
Abstract
Radiation therapy (RT) is a typical treatment for head and neck cancers. However, prolonged irradiation of the esophagus can cause esophageal fibrosis due to increased reactive oxygen species and proinflammatory cytokines. The objective of this study was to determine whether myogenic gene-transfected mesenchymal stem cells (MSCs) could ameliorate damage to esophageal muscles in a mouse model of radiation-induced esophageal fibrosis. We cloned esophageal myogenic genes (MyoD, MyoG, and Myf6) using plasmid DNA. Afterward, myogenic genes were transfected into Human Mesenchymal Stem Cells (hMSCs) using electroporation. Gene transfer efficiency, stemness, and myogenic gene profile were examined using flow cytometry, quantitative polymerase chain reaction, and RNA sequencing. In vivo efficacy of gene-transfected hMSCs was demonstrated through histological and gene expression analyses using a radiation-induced esophageal fibrosis animal model. We have confirmed that the gene transfer efficiency was high (∼75%). Pluripotency levels in gene-transfected MSCs were significantly decreased compared with those in the control (vector). Particularly, myogenesis-related genes such as OAS2, OAS3, and HSPA1A were overexpressed in the group transfected with three genes. At 4 weeks after injection, it was found that thickness collagen layer and esophageal muscle in MSCs transfected with all three genes were significantly reduced compared to those in the saline group. Muscularis mucosa was observed prominently in the gene combination group. Moreover, expression levels of myogenin, Myf6, calponin, and SM22α known to be specific markers of esophageal muscles tended to increase in the group transfected with three genes. Therefore, using gene-transfected MSCs has the potential as a promising therapy against radiation-induced esophageal fibrosis.
Introduction
For most head and neck cancers including upper digestive cancers, radiation therapy (RT) is incorporated as a standard treatment, which is performed in parallel with surgical therapy. 1 RT for head and neck cancer can expose normal bones and soft tissues of the jaw, face, and neck to radiation, causing tissue damage known as radiation fibrosis. 2,3 Any soft tissue can be affected, including skin, connective tissue, muscles, nerves, and blood vessels. A representative side effect occurring after RT of the upper digestive tract, such as oral, pharyngeal, or esophageal tissue, is the swallowing disorder. 4 This is a functional disorder due to tissue fibrosis of the mucous membrane and muscle layer of the esophagus. Apoptosis of tissue stem cells by ionizing radiation, cytokines, and chemokines released that can cause inflammatory responses and fibroblast activation is a main contributor to radiation-induced fibrosis. 5,6 As a result, in tissues exposed to radiation, excessive deposition of collagen and other extracellular matrix components can occur, leaving permanent tissue scarring.
Most studies on tissue fibrosis that occurs after RT are in the field of direct stem cell transplantation. Studies on inhibition of fibrosis by secretion of various cytokines (e.g., KGF, fibroblast growth factor [bFGF], Hepatocyte Growth Factor (HGF), Matrix Metalloproteinase (MMP), and so on) through a paracrine effect of stem cell transplantation have been reported. 7 –9 Despite these effects, stem cells have a low survival rate in tissues. They are vulnerable to various immune responses. Therefore, several medical technologies are being applied nowadays to overcome shortcomings of stem cells.
On the contrary, gene therapy is a technology that involves genetically modifying patient’s cells to treat and improve disease. In detail, this is a technology that loads a healthy therapeutic gene into a vector and delivers it to target cells to replace mutated and degenerated genes. 10 The delivery vector mostly used is a viral vector (adenovirus, retrovirus, etc.) with a high transduction efficiency. 11 However, in recent years, a nonviral vector (liposome, plasmid DNA, electroporation, etc.) has been developed to improve the safety of gene therapy. 12 –14 For example, Professor Lee and his colleague used plasmid DNA to transfect bFGF into vocal fold fibroblasts. 15 Next, they validated the efficiency of the gene delivery by injecting this into a vocal fold palsy animal model. Other works include transfection of the microdystrophin gene into skeletal muscle cells using peptide-functionalized dendrimer nanocarriers by Professor Deo’s team at the University of Miami School of Medicine. 16 These cells were then applied to a Duchenne muscular dystrophy animal model for evaluation of the therapeutic efficacy.
In this research, we hypothesized that the ultimate treatment of damaged esophageal tissue might be possible by restoring major genes of the damaged esophageal muscle layer after irradiation. Esophageal muscles are the most important target for fundamental restoration of the function of the esophagus. 17 Therefore, we designed an animal model based on previous studies in which we induced fibrosis by irradiating the esophageal muscle and mucosal layer. Tissues that were exposed to ionizing radiation, hydroxyl radicals are formed by reacting with intracellular water molecules, which lead to DNA damage in muscle cells. 18 This then resulted in the increased production of activated TGF-β, which resulted in decreased tissue oxygenation, leading to tissue ulceration. 19 In addition, we expected that the esophageal function could be restored through modulation of related genes after esophageal muscle fibrosis. Transcriptional factors such as Mrf-4,5,6, MyoD, and myogenin are recognized as important factors in the degenerated esophageal muscle layer in many studies. 20 –23
We selected MyoD, myogenin (MyoG), and Myf6 genes and cloned them into a nonviral vector to restore genetic modification due to DNA destruction in esophageal muscle cells after irradiation. Using electroporation, a vector incorporated with mesenchymal stem cell (MSC) genes was transfected. This research aimed to verify the efficacy of gene therapy after esophagus after transplanting transgenic stem cells into animal models with induced irradiation fibrosis.
Materials and Methods
Preparation of MSCs
BMMSCs were purchased from Lonza (Basal, Switzerland). BMMSCs were cultured with Dulbecco’s modified eagle medium (DMEM, LONZA, BioWhittakerTM, USA) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin. Cell culture medium was refreshed every 2 days. For subculture, 0.05% Trypsin-EDTA (1X, Phenol Red, Gibco, USA) was used for detaching cells. BMMSCs were then expanded until passage 6 for gene transfection. BMMSCs were characterized using cell surface markers by flow cytometry (FACS) analysis (positive: CD105, CD90, CD73, CD44; negative: CD34, CD11b).
Cloning of MyoD, MyoG, Myf6, and a combination of three genes using plasmids
To amplify MyoD, MyoG, Myf6 genes, clones were purchased from Addgene (MA, USA). Each gene was amplified using polymerase chain reaction (PCR). Amplified genes were applied in cloning using a pEGFP-C1 expression vector either independently as MyoD, MyoG, Myf6 or all at the same time as 3-gene combination that combined MyoD, MyoG, and Myf6. The cloning was commissioned to BIONICS (Seoul, South Korea).
Gene transfection using electroporation technique
For transfection of BMMSCs with MyoD, MyoG, Myf6, or 3-gene combination, Neon™ Transfection System (Invitrogen™, Neon™, USA) was used. After harvesting subconfluent MSCs, 1 × 106 cells were resuspended in the R buffer provided by the manufacturer, to which 2 μg of constructed DNA was added. Through repeated tests, optimal conditions for transfection of BMMSCs with MyoD, MyoG, Myf6, or 3-gene com plasmid were set (900 voltage (V), 40 width (ms), and 2 pulses. Gene transfection efficiency was determined by fluorescence microscopy (LEICA, DMI4000B, and BD LSR II flow cytometry) at 48 h post transfection. The efficiency of gene transfection was confirmed using real-time PCR.
Myogenic differentiation and immunofluorescent staining
For induction of myogenesis, in vitro cell culture was carried out using 5 × 105 naked MSCs or each gene-transfected MSCs at passage 5 in myogenic medium (high-glucose DMEM supplemented with 5% horse serum, 100 mM dexamethasone, and 10 ng/mL basic FGF). This medium was exchanged every 3 days for the remainder of the culture. Two weeks after inducing myogenic differentiation, cells were immune stained for myogenin and calponin and visualized with a fluorescence microscope. Cells were then fixed, permeabilized, blocked, and sequentially incubated with mouse anti-myogenin antibody (1:200; Santa Cruz Biotechnology) and rabbit monoclonal anti-calponin (1:200; Santa Cruz Biotechnology). Secondary antibodies against myogenin and calponin were Alexa Fluor® 596 goat anti-mouse (Invitrogen) and Alexa Fluor 488® goat anti-rabbit IgG antibodies (Abcam, UK), respectively. Nuclei were stained with 4,6-diamidino-2-phenylindole (DAPI; Vector Laboratories). Images were obtained via confocal microscopy (LSM510 META; Carl Zeiss, Germany). For quantification of positive cells, gene transfection was performed five times, and five replicates of immunocytochemistry staining were conducted. For each stained slide, five images were taken from different regions, and the percentage of positive cells were calculated by counting them together with the repeated experiments.
RNA isolation from transfected BMMSCs
After DNA-transfected BMMSCs were cultured for two days, cells were detached and lysed with QIAzol lysis reagent (Qiagen, Hilden, Germany). After lysis, RNA of each tissue sample was isolated using an RNeasy kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol.
RNA sequencing
Isolated total RNA was used for preparing mRNA sequencing library with an Illumina TruSeq Stranded mRNA Sample Preparation kit (Illumina) according to the manufacturer’s protocol. Quality and size of libraries were assessed using an Agilent 2100 bioanalyzer DNA kit (Agilent). All libraries were quantified by quantitative polymerase chain reaction (qPCR) using a CFX96 Real Time System (BioRad) and sequenced on NextSeq500 sequencers (Illumina) with a paired-end 75 bp plus single 8 bp index read run. To compare expression profiles among samples, normalized expression values of a few hundred of differentially expressed genes selected were unsupervised clustered with in-house R scripts. Scatter plots for gene expression values and volcano plots for expression-fold changes and p-values between two selected samples were also drawn with in-house R scripts.
Functional category analysis
To get insights on the biological function of differential gene expression between compared biological conditions, gene set overlapping test between analyzed differential expressed genes and functional categorized genes were performed, including biological processes of Gene Ontology, KEGG pathways, and transcription factor binding target gene sets using g:Profiler2 version 0.2.0.
Real-Time quantitative polymerase chain reaction (RT-PCR)
RNAs isolated from individual DNA samples were used to synthesize cDNAs with a high-capacity RNA-to-cDNA™ Kit (Applied Biosystems™, USA) following the manufacturer’s protocol. RNA levels were determined by quantitative real-time PCR using a LightCycler® 480 SYBR Green I Master (Roche, Swiss) following the manufacturer’s protocol. The PCR conditions were set at 30 cycles of target gene amplification, with a pre-denaturation at 95°C for 5 min, followed by denaturation at 95°C for 15 s and annealing at 60°C for 30 s. The relative expression of each target gene was calculated using the comparative ΔΔCt method and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression was used for normalization. Primers used in this study are listed in Supplementary Table S1.
For real-time PCR analysis, each gene transfection was performed a total of five times, and RNA was isolated for use in the experiments.
In vivo animal studies
Animal model of irradiation-induced damage to the esophagus. All animal procedures were implemented according to guidelines of the Institutional Animal Care and Use Committee of Seoul National University Hospital (approval number: 22-0039-S1A0). Animal model for fibrosis-induced esophagus was prepared using 8-week-old C57BL/6 mice (orientbio, Korea). A total of 48 male mice were divided into six groups: (1) normal, (2) saline injection (control), (3) injection of MyoD-transfected MSC, (4) injection of MyoG-transfected MSC, (5) injection of Myf6-transfected MSC, and (6) injection of MyoD+MyoG+Myf6 transfected MSC. Experimental mice were anesthetized via intraperitoneal injection with 5 mg/kg xylazine and 20 mg/kg Zoletil 50. Neck hair was shaved for each mouse. Mice were placed in a supine position. Irradiation was carried out using a 6-MV X-ray from a Truebeam linear accelerator (Varian Medical Systems, Palo Alto, CA, USA), at a dose rate of 750 MU focused on the neck skin from a distance of 10 cm (Fig. 3A). 24 X-ray irradiation was conducted twice a week with a total dose of 10 Gy.
Administration of MSC encoded with myogenic genes into the esophagus. At day 1 after the second irradiation, mice were anesthetized via intraperitoneal injection with 5 mg/kg xylazine and 20 mg/kg Zoletil 50. They were then placed in the supine position with surgical sites disinfected with betadine for aseptic surgery. A ventral midline incision was made. The strap muscle was divided to isolate the tracheoesophageal structure. Following an appropriate exposure of the cervical esophagus under a microscope, gene-transfected MSCs were injected (20 μl) into the esophageal muscle using a 100 μl syringe (26-gauge needle; Hamilton, Reno, NV, USA) (Fig. 3B and C). After closing the muscle and skin tissue with 5–0 Vicryl suture, each experimental mouse was subcutaneously injected with 1.6 mg/kg of meloxicam for analgesic effect. Postoperative monitoring was conducted daily for clinical signs of distress such as weight loss, spasms, dyspnea, or consciousness disorders. At 4 weeks post-injection, the efficacy of the gene-transfected MSCs treatment was confirmed by histological evaluation and qPCR analysis.
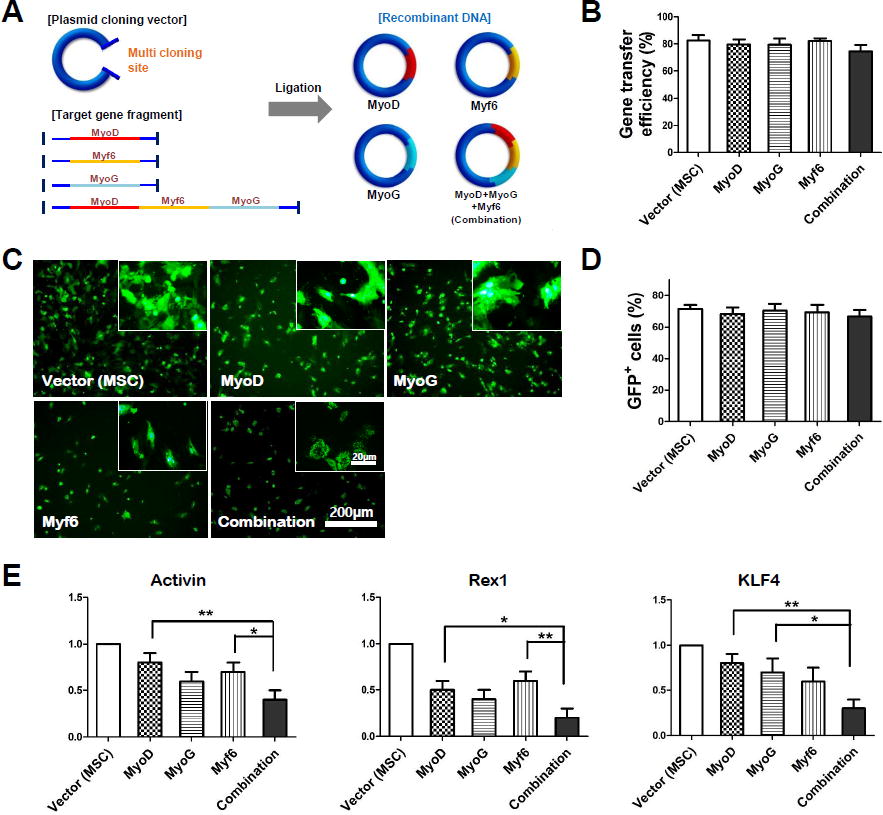
Gene encoding and transfection efficiency using plasmid cloning vector.
In vivo tracking of gene-transfected MSCs in injected site. To detect PKH-26-labeled MSCs around the esophageal muscle, gene-transfected MSCs were labeled with a PKH26 Red Fluorescent Cell Linker Kit (Sigma-Aldrich) solution according to the manufacturer’s instructions. Labeled cells (1 x 106/mL, PBS) were injected into esophageal muscles of irradiated mice. At one week after injection, tissue samples were frozen using 2-methybutane, an optimal cutting temperature compound (Tissue-Tek, USA), and sectioned at 4 μm in thickness with a cryotome (Leica CM1900, Germany). To minimize the decrease in fluorescence intensity during the immune-staining process, cryo-sectioned slides were immediately observed under a fluorescence microscope (BX43, Olympus, Japan).
qPCR using esophageal tissues. The esophageal tissue (three rats from each group) was removed at 4 weeks after injection with gene-transfected MSCs (MyoD, MyoG, Myf6, and Combination). Gene expression level was assessed by qPCR. Total RNA was isolated from homogenized esophageal tissues using Qiazol® Lysis reagent (QIAGEN sciences, MD, USA). Then 1 μg of total RNA was used to synthesize cDNA with a LeGene Premium Express First Strand cDNA Synthesis System (LeGene Bioscience, USA) according to the manufacturer’s protocol. qPCR assays were then performed using a Power SYBR Green PCR Master mix (Thermo Fisher Scientific, USA) and a Step One Plus RT-PCR System (Thermo Fisher Scientific, USA). For quantification, GAPDH was used as an internal control. Primer sequences are listed in Supplementary Table S1.
Histological examination. Four weeks post-injection, four rats from each group were euthanized and fixed in 10% neutral-buffered formalin. Tissue samples were dehydrated, embedded in paraffin blocks, and sectioned at a thickness of 4 μm. These sections were then deparaffinized and dehydrated in a graded series of ethanol. Tissue sections were then stained with hematoxylin and eosin (H&E) and Masson’s trichrome staining according to the manufacturer’s instructions. Histological images from each sample were obtained in triplicate using a light microscope (Olympus, Japan) at ⅹ200 magnification. The distribution of vacuolization in the submucosa was calculated as a percentage using H&E staining. The average value was calculated from six images at 400x magnification (high-power field) per slide. Subsequently, the thickness of collagen positive layer by Masson’s trichrome staining (blue color) was calculated using ImageJ tool (NIH; n = 5 per group). The average value was obtained from four images per tissue section. To guarantee the reliability of the results, all of the samples were processed in triplicate.
Immunohistological analysis. Specimens were immersed in 3% hydrogen peroxide (H2O2) in methanol for 20 min to inactivate endogenous peroxidase. Slides were then rinsed with PBS and incubated with 3% BSA for 1 h to block nonspecific sites. Tissue samples were then incubated with a rabbit polyclonal desmin antibody (Catalogue # ab8592, Abcam, UK) followed by incubation with a secondary antibody (Goat Anti-Rabbit IgG Alexa Fluor® 488, Catalogue # ab150077, Abcam, UK) at 4°C overnight. For quantitative analysis, the total thickness of the muscle layer expressed in green was calculated using the ruler tool in ImageJ (n = 4 per group). The average value was calculated by randomly measuring four images per tissue section. Tissue sections for CD68 (mouse monoclonal antibody, Abcam, ab955) were then incubated using a horseradish peroxidase-conjugated kit (Vectastain). They were then visualized using a chromogenic substrate 3,3'-Diaminobenzidine (DAB) (Vector, Catalogue # PK-7800, Vector Laboratories, USA). Cell nuclei were counterstained with hematoxylin. Histological images were captured using a fluorescence microscope (BX43, Olympus, Japan). The number of CD68-positive cells was counted from five different fields (200× magnification) (n = 6 per group). Regeneration of muscularis mucosa in the submucosa layer was evaluated via immunofluorescence staining against α-Smooth Muscle Actin (SMA). Primary antibodies used were mouse monoclonal anti-α-SMA (Catalogue # 14–9760-82, Invitrogen). Secondary antibodies were Alexa Fluor 488 goat anti-mouse IgG (Invitrogen). The fluorescent area (%) of α-SMA was calculated using ImageJ software by measuring the area of muscularis mucosa expressed in green relative to the total area (n = 4 per group). The average value was obtained from four different fields per sample.
Statistical analysis
Statistical significance was determined using one-way analysis of variance. When appropriate, Tukey–Kramer’s post hoc test was also used. All statistical analyses were performed using GraphPad Prism 5 software (La Jolla, CA, USA). Data are expressed as means ± standard deviation. *P < 0.05, **P < 0.01, and ***P < 0.001 were considered statistically significant.
Results
Generation of nonviral constructs based on a microporation system
MyoD, MyoG, Myf6, and combination (MyoD+MyoG+Myf6) known to be key regulators of myogenesis were inserted into the multicloning site of the pEGFP-N1 vector expressing GFP (Fig. 1A). BMMSCs used for gene cloning were found to be positive for MSC surface markers CD105, CD90, CD73, and CD44 but negative for hematopoietic markers CD34 and CD11b (Supplementary Figure S1). Flow cytometry analysis results confirmed that gene transfection efficiency was maintained over 75% in all groups (Fig. 1B). The transfection efficiency of vectors and MyoD, MyoG, Myf6, and Myo-combination genes was determined by counting GFP-positive cells using a fluorescence microscope. In the magnified image, GFP was clearly observed in the entire cytoplasm (Fig. 1C). The vector group showed the highest transfection efficiency at 73.7%, while the MyoD, MyoG, Myf6, and Myo-combination groups exhibited efficiencies of 70.5%, 72.4%, 72.3%, and 69.5%, respectively (Fig. 1D). Although there were slight differences in transfection efficiency, they were not significant, and these results were similar to those obtained by flow cytometry. This suggests that genetic transformation of MSC was successfully performed using a physical method of microporation.
Pluripotency and myogenic potential of gene-transfected MSCs
After gene transfection of hMSCs, real-time PCR was performed for stem cell pluripotency using pluripotent markers Activin, Rex1, and KLF4 (Fig. 1E). As a result, the same trend was confirmed for all three types of markers. Pluripotent gene expression in single gene-transfected group was decreased compared to that in the vector (MSC) group. Specifically, for KLF4 and Rex1 groups, the group transfected with their combination showed a significant decrease compared to other single-transfected groups. Myogenin and Calponin are commonly used as markers to assess phenotype of myogenic and smooth muscle cell (SMC) lineage. They were analyzed by immunocytochemistry (Fig. 2A). As a result, myogenin showed a distinct phenotype in the group transfected with all gene compared with the vector group. In addition, the calponin marker showed a phenotype of SMC differentiation in myf6 and combination groups, meaning that the transfer of myogenic genes had a positive effect on SMC differentiation. As a result of quantitative analysis, the possibility of differentiation into a more meaningful muscle lineage was suggested in the group in which all three genes were transfected (Fig. 2B).

Confocal microscope images showing MSC differentiation via myogenic cultivation.

The myogenic potential of gene-transferred MSCs through myogenic markers was then analyzed through real-time PCR (Fig. 2C). Most gene-transfected groups showed significant differences in all markers compared to the saline group. Notably, Pax7, a Myogenin marker, showed a significant increase in the combination group compared with myoD and MyoG groups. In addition, the Desmin gene showed a statistical difference in the combination group compared with the other three groups. These results demonstrated that muscle differentiation of MSCs was further improved in the group transfected with a combination of three genes compared with that in the group with a single gene transfection. Based on the expression of these genes, co-transfection of MyoD, MyoG, and Myf6 genes might help enhance myogenesis.
Cell tracking and histological analysis of 4 weeks after injection
In vivo cell tracking post-injection in the esophageal muscle layer was visually observed using PHK-26-labeled MSCs. Representative image for in vivo tracking of gene-transfected MSCs (combination group) are shown in Supplementary Figure S2. A large cluster of injected hMSCs were located in the esophageal muscle layer with a red color (yellow arrows). At 4 weeks after injection, the entire esophagus was observed after H&E staining (Supplementary Figure S3). In the saline group, unlike the normal esophagus, the lumen was irregular, and the submucosal layer was thickened asymmetrically. Irradiation of the esophagus resulted in significant deformation of the esophageal tissue. However, in the group injected with transgenic MSCs, deformation of the esophagus was clearly suppressed. In more detail, in groups injected with MSCs transfected with different genes, cytoplasmic vacuolization (yellow arrow) in the epithelium were observed more intermittently compared to the saline group (Fig. 3D). Quantitative analysis revealed that the combination group showed significantly lower average values compared to the single gene-transfected groups, with a notably greater decrease compared to the MyoD group (Fig. 3E). In addition, hyperplasia (yellow arrows) of mucosal epithelial cells was observed intermittently. However, in the combination group, epithelial cell hyperplasia was hardly observed. It was confirmed that recovery of the esophageal mucosa morphology in the group transfected with a combination of three genes was similar to that in the normal group.
Tissue reaction for radiation-induced esophageal fibrosis
The tissue damage to the esophageal mucosa and muscle due to radiation exposure may vary slightly between individuals. In addition, there may be differences in the retention of the injected cells within the target tissue. To minimize the data variability that could arise from these factors, we used a region of interest to be able to randomly measure the morphology of the lesions. In the case of esophageal tissue damage after irradiation, distinct pathological changes were observed in the submucosal layer (IEL). Extracellular Matrix (ECM) deposition and hypertrophy of collagen (saline group) due to penetration of fibroblasts into damaged tissue was confirmed by Masson’s trichrome staining (Fig. 4A). Such pathological changes in the submucosal layer were analyzed through quantification of the IEL thickness (Fig. 4B). As a result, in the saline group, hypertrophic proliferation of the collagen layer in the submucosal layer of the esophagus was confirmed after irradiation. On the contrary, in the group treated with MSCs transfected with genes, the collagen layer was significantly decreased compared with that in the control group. However, there was no statistical significance among the experimental groups. This is presumed to be a result of the injected MSCs inhibiting collagen synthesis in the submucosal layer. This was also verified by analyzing the expression pattern of proinflammatory cytokines (Fig. 4C). IL-1beta was significantly decreased in the three experimental groups (Myo, Myf6, combination) compared with that in the control group. In addition, for Col type 1, a statistical decrease compared with the control group was only confirmed in the combination group. More significantly, TGF-beta, a gene known to be specific for tissue fibrosis, showed a significant decrease in all four experimental groups compared with that in the saline group. This was presumed to be due to the antifibrosis effect of MSCs. 25
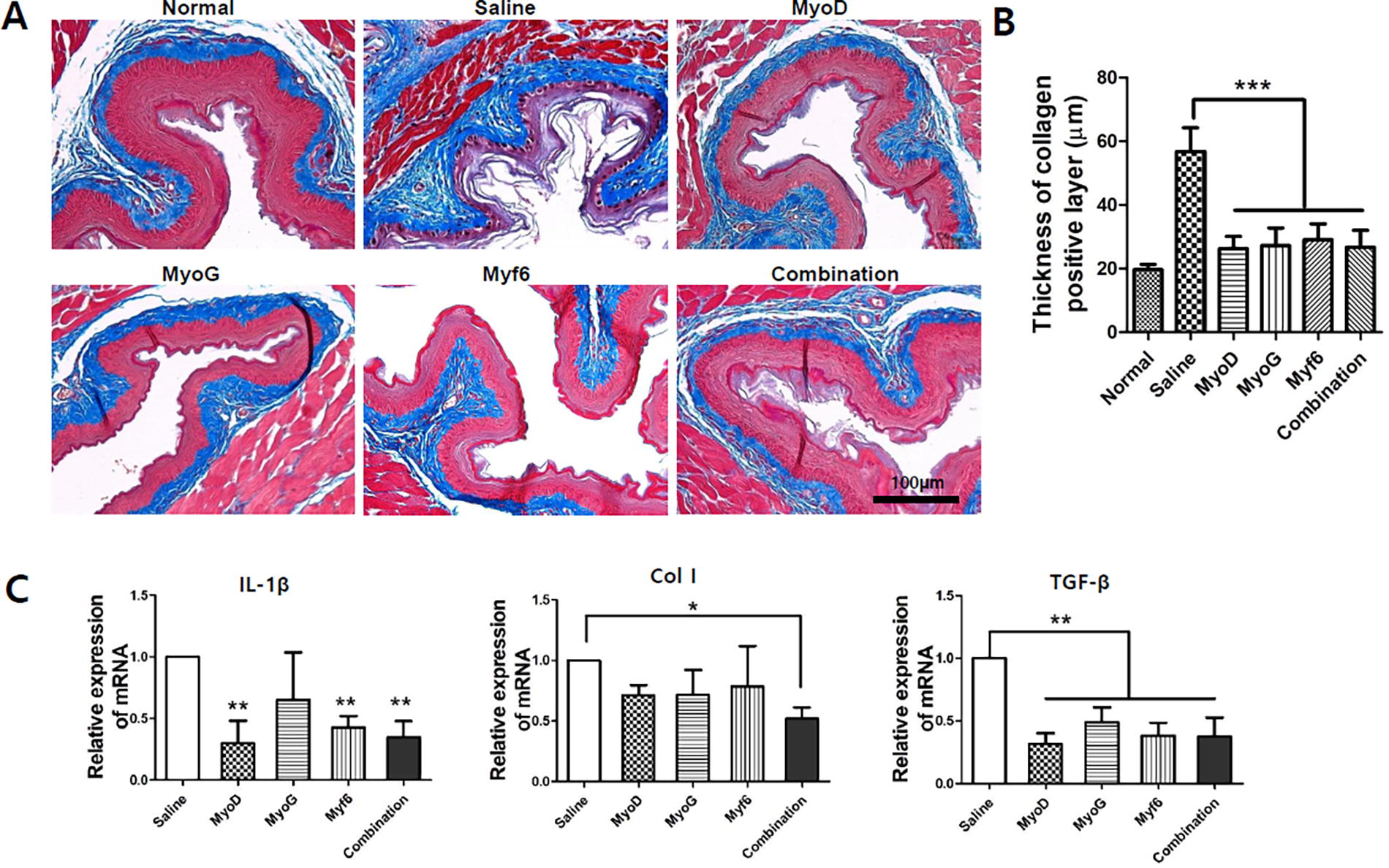
Histological evaluation for collagen deposition of inner esophageal layer after injection of gene-transfected MSCs.
Regeneration of esophageal muscle
Histological changes in the esophageal muscle layer for each group were observed through Desmin immunostaining (Fig. 5A). For the saline group, collapse of the inner circular and outer longitudinal muscle layer occurred and a hyper atrophied morphology was observed. Specifically, the muscle layer structure was normalized in the combination group. Thickness of the esophageal muscle layer was significantly reduced compared with that in the group with a single gene (Fig. 5B). Afterward, specific muscle gene expression was analyzed by qPCR (Fig. 5C). As a result, the transcriptional factor myf6 showed a similar tendency to the tissue staining result. In addition, a significant increase in myogenin was seen in the group with MyoG transfection compared with groups with transfection of the other two genes. Smooth muscle representative genes calponin and SM22α also showed a statistical difference in the combination group compared with those in groups with a single gene transfected. Notably, all gene-transfected groups showed statistical significance in calponin and myogenin compared with the saline group. This supports the finding that translocation of genes can affect the regeneration of damaged DNA in muscle genes.
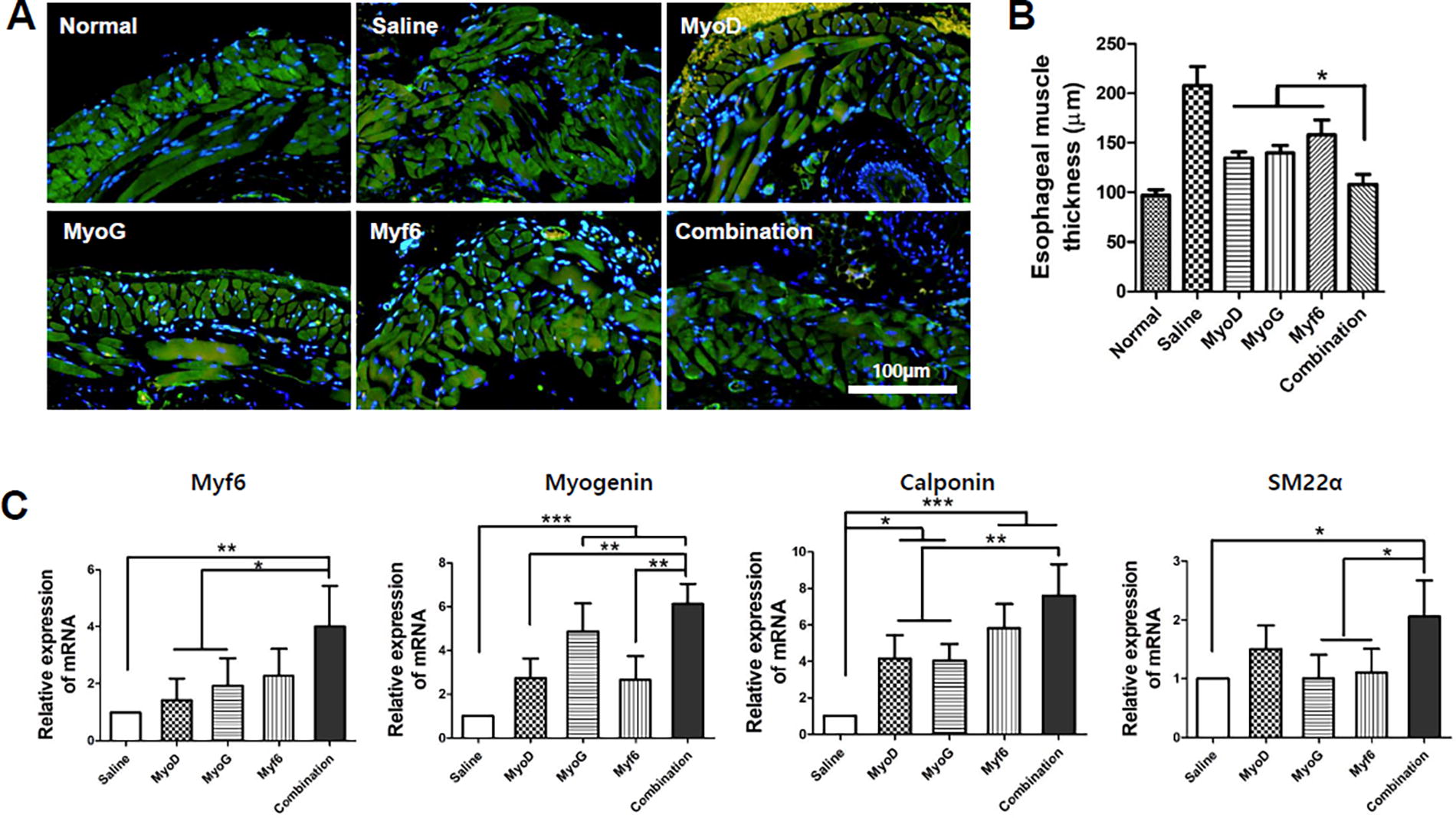
Evaluation of muscle remodeling in injected sites after administration of gene-transfected MSCs.
Macrophage expression and regeneration of muscularis mucosa
The distribution of macrophages expressed in the submucosa of each group was evaluated through CD-68 immunostaining (Fig. 6A). The macrophages are indicated by the brown color within the submucosal layer, with a pronounced expression of macrophages being observed in the saline group. This indicates an increased tissue immune response due to the ulceration within the submucosal layer caused by irradiation. In the saline group, the expression of macrophages was sharply increased at week 4. Tissue necrosis is induced by irradiation in the submucosal and muscle layers, which can lead to an increased expression of macrophages. However, macrophage expression decreased significantly in all four transgenic groups (Fig. 6B). This result was assumed to be due to the immunomodulatory effect of MSCs. No significant difference in macrophage expression was found between experimental groups.
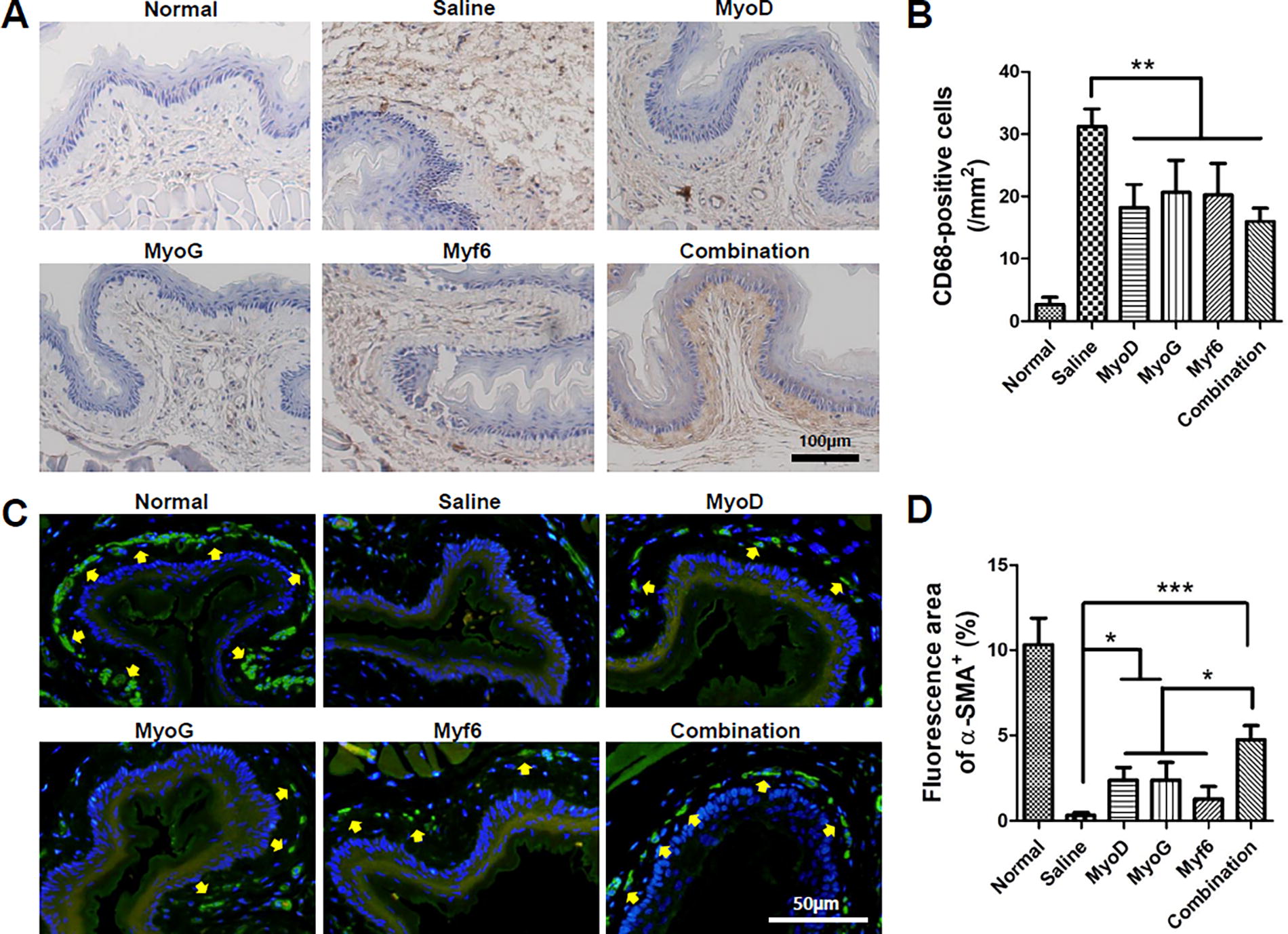
Representative image showing infiltration of macrophages by CD68 immunostaining
Changes of muscularis mucosa in the esophagus were observed through α-SMA immunostaining (Fig. 6C). In the normal group, mucosal muscularis expressed as the green color along the submucosal layer was clearly observed. The muscularis mucosa is a major subtissue involved in the functioning of the esophagus. Results are as follows. In the irradiated saline group, almost no mucosal layer was observed. On the contrary, in the gene combination group, regeneration of the muscularis mucosa was clearly observed. Quantitative analysis results showed a significant increase in the MyoD, MyoG, and combination groups compared with the saline group (Fig. 6D). Specifically, the combination group showed a significant increase in comparison to the other experimental groups. Although the average value was lower in the myf6 gene group among single-transfected groups, with no statistical difference was observed. Based on these results, it can be seen that esophageal function is clearly improved in the group treated with MSCs transfected with a combination of the three genes.
Mechanism changed by MyoD, MyoG, Myf6, and MIX gene transfection using RNA sequencing
Myogenesis enhancements in groups treated with MSCs transfected with MyoD, MyoG, Myf6 alone or a combination of the three genes were confirmed both in vitro and in vivo. In particular, improved regeneration ability of smooth muscle by such gene transfection was a very useful result. RNA sequencing was performed to determine the mechanism involved in the improvement of myogenic potential by transfecting these genes. To visualize transcriptomic differences between MyoD, MyoG, Myf6, and combination of these three genes, a heatmap was generated based on the criterion of |log2 (fold-change)| > 2 (Fig. 7A). Through the heat map, a significant difference in gene profiling was confirmed by each gene transfection. The volcano plot showed changes in gene profiling caused by MyoD, MyoG, Myf6 alone, and co-transfection of the three genes (Fig. 7B). It was possible to confirm the gene and distribution of genes under-expressed and over-expressed by each gene transfection. Among genes involved in myogenesis, USP18, ISG15, IRF7, and OAS1 were found to increase in expression in all groups, especially in the combination group. Myogenesis-related genes such as OAS2, OAS3, HSPA1A, HSPA1B, TGFA, APOL1, and SAMDHD1 were also overexpressed. In the combination group, the expression of genes related to myogenesis was much more expressed than in the group transfected by MyoD, MyoG, or Myf6 alone. The diversity of muscle-related genes expressed in the combination group provides evidence that transfection with these three genes could have a therapeutic effect on the recovery of esophageal muscles damaged by radiation.
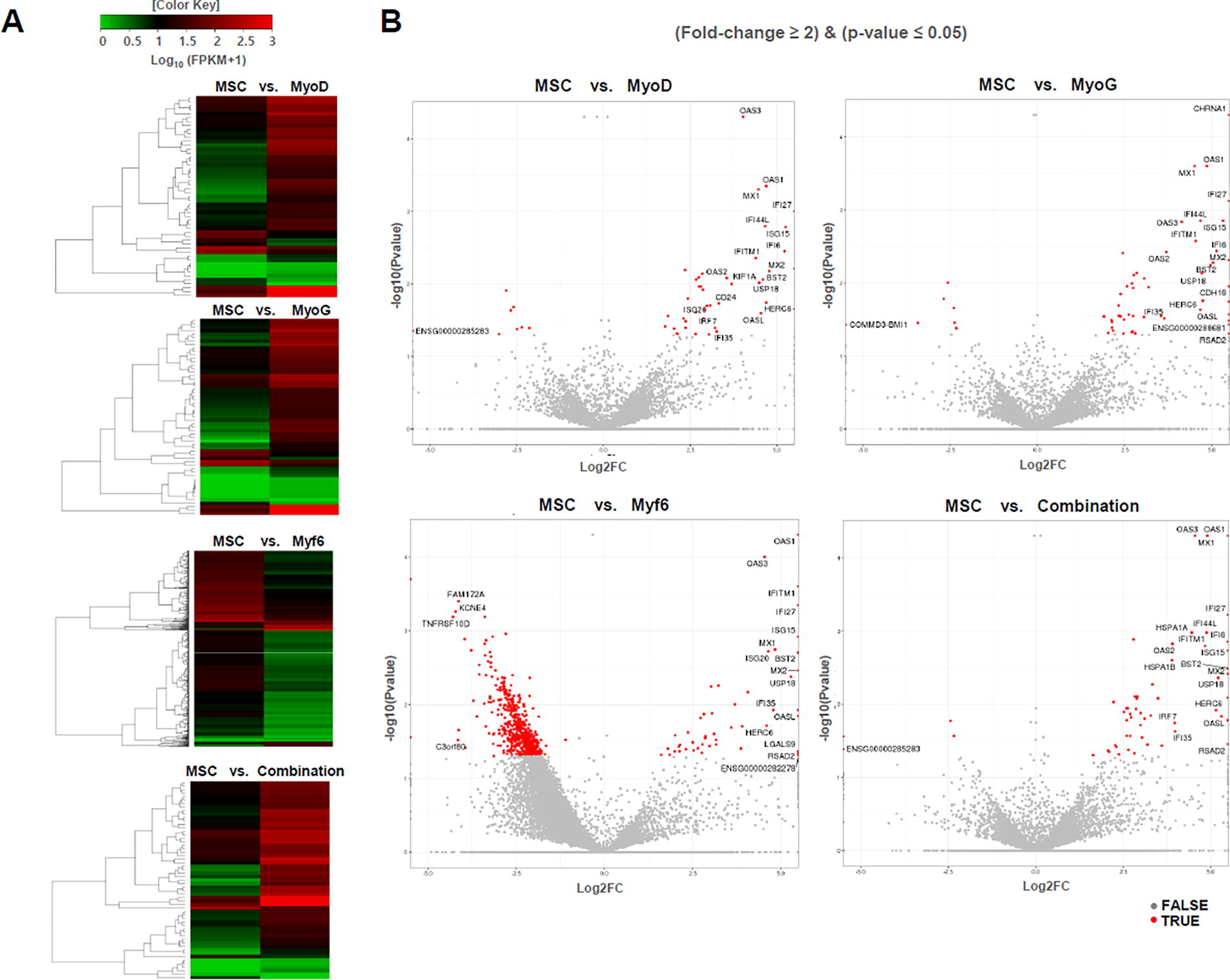
Heatmap and volcano plots of differential expression.
Discussion
Along with surgical therapy, RT in the treatment of head and neck cancer is recognized as an essential treatment method. Radiation exposure can cause subcutaneous fibrosis of soft tissues of the neck, which can cause neck pain and discomfort. 2,26 Dysphagia is a symptom that appears in ∼15% of patients with head and neck cancer. 27 The rate of dysphagia after RT varies greatly depending on the amount of radiation delivered to the muscle and the number of treatments. In soft tissue exposed to radiation, a reaction occurs with water molecules, forming hydroxyl radicals that can destroy intracellular DNA. 18 This is a key mediator of inflammation that triggers various immune cells to produce TGF-β. Activated TGF-β can reduce tissue and oxygenation and induce tissue ulceration and necrosis. 28 In the acute phase, radiation mucositis, dermatitis, and edema can occur due to tissue inflammatory reaction. 29 This inflammatory response might improve within three months. However, in many cases, the continuing cascade of cytokines in inflammation can cause chronic tissue damage and dysphagia. This can be explained by continuing inflammation due to long-term tissue hypoxia and oxidative stress following RT. Ultimately, genetic modification can be caused by destruction of tissue intracellular DNA. This can be a factor that hinders intrinsic functionality of the tissue.
This study aimed to develop a gene therapy system targeting mutated esophageal intramuscular genes. There are various tissue layers in the esophagus. Peristaltic movement of the esophagus is controlled by two muscle layers (longitudinal, circular). 30 The upper part of the esophagus is striated muscle. The lower esophagus consists of smooth muscles. 31 The middle part has the properties of both skeletal and smooth muscles. Importance of striated muscle in esophageal dysfunctions, including idiopathic myopathy, motility disorders, and so on, has been demonstrated. 32,33 Key genes for these esophageal muscle layers include MyoD, Myogenin, Myf6, MRF4, and so on. 20,34,35 Gene cloning using plasmid DNA, a nonviral vector, was performed for manipulation of altered genes due to radiation esophageal fibrosis. Meanwhile, studies that manipulate esophageal muscle genes and apply them to a radiation esophageal myofibrosis model have not been reported yet. As a gene fibrosis treatment model, studies on downregulation or cut-off of expression of a corrupted gene for treating liver fibrosis have been reported. 36,37 Researchers have claimed that genetic manipulation can transform myofibroblasts and turn them into healthy liver cells to support liver regeneration for treating liver fibrosis. Based on these research results, we hypothesized that by replacing major damaged genes in the esophageal muscle layer, we might be able to improve esophageal regeneration.
MSCs are well-known for their ability to enhance tissue regeneration and are among the most prominent cell therapies in the field of tissue engineering. 38 However, the limited number of cells available poses a significant hurdle to their development as effective cell therapies. In this study, we aimed to create high-functionality MSCs by transfected genes that enhance myogenesis, with the goal of maximizing functionality with a smaller number of cells. Combinational transfection of myogenic genes demonstrated superior effects compared with when MSCs alone, indicating the potential for high-functionality cell therapy. In addition to myogenesis, gene introduction related to chondrogenesis, osteogenesis, neurogenesis, adipogenesis, and various tissue regeneration is expected to enable the development of high-functionality MSCs. Furthermore, besides gene transfection, growth factor, hormones, and epigenetic regulators could also be used to develop high-functionality MSCs. The enhancement of esophageal reconstruction through the introduction of muscle-related genes into MSCs has the potential for various therapeutic applications. For this purpose, three types of muscle genes were transfected into human mesenchymal stem cells. High transfection efficiency and sustained expression of GFP were verified in vitro (Figs. 1B, 1C). MSCs have been reported to have the ability to self-renewal and differentiate into a desired cell lineage. 39 –41 Thus, they were used as a gene transfer cell model in the present study. In addition, cell regulatory factors secreted from MSCs have a significant paracrine effect on tissue regeneration. Sun and colleagues have reported that adipose-derived stem cell transplantation can alleviate irradiated fibrosis by suppressing TGF-β1 levels in radiation-damaged muscles. 42 Thus, ultimate treatment of fibrosis after irradiation through synergistic effect of gene recombination technology and tissue regeneration ability of adult stem cells is possible.
In the present study, the functional difference between MSC transfected with three type of genes and the control group (naïve hMSC) was verified in vitro. As a result, it was confirmed that the gene-transfected MSCs had high transgenic efficiency, maintained stemness, and had pluripotency. Therefore, we attempted to figure out the differential therapeutic efficacy between MSCs transfected with each gene through fibrosis-induced esophageal animal model. We have already verified the antifibrotic effects of hMSC through previous studies.
Transgenic MSCs were injected into the esophageal fibrosis model after irradiation. Various histological analyses were then performed at the one-month time point. Anatomical changes in three major esophageal tissue layers (EPI, IEL, and ME) were analyzed using H&E staining. Of these, the submucosal layer consists of dense and irregular connective tissue elastic collagen fibers. In addition, this layer includes veins, lymphatic vessels, and Meissner’s plexuses. It plays a major role in esophageal function. Basal cell hyperplasia due to submucosal fibrosis and countless vacuoles were observed. Fountain and his colleagues have described histological changes in detail in the esophageal submucosa that occur due to irradiation. 43 Many studies have reported the occurrence of edema due to hyperplasia of the collagen layer in the submucosal layer after irradiation. 4,44 We compared and analyzed fibrosis treatment efficacy of transgenic MSCs by measuring thickness of the submucosal layer. In addition, exposure to radiation within esophageal tissue led to changes in basal cell direction and polarity. In Figure 3D, hyperplasia of epithelial basal cells was observed, and a clear normalization of morphology was confirmed in the combination group. Therefore, the results of improved muscle formation by MyoD, MyoG, Myf6 gene-transfected MSCs were confirmed through histological analysis which indicated beneficial effects. RNA sequencing was performed to validate these results. Specifically, USP18, ISG15, IRF7, and OAS1 were notably upregulated in the combination group. SP18 has been reported as an essential regulator for the differentiation and maturation of muscle cells, and it is known to weaken immune responses by removing ISG15. However, it is also known to act independently in myogenesis. Our results also show its independent involvement and indicate that the expression of these two genes occurs together. These findings suggest that key regulator genes controlling muscle cell differentiation are enhanced through gene transfer. 45 Yamane reported that TGFA plays an important role in tongue muscle development in mice, and other researchers have also documented TGFA as a crucial gene in muscle differentiation. Our results further confirmed that TGF A is expressed in combination of three genes and influences muscle regeneration. 46,47 The transfer of three genes had a significant impact on enhancing myogenesis, which is crucial for muscle differentiation. It is therefore believed that the transplantation of esophageal tissues with these genetically modified MSCs would have effectively improved esophageal reconstruction.
However, the present study also has limitations. Although electroporation is much safer than viral vectors, plasmid DNA can also cause inflammation in transduced cells and leads to mutagenesis. To address these issues, using an appropriate concentration of plasmid DNA can be a solution. In addition, compared with viral vectors, the efficiency of gene delivery into cells may be lower. 48 In addition, efficacy verification of this study was performed in a relatively short period of one month. It is necessary to observe the efficacy of gene therapy on fibrosis from a long-term perspective. Generally, it takes a considerable amount of time to restore cell function by replacing genes lost due to radiation with normal genes. From a long-term perspective, verifying the efficiency and safety of gene delivery will be crucial information for the clinical application of gene therapy.
Conclusion
We generated MSCs transfected with three genes (MyoD, MyoG, and Myf6) as genetic regulator in esophageal muscle using a nonviral vector. Gene-transfected MSCs seem to provide a more suitable environment for gene restoration and muscle regeneration in esophagus undergoing radiation fibrosis. The signaling mechanism of MSC translocated genes within damaged muscle tissue remains unclear. However, various myogenesis-related genes were detected from RNAs of transgenic MSCs through gene profiling. Moreover, we found a positive signal of gene expression in muscle cells due to gene transfer. It was histologically proved to have a tissue regeneration effect. Furthermore, this technology is expected to be fully applicable as a treatment method for other conditions such as mucosal damage, lung function impairment, and bone marrow suppression that were caused by RT. It can be applied through the recombination of specific genes to the damaged organs and related diseases. This gene recombination technology, along with the antifibrosis effect of MSCs is expected to be broadly applicable as an alternative treatment for various radiation-induced fibrosis.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Korea Health Technology R&D Project through the
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3