
Abstract
Neural stem/progenitor cells (NSPCs) persist in the mammalian subventricular zone (SVZ) throughout life, responding to various pathophysiological stimuli and playing a crucial role in central nervous system repair. Although numerous studies have elucidated the role of stanniocalcin 2 (STC2) in regulating cell differentiation processes, its specific function in NSPCs differentiation remains poorly understood. Clarifying the role of STC2 in NSPCs is essential for devising novel strategies to enhance the intrinsic potential for brain regeneration postinjury. Our study revealed the expression of STC2 in NSPCs derived from the SVZ of the C57BL/6N mouse. In cultured SVZ-derived NSPCs, STC2 treatment significantly increased the number of Tuj1 and DCX-positive cells. Furthermore, STC2 injection into the lateral ventricle promoted the neuronal differentiation of NSPCs and migration to the olfactory bulb. Conversely, the STC2 knockdown produced the opposite effect. Further investigation showed that STC2 treatment enhanced AKT phosphorylation in cultured NSPCs, whereas STC2 inhibition hindered AKT activation. Notably, the neuronal differentiation induced by STC2 was blocked by the AKT inhibitor GSK690693, whereas the AKT activator SC79 reversed the impact of STC2 knockdown on neuronal differentiation. Our findings indicate that enhancing STC2 expression in SVZ-derived NSPCs facilitates neuronal differentiation, with AKT regulation potentially serving as a key intracellular target of STC2 signaling.
Introduction
Postnatal mammalian brains primarily contain neural stem/progenitor cells (NSPCs) in two specialized regions: the subgranular zone of the hippocampal dentate gyrus and the subventricular zone (SVZ) of the lateral ventricles (LVs). 1 In the postnatal central nervous system (CNS), the SVZ serves as the principal site for NSPCs. Neuroblasts originating from NSPCs in the SVZ migrate anteriorly and tangentially through the rostral migratory stream (RMS) to the olfactory bulb (OB). There, they disperse radially and differentiate into functional interneurons. 2 It is worth noting that NSPCs can be activated in response to various pathophysiological stimuli, such as stroke and traumatic injury. These activated cells then migrate to the site of damage and exert effects on CNS repair. 3 Therefore, understanding the regulation of NSPCs differentiation and migration is of paramount importance for therapeutic applications.
The adult mammalian NSPCs inhabit specialized extracellular microenvironments, known as niches, which play a crucial role in regulating NSPCs self-renewal, proliferation, and differentiation. 4 Drug targeting of niche modulation is preferred over genetic engineering, primarily because of its inherent advantage in therapeutic delivery, which exhibits substantial promise for the treatment of CNS disorders. As a member of the conserved secreted glycoprotein hormone family, stanniocalcin 2 (STC2) has been implicated in various biological processes such as cell proliferation, development, metabolism, and the regulation of calcium and phosphate homeostasis. 5 –7 Previous studies have shown elevated levels of STC2 in a wide range of tumor cells, where it contributes to their growth, progression, and invasive capabilities. 8 –10 Notably, recent investigations have further elucidated STC2’s involvement in stem cell biology, particularly in differentiation processes. For instance, during human mesenchymal stem cell adipogenic differentiation, a significant alteration in STC2 expression is observed. Enhanced expression of STC2 has been found to exert inhibitory effects on adipogenesis. 11 Silencing STC2 expression impairs osteoblastic cell differentiation and mineralization, whereas its overexpression enhances these processes. 12 These findings collectively indicate that STC2 may play a pivotal role in the differentiation and migration of stem cells. Despite these advancements, the specific function and mechanistic action of STC2 in NSPCs remain largely unexplored.
This study aimed to investigate the role of STC2 in the differentiation and migration of mouse SVZ NSPCs, as well as its underlying intracellular mechanism. Initially, we cultured primary SVZ NSPCs isolated from postnatal 7-day (P7) mice. Subsequently, we examined the impact of STC2 on neuronal differentiation and NSPCs migration both in vitro and in vivo. Lastly, we examined the involvement of intracellular AKT signaling. Our findings demonstrate the significant potential of STC2 as a therapeutic target for neural regeneration in CNS injuries, specifically by promoting neuron generation and migration from NSPCs.
Materials and Methods
Animals
The C57BL/6N mouse strain used in this study was procured from the Experimental Animal Center of Xi’an Jiaotong University. To minimize the impact of hormonal fluctuations on cell behavior, this study employed male mice as an experimental model. Adult male mice, weighing between 240 and 260 g, were employed for in vivo investigations, whereas P7 male mice served as the source for in vitro NSPCs isolation and cultivation. Mice were maintained in accordance with standard guidelines, including a 12-h light/dark cycle, unlimited access to food and water, and a temperature range of 20–24°C. All experimental procedures involving mice were ethically approved by the Biomedical Ethics Committee of the Health Science Center at Xi’an Jiaotong University, with a primary focus on minimizing animal suffering and minimizing the number of animals used.
Cannula implantation and stereotactic injection
Adult male mice were anesthetized with a 0.08 mg/g dose of pentobarbital sodium, ensuring a sterile surgical environment and maintaining their body temperature at 37°C using heating pads. The mice were placed in a stereotactic frame (RWD Life Science, 69100), and a midline incision was performed to expose the bregma. A gauge guide cannula (0.48 mm diameter, RWD Life Science) was precisely implanted into the right LV using the following coordinates: anterior–posterior, 0.10 mm anterior to the bregma; medial–lateral, 0.8 mm; and dorsal–ventral, 2.40 mm below the skull surface. To prevent infection and blockage, the guide cannula was sealed with a stylet extending beyond its tip until drug administration. Following cannulation, the mice were individually housed and allowed a minimum of 7 days for recovery before the infusion process. For STC2 administration, mice were anesthetized with 4% isoflurane and maintained at 2.5% isoflurane. A total of 5 μL of recombinant STC2 protein (10 ng/μL, R&D SYSTEMS, 9406-SO-050) was injected into the LV at a flow rate of 0.5 μL/min using a KD Scientific Legato™ 130 microsyringe pump. After the infusion, the injection cannula was retained in place for 5 min to prevent backflow and facilitate drug diffusion. Then, the internal injection cannula was slowly withdrawn, followed by the reinsertion of the stylet into the guide cannula.
For lentiviral injections, mice were positioned in a stereotactic frame, and a longitudinal incision was made along the midline of the skull to expose the bregma. A small hole was then drilled, and the lentivirus was delivered stereotaxically into the right LV using the following coordinates: anterior–posterior, 0.8 mm anterior to the bregma; medial–lateral, 0.9 mm; and dorsal–ventral, 3.6 mm below the skull surface. A total of 200 nL of the lentiviral solution was injected into the striatum at a flow rate of 20 nL/min using a microsyringe pump. The needle remained in place for 5 min before slow removal.
Primary culture of SVZ-derived NSPCs
P7 mice were anesthetized with 5% isoflurane and sacrificed by exposure to 65% CO2, and SVZ tissues were then isolated. The tissue samples were meticulously diced with scissors in ice-cold Dulbecco’s modified eagle medium/nutrient mixture F-12 (DMEM/F12) (Thermo Fisher Scientific, 11320033) and incubated in Trypsin-Ethylenediaminetetraacetic acid (EDTA) (Sigma-Aldrich, T4049) at 37°C for 3 min. Mechanical dissociation was performed using a P1000 pipette, followed by filtration through a 40-μm cell strainer (Corning Falcon, 352340) and centrifugation at 1,000 g for 3 min at 4°C. The resulting cells were seeded at a density of 30,000 cells/mL in nonadhesive T75 flasks containing complete medium. The complete medium consisted of serum-free DMEM/F12 supplemented with 1% N-2 (Cat. No. A13707-01), 2% B-27™ (Cat. No. 12587010), 20 ng/mL epidermal growth factor (EGF, Cat. No. PHG0311), and 10 ng/mL basic fibroblast growth factor (bFGF, Cat. No. PHG0369). All reagents were obtained from Gibco. Cultures were maintained at 37°C in a CO2 incubator (Memmert, ICO150) with Epidermal growth factor (EGF) and Basic fibroblast growth factor (bFGF) replenished every other day to promote neurosphere formation.
To induce differentiation, cells were transferred to a natural differentiation medium consisting of serum-free DMEM/F12 with 1% N-2 and 2% B-27 for 3 days. For dose-dependent treatment, NSPCs were exposed to various concentrations (1, 5, 10, 20, and 50 ng/mL) of recombinant STC2 protein for 3 days. To inhibit AKT activity, 10 μM GSK690693 (Selleckchem, S1113) was introduced 2 h before STC2 treatment. Conversely, to activate AKT, 10 μM SC79 (Selleckchem, S7863) was included in the medium of STC2 knockdown NSPCs, which were then cultured for 3 days.
STC2 knockdown treatment
The target-specific Small interfering RNA (siRNA) for STC2 was synthesized by Genechem, with the sequences as follows: si-STC2-1 (5′-UACAAGGUCCACGUAGGGUUCAUGC-3′), si-STC2-2 (5′-UUGUGCAGAAACGUCAUGCAAAUCC-3′), and siRNA negative contro(siNC) (5′-CGTACGCGGAATACTTCGATT-3′). Before transfection, NSPCs were cultivated on Poly-d-lysine (PDL)-coated plates for 2 days. The siRNA negative contro (siNC) delivery was accomplished using LipofectamineTM 3000 (Thermo Fisher Scientific, L3000001), and the transfection efficiency was evaluated using an inverted fluorescence microscope (DMI3000B, Leica). Knockdown efficacy was assessed through western blot analysis post-transfection. All experiments were conducted in triplicate and replicated independently at least three times to ensure reproducibility.
The lentivirus vector containing Small hairpin RNA (shRNA) targeting STC2 (snockdown-STC2 (KD-STC2), 7 × 107 TU/mL) and negative control vectors (shNC, 1 × 108 TU/mL) was purchased from GenePharma. This study used two lentiviral vectors: one expressing enhanced green fluorescent protein for transfection efficiency assessment and another without fluorescence for subsequent experiments. For in vivo experiments, the surgical procedure followed established protocols. A lentiviral solution (600 nL) was administered to the brain at a rate of 50 nL/min using a microsyringe pump, with the needle retained for 5 min before slow withdrawal. For in vitro experiments, mouse SVZ-derived NSPCs were transfected with KD-STC2 or shNC. Following passaging, the infected cells were selected with puromycin (1.6 μg/mL).
Immunostaining
Mice were anesthetized and subjected to transcardial perfusion with 50 mL of normal saline, followed by 200 mL of 4% paraformaldehyde (PFA). The brains were then dissected and fixed overnight at 4°C in 4% PFA, followed by cryoprotection in 30% sucrose for at least 3 days. Serial coronal sections, 16 μm thick, were obtained using a freezing microtome (Leica) along the SVZ-RMS-OB trajectory and mounted onto slides treated with an adhesive. SVZ-derived NSPCs were cultured, fixed with 4% PFA for 30 min at room temperature, and washed five times with 0.01 M Phosphatebuffered saline (PBS). Both slices and cell coverslips were permeabilized with 0.3% Triton X-100 (Sigma-Aldrich, X100-100ML) for 15 min. The samples were then washed and blocked with a solution containing 5% bovine serum albumin (Sigma-Aldrich, A8022) and 5% normal goat serum (Sigma-Aldrich, NS02L) for 2 h followed by incubation overnight at 4°C with primary antibodies (Supplementary Table S1). Negative controls were processed without primary antibodies. After washing, secondary antibodies (Supplementary Table S1) were applied for 2 h, followed by 4¢,6-diamidino-2-phenylindole(DAPI) or PI counterstaining. Images were captured using a fluorescence microscope (Olympus, BX51 + DP71).
In vivo analysis involved BrdU/DCX double labeling in three consecutive sets of sections from the SVZ (0.5–1.16 mm anterior to the bregma), the RMS (2.80–3.20 mm anterior), and the OB (3.82–4.28 mm anterior). Cells were counted using Image-Pro Plus 5.0 software (Olympus), using an unbiased stereological estimation method for accurate quantification. 13 For in vitro experiments, immunostained cells were counted in 10 randomly selected fields per sample, in triplicate, with the percentage of labeled cells normalized to DAPI nuclei staining. Image analysis was done using Image-Pro Plus 5.0 to automatically count nuclei with positive staining. Each assay was performed with three independently prepared NSPCs cultures.
BrdU labeling
BrdU (80 mg/kg) was administered intraperitoneally to each mouse in a regimen of four consecutive administrations, with each dose given at 6-h intervals throughout a single 24-h period. Following the final BrdU injection, recombinant STC2 protein was infused through LV injection, commencing 12 h later. To perform BrdU staining, samples underwent a pretreatment with 2N HCl at 37°C for 40 min, followed by neutralization in a 0.1 M borate buffer (pH 8.5) for 15 min.
Western blot analysis
At the end of experiments, cultured cells were harvested and lysed in RIPA buffer (Abcam, ab156034) supplemented with Protease Inhibitor Cocktail (Roche, 11697498001). The lysis process lasted 15 min on ice, followed by sonication using Sonics equipment, and subsequent centrifugation at 12,000 g for 10 min at 4°C. Protein concentration was determined using Bicinchoninic acid (BCA) Protein Assay Kits (Thermo Fisher Scientific, 23227). Each sample (20–40 μg) was subjected to 12% Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel electrophoresis and transfer to Polyvinylidene fluoride (PVDF) membranes (Bio-Rad, 1620177). Membranes were blocked with 5% nonfat milk for 2 h at room temperature and then incubated with primary antibodies (Supplementary Table S1) overnight at 4°C. Following three washes with Tris-buffered saline with Tween 20 (TBST), the membranes were exposed to Horseradish peroxidase (HRP)-conjugated secondary antibodies (Supplementary Table S1) for 2 h. The immunoreactive bands were visualized using SuperSignal™ West Pico PLUS Chemiluminescent Substrate (Thermo Fisher Scientific, 34580) and captured with a Fuji X-ray film. The bands were acquired using a gel imaging system (G: Box, Syngene) and analyzed with ImageJ software (NIH). β-Actin was used as a housekeeping protein for normalizing target protein expression levels. All western blot data displayed are from at least three independent experiments.
Statistical analysis
Statistical analyses were performed with GraphPad Prism 5.0 software. Normality and homoscedasticity were evaluated before comparing the data. One-way analysis of variance was used for significance testing, whereas Fisher’s Protected least significant difference (PLSD) test was deployed for comparing the two groups. The Kolmogorov–Smirnov test verified the data’s normality and homoscedasticity. Results were reported as mean ± standard deviation, with statistical significance defined as P < 0.05. Throughout the study, there were no instances of accidental mouse mortality, and all animal procedures exhibited normalcy. Subsequently, all mice were included in the result analysis, with no data points excluded. No outlier tests were conducted.
Results
STC2 is expressed in NSPCs in the mouse SVZ
Primary NSPCs were isolated from P7 mice. Immunostaining revealed that 95.1% ± 3.8% of cultured cells were SOX2-positive, with 96.1% ± 3.2% of these cells also expressing nestin (Fig. 1A). These cells were then cultured in a natural differentiation medium for 3 days, resulting in cells positive for Tuj1 (an immature neuron marker), GFAP (an astrocytic marker), and NG2 (an immature oligodendrocyte marker) (Fig. 1B–D). To investigate STC2 expression in mouse SVZ-derived NSPCs, adult brain tissue and cultured cells were analyzed. Immunostaining revealed a colocalization of STC2 with the NSPCs marker SOX2, both in vivo and in vitro (Fig. 1E, F). These results indicated the successful cultivation of SVZ-derived NSPCs and demonstrated that mouse NSPCs expressed STC2. This established culture system was subsequently used for further experiments.
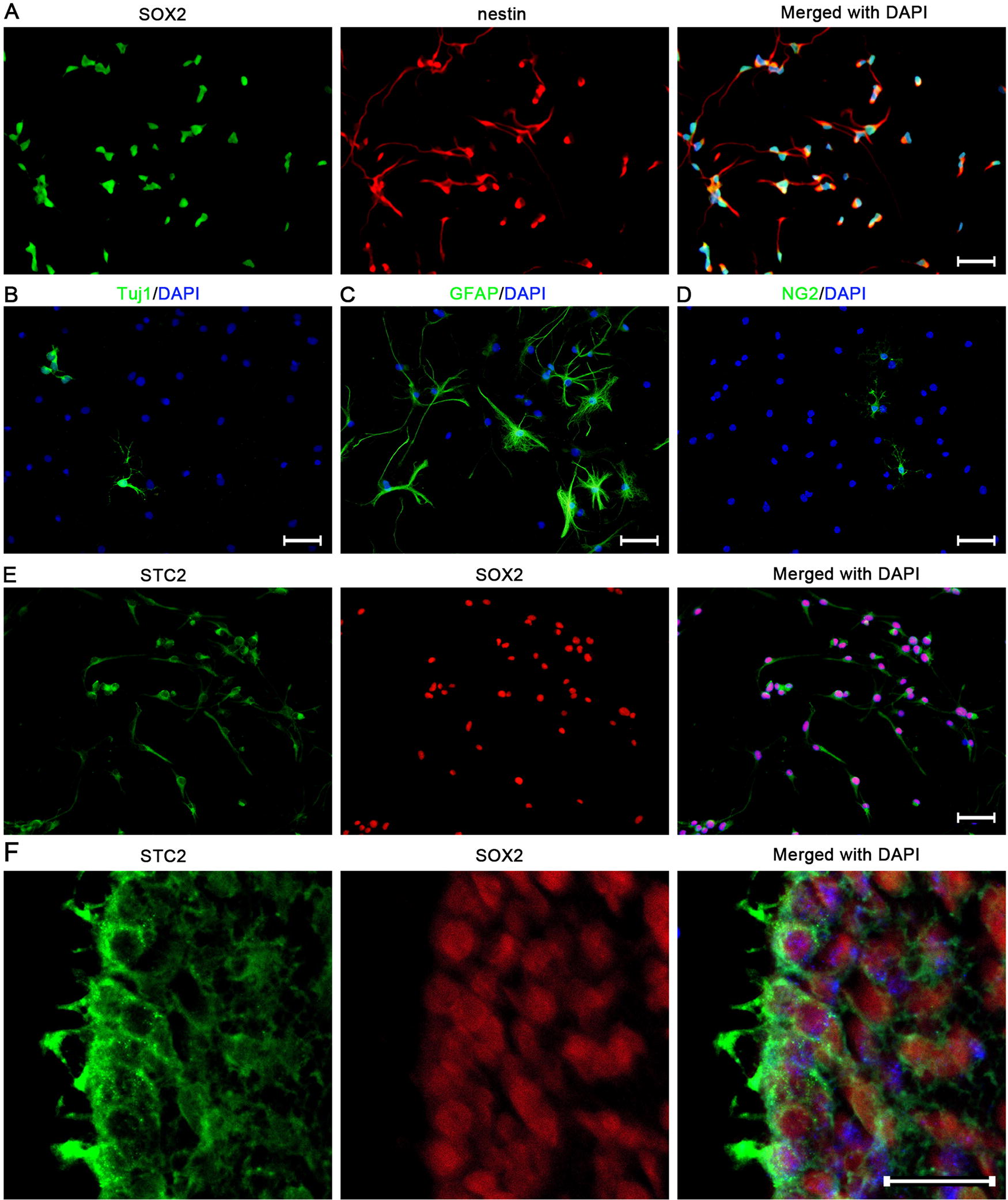
The expression of STC2 in SVZ-derived NSPCs.
STC2 promotes neuronal differentiation in cultured NSPCs
To rule out “off-target” effects, two STC2-specific siRNAs were employed to downregulate STC2 expression. These siRNAs were efficiently transfected into cultured NSPCs, resulting in a notable reduction in STC2 expression (Supplementary Fig. S1 A–C). Owing to its superior inhibitory efficacy, si-STC2-2 was chosen for lentiviral packaging. As anticipated, NSPCs were infected with high efficiency, leading to a substantial decrease in STC2 expression (Supplementary Fig. S1 D–F). This lentivirus was subsequently used for further experiments.
The NSPCs were cultured in a natural differentiation medium and exposed to various concentrations of STC2 for 3 days. Immunostaining revealed a significant dose-dependent increase in the number of Tuj1-positive cells (Fig. 2A). To exclude potential apoptotic effects, a Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay was performed, showing no significant difference between STC2 supplementation and STC2 expression inhibition (Fig. 2B, C). In addition, to further validate the effect of STC2 on NSPCs differentiation, immature neurons were identified through Tuj1 and DCX double staining. The results demonstrated that STC2 treatment led to a significant increase in both Tuj1 and DCX-positive cells, whereas STC2 knockdown exhibited the opposite effect (Fig. 2D–G). These data suggest the promotion of STC2 activation in the neuronal differentiation of NSPCs culture.
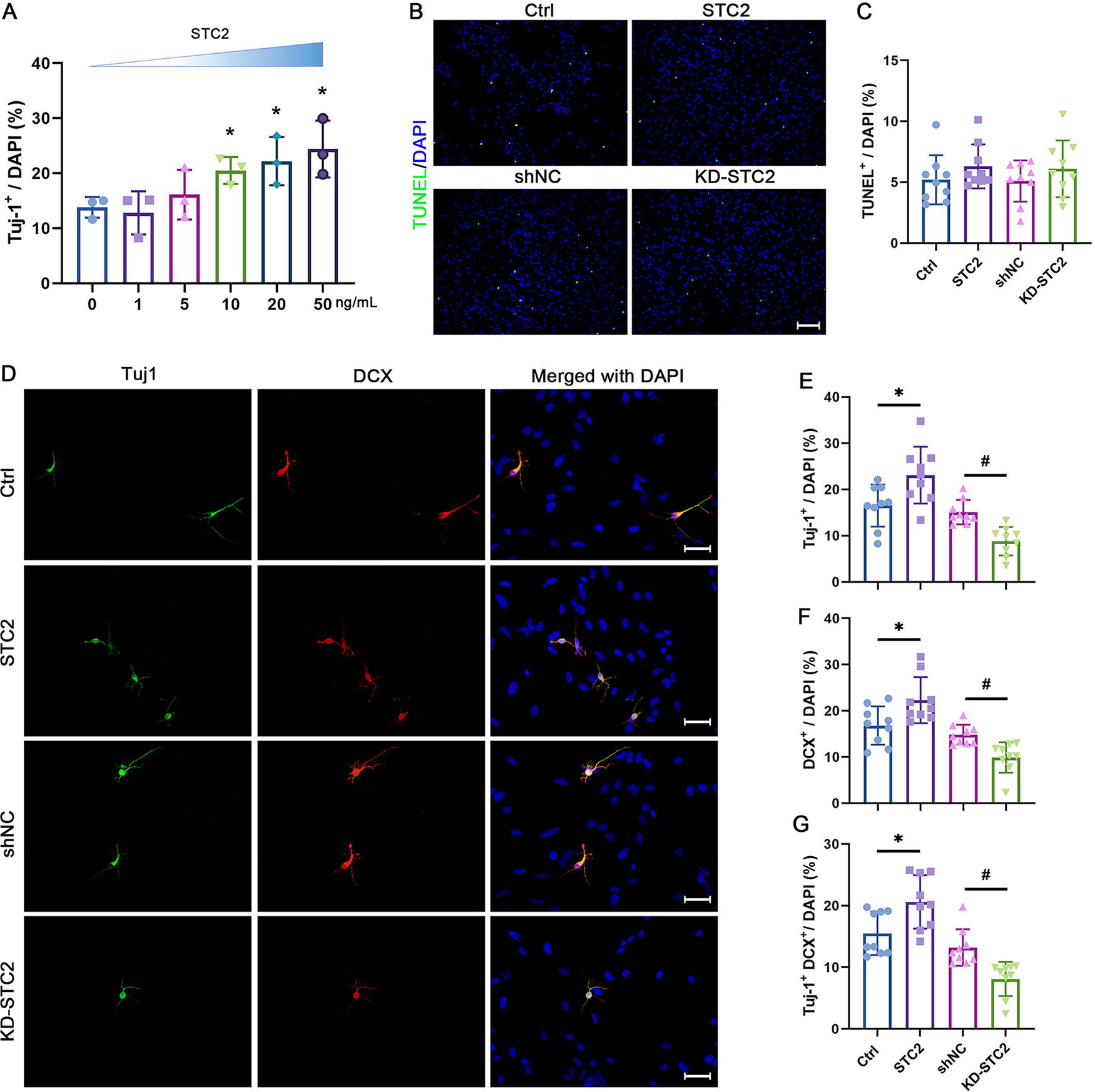
STC2 promotes neuronal differentiation in cultured NSPCs.
STC2 promotes neuronal differentiation of NSPCs in the SVZ
NSPCs give rise to neuroblasts which migrate long distance in the RMS to the OB, where they differentiate into neurons (Fig. 3A). To investigate cell differentiation, SVZ NSPCs were prelabeled with BrdU before the intervention. The number of double-labeled cells expressing DCX and BrdU was quantified in the SVZ, RMS, and OB at 14 days postinjection (Fig. 3B). The findings revealed a significant increase in BrdU/DCX colabeled cells in all regions following STC2 administration, whereas inhibition of STC2 led to a decrease (Fig. 3C, D; Fig. 4A–D). These data compellingly demonstrate that enhancing STC2 expression facilitates neuronal differentiation of NSPCs in adult mouse SVZ.

STC2 promotes neuronal differentiation in adult SVZ.
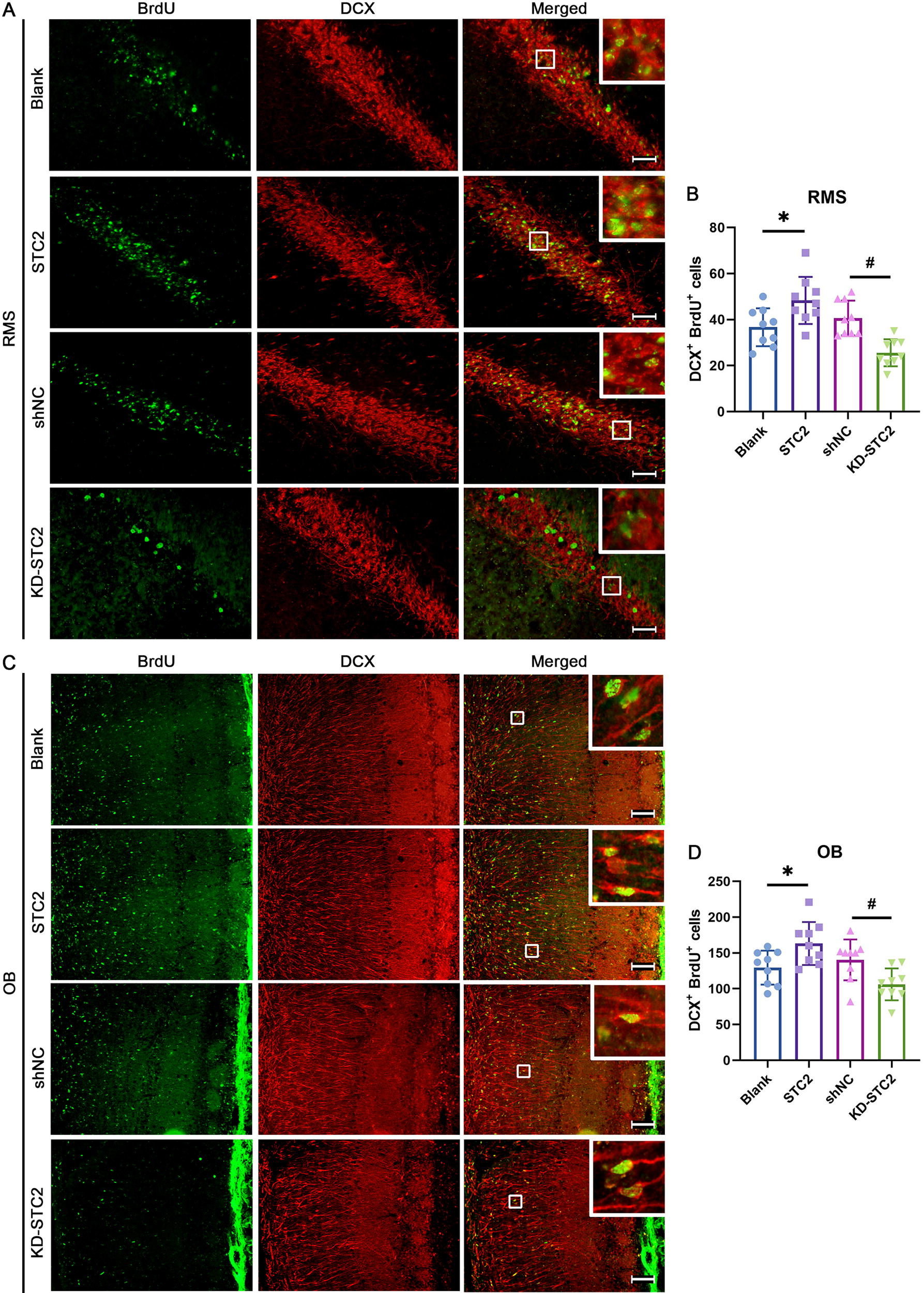
STC2 promotes neuronal differentiation in adult both RMS and OB. The mice underwent the previously described treatment, and BrdU and DCX-positive cells were examined in both the RMS
STC2 promotes neuronal differentiation of NSPCs involving AKT activation
The AKT signaling pathway plays a central role in neuronal differentiation, as highlighted by several studies. 14 –16 Given the established connection between STC2 and the regulation of AKT activation, 17 –19 our investigation centered on this pathway. To uncover the molecular mechanisms underlying the STC2-induced promotion of neuronal differentiation, we examined the effect of STC2 on AKT pathway activation. Western blot analysis demonstrated a significant dose-dependent elevation in the phosphorylation of AKT upon STC2 treatment (Fig. 5A, B). Remarkably, downregulating STC2 significantly inhibited AKT activation (Fig. 5C, D). These results unambiguously establish that STC2 is a critical regulator of AKT pathway activity.

The influence of STC2 on neuronal differentiation in NSPCs derived from the SVZ is associated with the modulation of AKT activation. NSPCs were exposed to varying concentrations of STC2 (0–50 ng/mL). Representative bands
To investigate the role of AKT in STC2-mediated regulation of SVZ-derived NSPCs’ neuronal differentiation, we pretreated NSPCs with the AKT inhibitor GSK690693 (10 μM) and the AKT activator SC79 (10 μM), respectively. Immunostaining revealed that GSK690693 attenuated the effect of STC2 on the number of Tuj1 and DCX-positive cells. Consistent with the GSK690693 treatment, SC79 abolished the neuronal differentiation effect observed upon STC2 knockdown (Fig. 5E–H). These results suggested that the promotion of neuronal differentiation by STC2 may involve modulation of the AKT signaling pathway (Fig. 6).
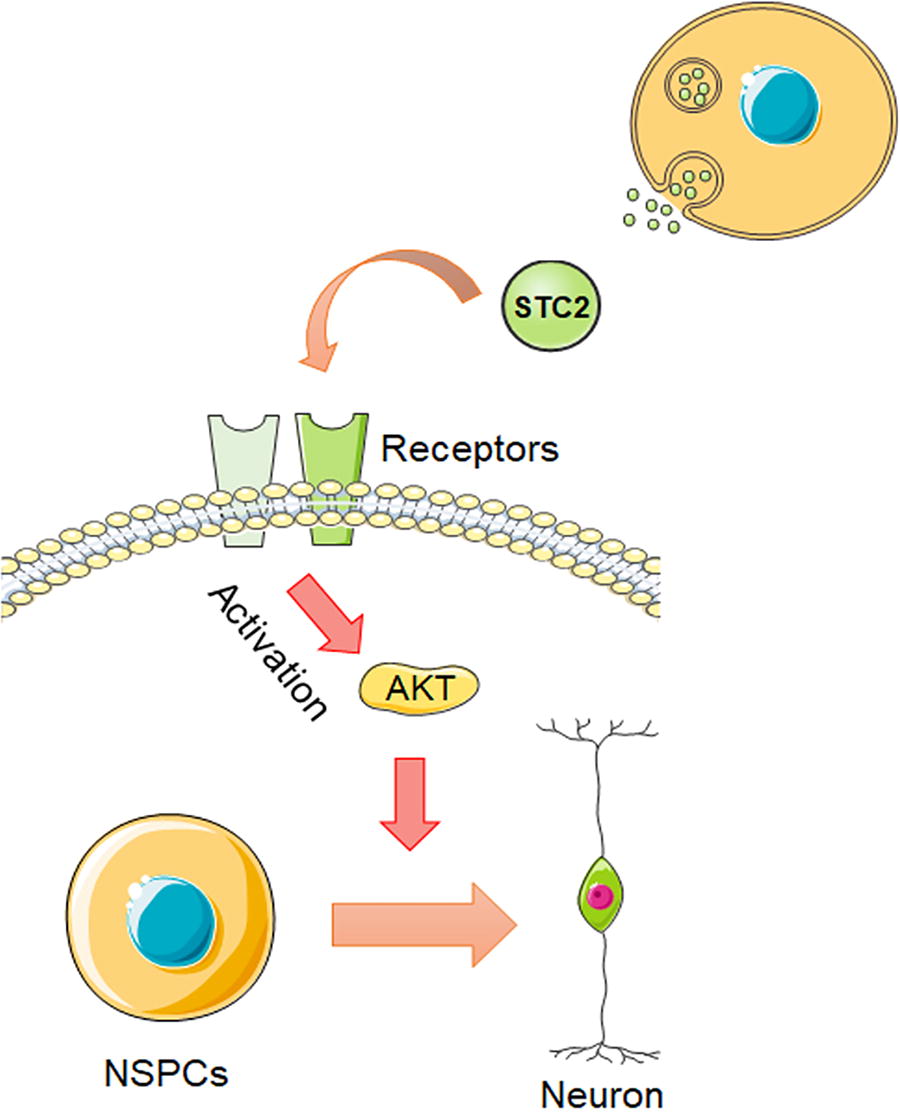
The illustration delineates the mechanism through which STC2 governs the neuronal differentiation of NSPCs. The secretion of the STC2 protein facilitates the activation of AKT by binding to its receptors, thereby promoting the neuronal differentiation of SVZ-derived NSPCs.
Discussion
CNS diseases, such as neurodegenerative diseases, cerebrovascular illnesses, and traumatic brain injuries, pose significant health challenges. However, conventional therapies exhibit limited efficacy against these conditions. NSPCs persist in the mammalian brain throughout life. Numerous studies have suggested that enhancing neuronal differentiation and migration of NSPCs could contribute to CNS repair and disease treatment. 20 –22 Our present study demonstrates that STC2 was expressed in the SVZ-derived NSPCs. Overexpression of STC2 promotes NSPC differentiation into neurons and facilitates their migration into the OB, whereas its inhibition yields opposite effects. The regulation of the AKT signaling pathway may represent a potential intracellular target of STC2 in NSPCs. These findings highlight the promising potential of STC2 as a therapeutic target for neural regeneration in the treatment of CNS injuries and enhancing neuron production from NSPCs. Nonetheless, it is crucial to acknowledge the substantial disparities between adult human NSPCs and their rodent counterparts, as their functions cannot be equated. 23 –25 For instance, human NSPCs exhibit higher diversity and enhanced proliferative potential compared with rodent NSPCs. 26 To fully comprehend the role of STC in regulating human NSPCs differentiation, further investigations are required.
Mammals express two isoforms of stanniocalcins, STC1 and STC2, involved in various biological processes including calcium homeostasis, cell differentiation and proliferation, oxidative stress, metabolism, and cancer development. 27,28 Stanniocalcins primarily exert their regulatory effects through paracrine and autocrine mechanisms. 29 Although an identifiable receptor for STC2 remains elusive, there is a proposition suggesting the involvement of the Pregnancy-associated plasma protein Ainsulin-like growth factorbinding protein (PAPP-A-IGFBP) IGF axis. 30 Numerous studies have consistently shown that insulin-like growth factor (IGF) significantly enhances AKT activation. 31,32 In addition, activation of the IGF receptor has been shown to facilitate cell migration in the SVZ-OB region, as well as promote neuronal survival and axonal growth in both the SVZ-OB and hippocampus. 33,34 Hence, it is plausible to hypothesize that STC2 regulates neuronal differentiation of NSPCs via the IGF receptor. Further research is necessary to confirm this hypothesis and explore the role of other potential receptors involved in the process.
Our study demonstrates that STC2 treatment significantly enhances AKT phosphorylation, whereas knockdown of STC2 exerts the opposite effect. More importantly, manipulation of AKT activation can abolish the effect of SCT2 on AKT activation, suggesting a plausible link between STC2 and neuronal differentiation via AKT activation. AKT, also known as protein kinase B, plays a pivotal role in the PI3K-AKT pathway, which is crucial for NSPCs differentiation. 35 –37 Valproic acid-induced neuronal differentiation in NSPCs also relies on AKT activation. 38 However, inhibition of PI3K/AKT with LY294002 reduces proliferation and inhibits dopaminergic neuronal differentiation in human NSPCs. 39 Conversely, AKT inhibition by activating mGluR4 promotes proliferation and inhibits neural differentiation in NSPCs. 40 Overexpression of AKT diminishes GFAP and Tuj1 expression during differentiation, suggesting an inhibitory role in cell differentiation. 41 This paradoxical behavior of AKT signaling may result from its interactions with other signaling pathways rather than functioning in isolation. Protein kinase cascades typically operate within a network of cross-talk between different signaling pathways, where modulation of one kinase cascade can influence the activity of others. 42 –44 These interconnected pathways form a complex biochemical network that modulates cellular behaviors such as neuronal differentiation and migration. Future investigations are needed to fully elucidate the cross-talk between AKT and other pathways in the context of STC2-induced neuronal differentiation. In addition, further research is needed to determine whether other signaling pathways mediate the aforementioned differentiation mechanism.
Footnotes
Acknowledgment
The authors thank the Experimental Animal Center of Xi’an Jiaotong University for mouse husbandry.
Ethics Statement
All animal experiments were approved by The Biomedical Ethics Committee of Health Science Center of Xi’an Jiaotong University (No. XJTUAE2023-403). This study conforms to the Declaration of Helsinki.
Data Availability Statement
The data used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This work was supported by Key Research and Development Projects of Shaanxi Province (2019SF-028).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1