed after 6 and 12 h of hypoxia. Results indicated that 1 nM VD3 treatment significantly augmented ASC viability compared with untreated and pretreated cell cultures after 6 h of hypoxia (Fig. 2A; P < 0.01 for both). However, VD3 had no significant impact on ASC viability after 12 h of hypoxia. Interestingly, cells had no significant difference in apoptotic events at both time points (Fig. 2B), but when the apoptosis to viability ratio (AVR) was calculated, treated cells had a significantly lower AVR than pretreated cells after 6 h (P < 0.01) and both untreated and pretreated cells after 12 h (Fig. 2C; P < 0.01 for both). ATP determination assays were also performed on cell culture media to quantify extracellular ATP after six and 12 h of hypoxia among treatment groups, then normalized to DAPI. Although there were no significant differences among treatment groups at each time point, cell cultures pretreated with VD3 had a 23.6% greater proportion of relative luminescence after 6 h of hypoxia (Fig. 2D).
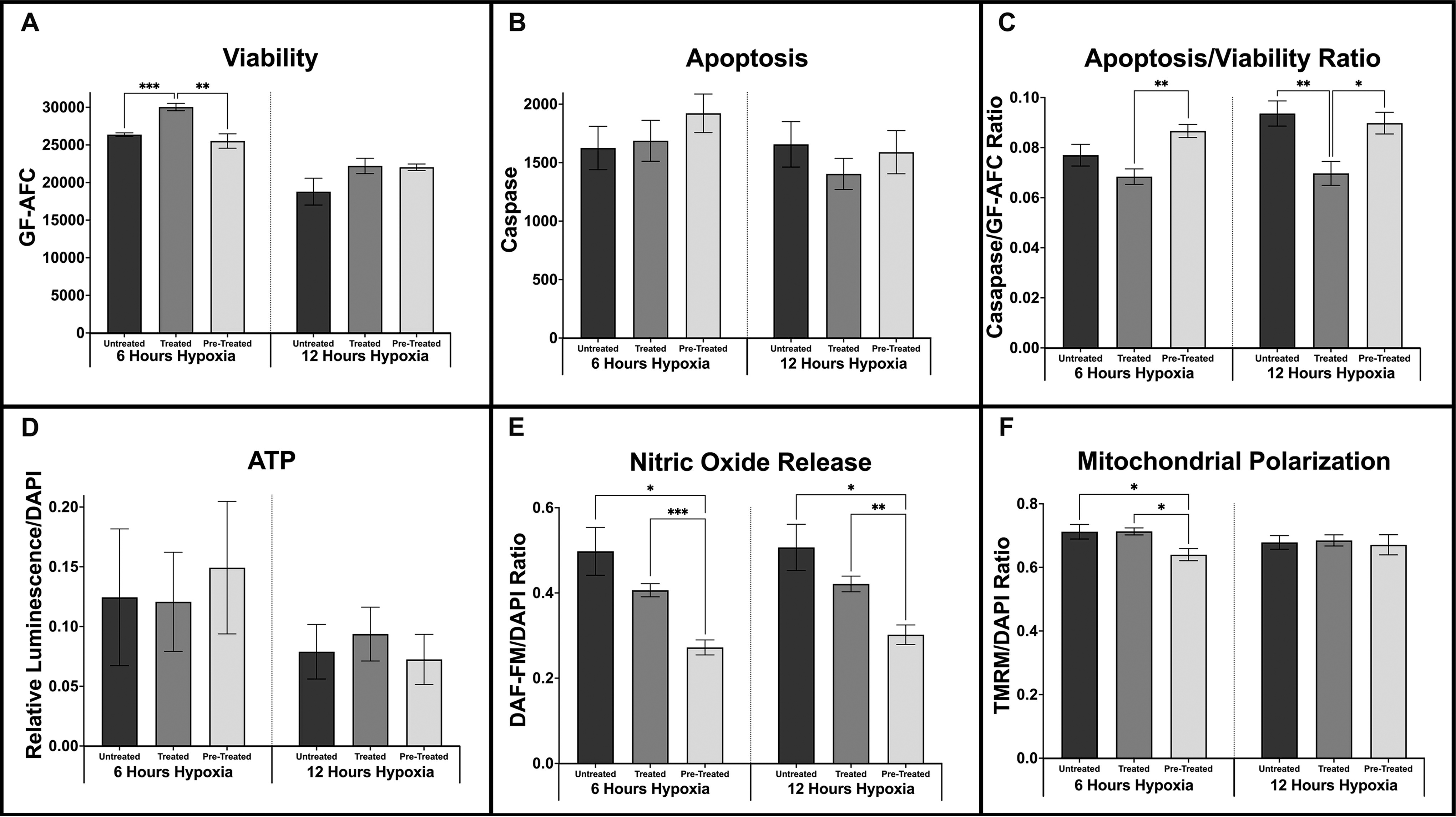
Hypoxia values (mean ± SEM) for untreated, treated, and pretreated cell cultures at 6 and 12-hour time points.
Interestingly, after 12 h of hypoxia, VD3-treated cells now had the greatest relative luminescence of ATP assays, 29.3% greater than pretreated cells (Fig. 2D). Pretreated cells released significantly less NO than both untreated and treated cells after 6 h (P < 0.05, P < 0.01) and 12 h of hypoxia (Fig. 2E; P < 0.05, P < 0.01). Interestingly, pretreated cells had significantly lower levels of TMRM normalized to DAPI, indicative of mitochondrial oxidative phosphorylation, than untreated and treated cells after 6 h of hypoxia (Fig. 2F; P < 0.05, P < 0.05). By 12 h of hypoxia, all groups had similar levels of mitochondrial activity.
High-dose, inactive VD3 improved graft weight and volume retention equivalently to calcitriol
Given promising in vitro results with VD3, we next investigated in vivo xenograft retention in a murine model with calcitriol and dose-increasing VD3 treatment. After 12 weeks in vivo, grafts treated with calcitriol and 5,000 ng of VD3 macroscopically appeared qualitatively more viable than other treatment groups or controls (Fig. 3). Upon excision and weighing, calcitriol and 5,000 ng VD3-treated grafts had significantly greater weights than control grafts (P < 0.02, P < 0.05; Fig. 4A). Calcitriol and 5,000 ng VD3-treated grafts also had significantly higher volumes than control grafts (P < 0.02, P < 0.05; Fig. 4B). Grafts treated with 50 and 500 ng of VD3 did not differ significantly in either weight or volume from control grafts.
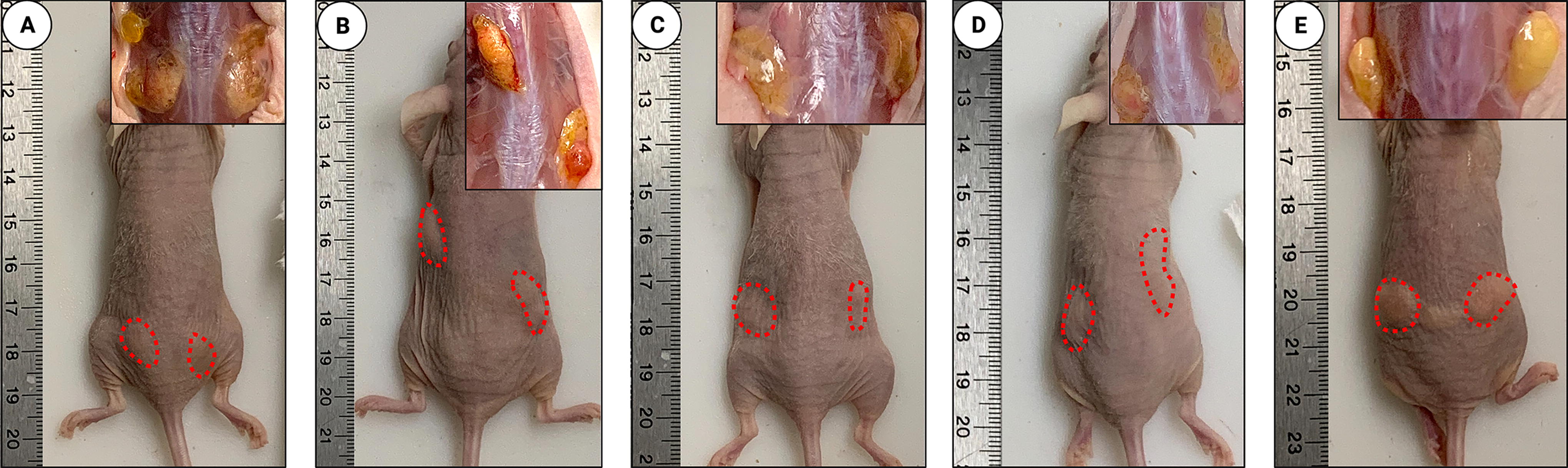
Macroscopic visualization of grafts after 12 weeks in vivo.
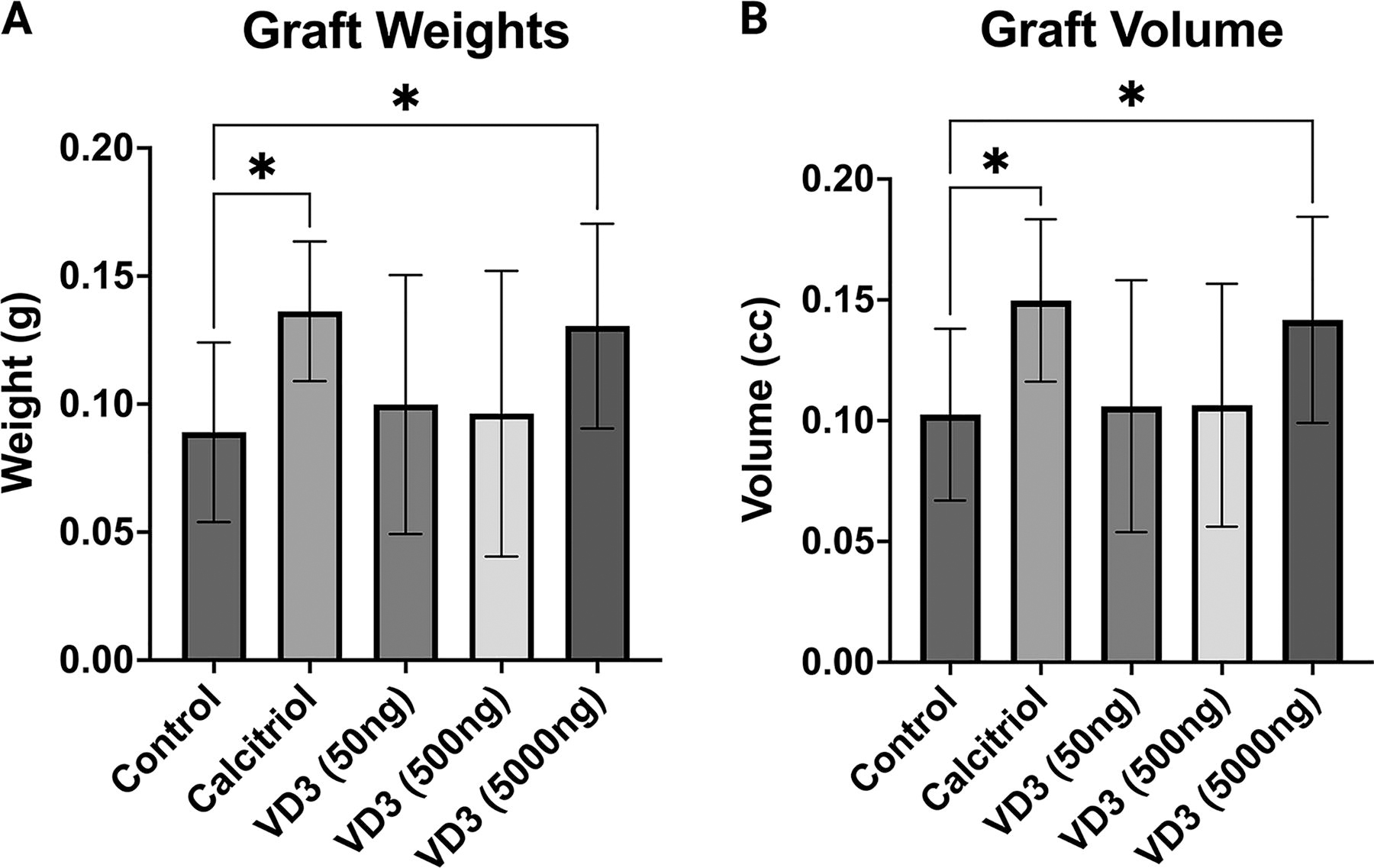
Graft retention.
Calcitriol and high-dose VD3-treated grafts demonstrated enhanced vascularity and adipocyte content
CD31 and perilipin staining were observed with immunohistochemistry to highlight neovasculature and adipocyte survival in xenografts excised after 12 weeks in vivo. Control grafts showed minimal neovasculature via CD31 staining, a high prevalence of oil cysts, and an absence of adipocyte survival with perilipin staining (Figs. 5A and 6A). Grafts treated with 50 and 500 ngVD3 demonstrated improved angiogenesis in a dose-dependent manner but maintained presence of oil cysts (Fig. 5C and D). However, oil cysts were reduced in size and incidence with increasing dosage. Furthermore, adipocyte survival visualized with perilipin showed increased prevalence compared with controls, though less than the grafts treated with calcitriol and 5,000 ng VD3 (P = 0.067, P > 0.99; Fig. 6C and D). Animals receiving calcitriol and 5,000 ng VD3 had significantly improved adipocyte architecture and increased angiogenesis compared with controls (Fig. 5B and E), as well as significantly greater adipocyte survival by perilipin staining compared with control grafts (P < 0.01, P < 0.05; Fig. 6B and E).

CD31 immunohistochemistry with asterisks denoting oil cysts and arrows highlighting neovasculature.
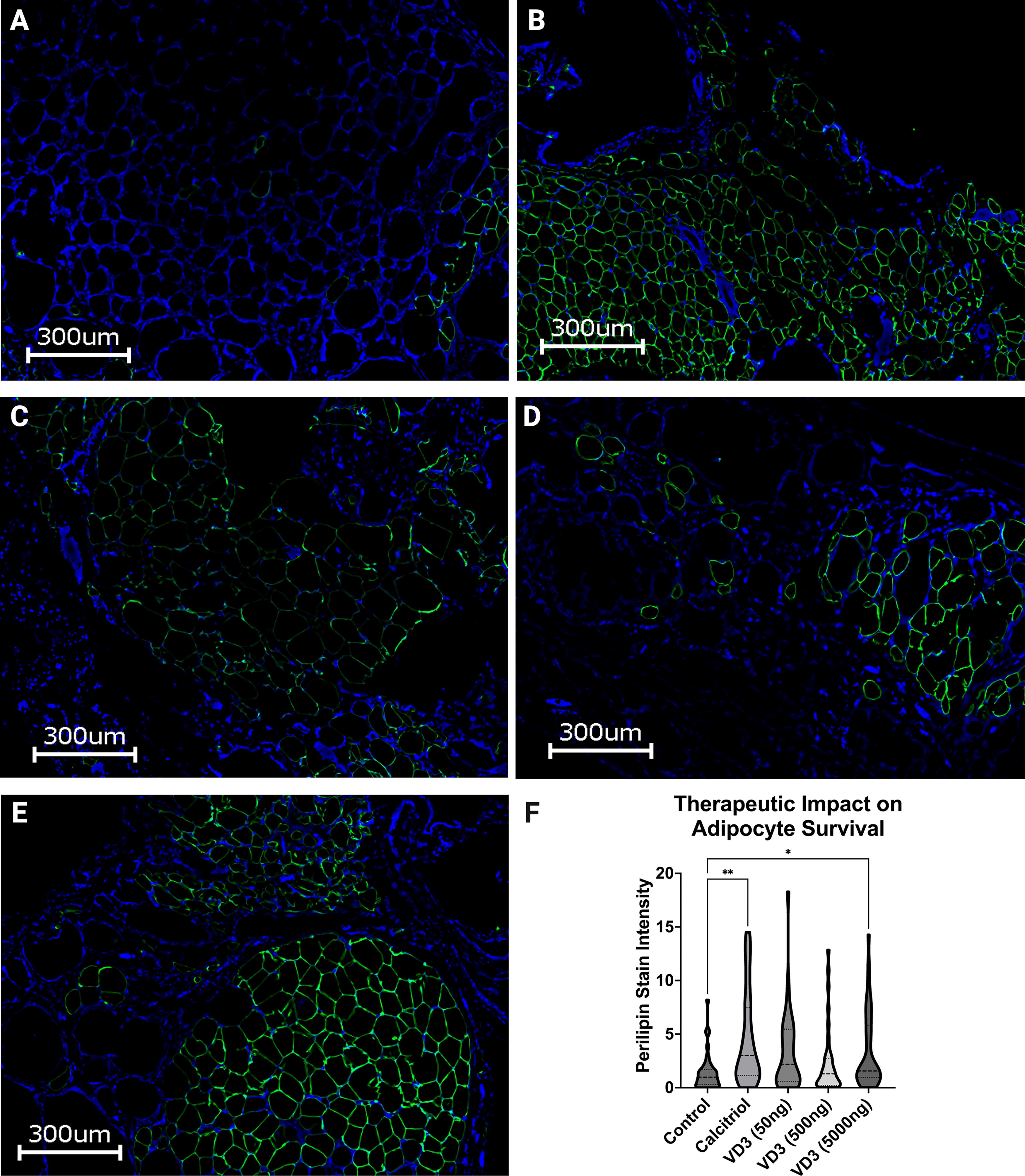
Perilipin (PLN1) stain.
Discussion
Vitamin D functions as an essential hormone, balancing innate immune activation with attenuated hyperinflammatory responses, making it a promising candidate for medically improving fat graft retention. Given the physiological risks of long-term, high-dose calcitriol administration required to improve fat grafting outcomes, we investigated the effects of inactive VD3 on adipose graft survival in whole fat particle culture, ASC culture, and our established xenograft mouse model. 11 Although vitamin D3 intermediates can have some degree of activity if able to partially bind with a target ligand, the active form, calcitriol, is significantly more potent. Mice can metabolize hormonally inactive VD3 to its active form through typical pathways involving hydroxylases in the liver and kidney, and high-dose VD3 administration showed comparable results to those obtained with calcitriol, including significantly greater graft weight and volume, upregulated neovascularization, survival of adipocytes, and minimization of oil cyst development compared with control.
Mounting research has suggested that adipose may play an active role in enzymatically metabolizing vitamin D3 from cholecalciferol to calcitriol. 18 –20 This may explain our results showing that inactive cholecalciferol significantly enhanced stromal cell viability of adipose particle cultures. Furthermore, VD3 intermediates have some degree of activity if able to partially bind with a target ligand, although calcitriol is significantly more potent. Finally, vitamin D metabolites and derivatives are potent plasma membrane stabilizers through hydrophobic interaction between the sterol rings and membrane phospholipids, exerting a fluidizing effect dependent on the disruption of packing around the side chains. 21 As our previous studies had shown that calcitriol increased adipose stem cell, endothelial cell, and pericyte survival in hypoxic cultures, and ultimately improved fat graft retention in a xenogeneic model, this study sought to determine if hormonally inactive VD3 was similarly beneficial and could be therapeutically used in lieu of the FDA-regulated drug.
Our studies suggest that ASCs pretreated with VD3 had reduced mitochondrial polarization and NO release in hypoxic culture conditions. Furthermore, pretreated cells had significantly reduced viability, significantly elevated measurement of caspases 3 and 7, and no significant differences measured for extracellular ATP, an indication of cell lysis. Taken together, these results suggest pretreated adipose stromal cells were more prone to apoptosis during hypoxic events than control cells or cells treated with VD3 at the time hypoxia was initiated. These findings reflect abundant literature relating to the use of vitamin D3, or its derivates, in the context of cancer, whereby VD3 causes tumor cell cycle arrest followed by the induction of apoptosis. 22 –24 Mechanistically, VD3 and molecular derivates thereof induce apoptosis by releasing calcium stores from the endoplasmic reticulum, which elevates intracellular calcium levels. Under aerobic conditions, healthy cells have abundant ATP-dependent mechanisms to restore intracellular calcium levels to baseline values, including plasma membrane calcium pumps, such as the plasma membrane calcium ATPase, to actively pump calcium ions out of the cytoplasm and back into the extracellular space, and sarco/endoplasmic reticulum calcium ATPase pumps that actively transport calcium ions from the cytoplasm into the ER lumen. When cells are unable to generate sufficient ATP, either owing to hypoxic conditions or in mitochondrially damaged cell populations, cells attempt to restore calcium homeostasis through energy-independent mechanisms such as mitochondrial calcium uniporters that facilitate calcium uptake into the mitochondrial matrix and/or mitochondrial sodium–calcium exchanger, which exchanges calcium ions for sodium ions across the mitochondrial inner membrane. These latter calcium balancing mechanisms disrupt the polarity of the mitochondrial membrane, diminishing the mitochondrial membrane potential (ΔΨ), 25 and impair mitochondrial oxidative phosphorylation metabolism. 26,27 As a result, VD3 pretreated cells under hypoxic conditions release apoptotic-inducing factors leading to calpain or caspase activation, depending on the cell type. 28
The observation that VD3 treatment significantly reduced NO production in both experimental groups, treated and pretreated, was unexpected and did not reflect observed trends in cell viability or apoptosis. For example, multiple publications have reported that VD3 and its derivatives significantly increase NO production in multiple cell types, 29 –31 typically instantaneously, with peak NO production measured within the first 10 s of VD3 exposure. 32 However, the impact of VD3 on NO production appears to be biphasic, with longer exposure (e.g., 12 h) causing a genetic downregulated expression of NADPH oxidase, eNOS, and iNOS, reducing production of NO. 32 Although we did not measure the expression levels of any gene related to NO production, our results reflect mechanistic data from longer term exposure as we measured NO production with 6, 12, and 18 h (for pretreated samples) of VD3 exposure.
The potential finding presented herein that VD3 pretreated cells may be prone to apoptotic cell death mechanisms during hypoxia has interesting implications for postgrafting interactions with infiltrating immune cells as apoptosis is recognized as inducing tissue regenerative responses, 33,34 whereas necroptosis 35 or ferroptosis 36 typically induce more inflammatory outcomes. However, immune response to autologous fat grafting is a complex sequence of events, shaped by its microenvironments, which is difficult to predict with an in vitro model. A delicate interplay between cell survival, mechanism of eventual death and immune response ultimately determines graft volume and composition. The use of nude mice limits the modeling of an immune response owing to their lack of an adaptive immune system until 12 weeks of age, after which they mount an antihuman immune response to xenografts, potentially compromising the quality of xenografts and appropriate quantification of xenograft retention. Lastly, without collecting blood samples from mice, analysis of VD3 conversion to a stable metabolite is limited. Consequently, evaluating dosage is also limited. Future studies are needed to determine optimal VD3 timing and dosage for graft retention as well as understand the critical mechanisms which facilitate graft survival or replacement.
Footnotes
Author Disclosure Statement
None of the authors have any financial interests to disclose.
Funding Information
This work was funded by Plastic Surgery Foundation National Endowment Award for Plastic Surgery #410329. The University of Pittsburgh holds a Physician-Scientist Institutional Award from the Burroughs Wellcome Fund supporting Dr Loder.