
Abstract
The periodontal ligament (PDL) is a fibrous connective tissue that connects the cementum of the root to the alveolar bone. PDL stem cells (PDLSCs) contained in the PDL can differentiate into cementoblasts, osteoblasts, and PDL fibroblasts, with essential roles in periodontal tissue regeneration. Therefore, PDLSCs are expected to be useful in periodontal tissue regeneration therapy. In a previous study, we differentiated induced pluripotent stem cells (iPSCs) into PDLSC-like cells (iPDLSCs), which expressed PDL-related markers and mesenchymal stem cell (MSC) markers; they also exhibited high proliferation and multipotency. However, the iPSCs used in this differentiation method were cultured on mouse embryonic fibroblasts; thus, they constituted on-feeder iPSCs (OF-iPSCs). Considering the risk of contamination with feeder cell-derived components, iPDLSCs differentiated from OF-iPSCs (ie, OF-iPDLSCs) are unsuitable for clinical applications. In this study, we aimed to obtain PDLSC-like cells from feeder-free iPSCs (FF-iPSCs) using OF-iPDLSC differentiation method. First, we differentiated FF-iPSCs into neural crest cell-like cells (FF-iNCCs) and confirmed that FF-iNCCs expressed NCC markers (eg, Nestin and p75NTR). Then, we cultured FF-iNCCs on human primary PDL cell-derived extracellular matrix for 2 weeks; the resulting cells were named FF-iPDLSCs. FF-iPDLSCs exhibited higher expression of PDL-related and MSC markers compared with OF-iPDLSCs. FF-iPDLSCs also demonstrated proliferation and multipotency in vitro. Finally, we analyzed the ability of FF-iPDLSCs to form periodontal tissue in vivo upon subcutaneous transplantation with β-tricalcium phosphate scaffolds into dorsal tissues of immunodeficient mice. Eight weeks after transplantation, FF-iPDLSCs had formed osteocalcin-positive bone/cementum-like tissues and collagen 1-positive PDL-like fibers. These results suggested that we successfully obtained PDLSC-like cells from FF-iPSCs. Our findings will contribute to the development of novel periodontal regeneration therapies.
Introduction
Periodontal tissue, consisting of gingival, cementum, alveolar bone, and periodontal ligament (PDL), plays an important role in maintaining tooth health. 1 In particular, PDL stem cells (PDLSCs; ie, stem cells in the PDL) contribute to periodontal tissue regeneration. 2 Seo et al. demonstrated that PDLSCs can differentiate into osteoblasts and adipocytes in vitro. 3 Moreover, PDLSCs express both PDL and mesenchymal stem cell (MSC) markers. 2,3 Previous studies have revealed that when PDLSCs are subcutaneously implanted in mice, they form cementum-like and PDL-like tissues. 3,4 Furthermore, autologous transplantation of PDLSCs reportedly can regenerate injured periodontal tissue. 5,6 However, because acquisition of PDLSCs requires tooth extraction, it is difficult to obtain the large numbers of PDLSCs required for clinical applications. Thus, there is a need for a method that enables a large number of PDLSCs to be acquired without tooth extraction.
Yamanaka et al. created induced pluripotent stem cells (iPSCs) by transfecting skin fibroblasts with four genes: OCT3/4, SOX2, KLF4, and c-Myc. These cells expressed embryonic stem cell markers, such as NANOG and E-Ras. 7 iPSCs possess the ability to differentiate into various cell types, including neurons, myocardial cells, and endothelial cells; they are expected to play key roles in regenerative medicine. 8 –10 Initially, mouse embryonic fibroblasts (MEFs) were required as feeder cells for cultures of iPSCs. 7 However, on-feeder iPSCs (OF-iPSCs) are unsuitable for clinical applications due to the risk of contamination with feeder cell-derived components. In addition, the OF-iPSC culturing method is complex because it involves MEF preparation and specialized passaging techniques. Therefore, a feeder-free iPSC (FF-iPSC) culture method has been developed. 11 This FF method is suitable for clinical applications because it eliminates the risk of contamination with feeder cell-derived components and simplifies the subculture of iPSCs.
In our laboratory, we have successfully differentiated OF-iPSCs into PDLSC-like cells (OF-iPDLSCs). 12 We demonstrated that OF-iPDLSCs exhibit high proliferation and multipotency; they also express MSC and PDL-related markers. However, because the differentiation process to obtain OF-iPDLSCs involves the use of MEFs and human primary PDL cell (HPDLC)-derived extracellular matrix (ECM), clinical applications for these cells and matrix are challenging. In this study, we investigated whether PDLSC-like cells could be obtained from FF-iPSCs using the OF-iPDLSC differentiation method.
Materials and Methods
Cell culture
iPSCs were obtained from RIKEN (Ibaraki, Japan; HPS 0063). This iPSc line was established from skin fibroblasts from a 36-year-old healthy woman. OF-iPSCs were cultured on MEFs (ReproCELL, Kanagawa, Japan) and maintained in primate embryonic stem cell medium (serum-free; ReproCELL) containing 5 ng/mL human recombinant basic fibroblast growth factor (b-FGF; ReproCELL). FF-iPSCs were cultured on dishes coated with iMatrix-511 (Nippi, Tokyo, Japan) using StemFit AK03N Medium (serum-free; Ajinomoto, Tokyo, Japan). HPDLCs were isolated from the third molar of a healthy 25-year-old female patient at Kyushu University. The HPDLCs were cultured in alpha-minimum essential medium (α-MEM; Gibco-BRL, Grand Island, NY, USA) supplemented with 50U/µL penicillin and 50 µg/mL streptomycin (Wako, Osaka, Japan), as well as 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO, USA) (10% FBS/α-MEM). For this study, HPDLCs from passages 6 through 8 were used. All procedures adhered to the principles outlined in the Declaration of Helsinki and were conducted in accordance with the guidelines of the Research Ethics Committee of Kyushu University Certified Institutional Review Board for Clinical Trials (Approval No. 2–115, issued on August 10, 2020).
Differentiation of OF/FF-iPSCs into neural crest-like cells
OF/FF-iPSCs were differentiated into OF/FF-neural crest-like cells (OF/FF-iNCCs) using an established method. 13,14 Briefly, OF-iPSCs were dissociated into single cells with Accutase (Nacalai Tesque, Kyoto, Japan) and cultured on Matrigel-coated dishes (1 × 105 cells/3.5 cm dish) using MEF-conditioned medium supplemented with 10 mM Y-27632 (Rho-associated coiled-coil containing kinase/Rho-binding kinase inhibitor; Enzo Life Sciences, Farmingdale, NY, USA) and 10 ng/mL b-FGF. After the cultures reached 50%–70% confluence, cells were cultured in KnockOut Serum Replacement medium (serum-free) and neurobasal medium (serum-free) supplemented with Noggin (PeproTech, Inc., Rocky Hill, NJ, USA) and SB431542 (TGF-β inhibitor; Tocris, Minneapolis, MN, USA). 13 After 12–13 days, cells expressing p75NTR, a neural crest marker, were isolated by MACS (Miltenyi Biotec, Bergisch Gladbach, Germany). The isolated cells were named OF-iNCCs.
FF-iPSCs (1.3 × 104 cells/well) were seeded in a six-well plate (Corning Incorporated, Corning, NY, USA) coated with iMatrix-511 and cultured in StemFit AK03N Medium containing 10 mM Y-27632. The following day and 3 days later, the medium was changed to StemFit AK03N Medium without Y-27632. After 4 days, cells were cultured in AK03N Medium supplemented with 10 μM SB431542 (Fujifilm Wako) and 1 µM CHIR99021 (GSK-3 inhibitor; Axon Medchem, Reston, VA, USA) for 10 days. The medium was changed every 3 days (days 0–6) and then every 2 days (days 7–10). 14 The resulting cells were named FF-iNCCs.
Differentiation of OF/FF-iNCCs into OF/FF-iPDLSCs
HPDLCs were seeded in 24-well plates in 10% FBS/α-MEM (1.0 × 104 cells/well). After 4 days, detachment was achieved using 2% ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque) in phosphate-buffered saline (PBS) to obtain dishes coated with ECM. OF-iNCCs (1.0 × 105 cells/well) and FF-iNCCs (1.0 × 104 cells/well) were seeded on these HPDLC-derived ECM-coated dishes; the cells were cultured for 2 weeks and named OF-iPDLSCs and FF-iPDLSCs, respectively.
Quantitative reverse transcription polymerase chain reaction
Total cellar RNA was isolated with TRIzol Reagent (Invitrogen, Carlsbad, CA, USA). Quantitative reverse transcription polymerase chain reaction (RT-PCR) was performed using a KAPA SYBR Fast qPCR kit (Nippon Genetics, Tokyo, Japan) in a Thermal Cycler Dice Real Time System (Takara Bio Inc., Shiga, Japan). Primer sequences, product sizes, and annealing temperatures for each gene are listed in Table 1. β-actin was used as an internal control. The expression levels of the target genes were determined by calculation of ΔΔCt values.
Primer Sequences, Product Sizes, and Annealing Temperatures for Quantitative Reverse Transcription Polymerase Chain Reaction
BMP2, bone morphogenetic protein 2; BSP, bone sialoprotein; COL1, collagen I; FBN2, fibrillin-2; LPL, lipoprotein lipase; MKX, Mohawk; OPG, osteoprotegerin; PLAP1, periodontal ligament-associated protein 1; PPARγ, peroxisome proliferator-activated receptor γ; SOST, sclerostin.
Immunofluorescence staining
OF/FF-iPSCs, iNCCs, and iPDLSCs were fixed with 4% paraformaldehyde (Nacalai Tesque) and 0.5% dimethyl sulfoxide (Nacalai Tesque) in PBS. Subsequently, cells were permeabilized with 0.3% Triton X-100 (Nacalai Tesque) in PBS, then blocked with 2% bovine serum albumin (BSA; Nacalai Tesque) in PBS. Cells were incubated overnight at 4°C with antihuman NANOG antibody (Abcam, Inc., Cambridge, MA, USA) and antihuman Nestin antibody (Novus Biologicals, Centennial, CO, USA). OF/FF-iPDLSCs were also incubated with antihuman osteoprotegerin (OPG) antibody (Abcam) and antihuman COL1 antibody (Abcam). Normal rabbit IgG (Cell Signaling Technology, Danvers, MA, USA) or normal chicken IgY control (R&D Systems, Minneapolis, MN, USA) was used as a control for each antibody. Next, cells were incubated with an Alexa Fluor 488-conjugated chicken antirabbit antibody, Alexa Fluor 647-conjugated goat antirabbit antibody, or Alexa Fluor 488-conjugated goat antichicken antibody (Invitrogen). The nuclei were stained with 4′,6-diamidino-2-phenylindole (Nacalai Tesque). Stained cells were imaged and analyzed using a Keyence BZ-X700 microscope (Keyence Corporation, Osaka, Japan).
Flow cytometry analysis
OF/FF-iPDLSCs were detached using Accutase, adjusted to a density of 5 × 105 cells/tube, and then incubated with R-phycoerythrin-conjugated mouse antihuman antibodies targeting CD34, CD45, CD90, CD105, CD146 (all from BioLegend, San Diego, CA, USA), IgG1, or IgG2a (both from iCyte, Tokyo, Japan) for 45 min at 4°C. The percentages of positive cells were assessed by flow cytometry (EC800 Cell Analyzer; Sony, Tokyo, Japan) and Eclipse software (Sony).
Proliferation assay
OF/FF-iPDLSCs were seeded in a 48-well plate (5 × 103 cells/well). Subsequently, on days 0, 1, 2, 3, proliferation was assessed using a water-soluble tetrazolium (WST)-1 cell proliferation assay kit (Takara Bio, Inc.) according to the manufacturer’s protocol. The supernatant was collected from each well and subjected to measurement of optical density at 450 nm using an iMark microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Osteoblast differentiation
OF/FF-iPDLSCs (2 × 104 cells/well) were seeded in a 24-well plate and cultured in 10% FBS/α-MEM. After cells had reached confluence, the culture medium was changed to osteoblast differentiation medium, consisting of 10% FBS/α-MEM supplemented with 50 µg/mL ascorbic acid (Nacalai Tesque) and 2 µM β-glycerophosphate (Sigma-Aldrich). After 4 weeks of culture, Alizarin Red S staining was performed. Images of stained cells were captured with a Keyence BZ-9000 microscope (Keyence Corporation); areas of positive staining were measured using BZ-X Analyzer Software (Keyence Corporation).
Adipocyte differentiation
OF/FF-iPDLSCs (2 × 104 cells/well) were seeded in a 24-well plate and cultured in 10% FBS/α-MEM. After cells had reached confluence, the culture medium was changed to adipocyte differentiation medium, consisting of 10% FBS/α-MEM supplemented with 1% L-glutamine (Nacalai Tesque), 0.1 mM L-ascorbic acid (Fujifilm Wako), 1 mM sodium pyruvate solution (Nacalai Tesque), 10 µM hydroxyethyl-piperazinyl ethanesulfonic acid (Nacalai Tesque), 60 µM indomethacin (Sigma-Aldrich), and 10−7 M dexamethasone (Merck Millipore, Darmstadt, Germany). After 3 weeks of culture, Oil Red O staining was performed. Fatty lipids were imaged with a Keyence BZ-9000 microscope and the number of Oil Red O-positive cells was determined using BZ-X Analyzer Software. Nine fields were randomly selected for quantification of positive cells.
Subcutaneous implantation of FF-iPDLSCs in immunodeficient mice
FF-iPDLSCs (3 × 106 cells) were mixed with 30 mg β-tricalcium phosphate (β-TCP) powder (OSferion; Olympus, Tokyo, Japan) and incubated at 37°C overnight. Next, the mixture was centrifuged and the supernatant was aspirated. The pellets were mixed with fibrinogen (Sigma-Aldrich) and thrombin (Sigma-Aldrich). Then, we made mid-longitudinal skin incisions on the dorsal surface of 6-week-old male immunodeficient mice (NOD SCID; Nihon Crea, Tokyo, Japan) and separated the skin bluntly from the muscle to form a pocket. Finally, the construct was placed into each pocket. As a control, cell-free β-TCP was transplanted.
After 8 weeks, the mice were sacrificed; harvested tissues were fixed with 4% paraformaldehyde and decalcified with 10% EDTA. Subsequently, the tissues were embedded in paraffin and sectioned at a thickness of 5 µm.
Histological analysis of FF-iPDLSCs
Sections were subjected to Masson’s trichrome staining or immunohistochemical staining. Briefly, sections were blocked with 2% BSA in PBS, then incubated at 4°C overnight with antihuman COL1 (1:250 dilution, Abcam), COL3 (1:1000 dilution, Cell Signaling Technology), OPG (1:4000 dilution, Abcam), OCN (1:2000 dilution, Invitrogen), Osterix (OSX; 1:500 dilution, Abcam), NuMA antibody (1:500 dilution, Abcam), and normal rabbit IgG (Cell Signaling Technology). Next, sections were incubated with biotinylated antirabbit IgG (Nichirei Biosciences, Tokyo, Japan), followed by an avidin–peroxidase conjugate (Nichirei Biosciences). Immunopositive reactions were identified using a simple diaminobenzidine staining solution (Nichirei Biosciences) and nuclei were counterstained with Mayer’s hematoxylin solution (Fujifilm Wako).
Statistical analysis
Statistical analysis was performed using unpaired two-tailed t-tests (for comparisons of two groups), one-way ANOVA followed by the Bonferroni method, or Kruskal–Wallis test followed by the Steel–Dwass method, (for comparisons of ≥3 groups). P values < 0.05 were considered statistically significant by EZR (Saitama Medical Center, Jichi Medical University, Saitama, Japan).
Results
Expression of iPSC and NCC markers in OF/FF-iPSCs, OF/FF-iNCCs, and OF/FF-iPDLSCs
We investigated whether FF-iNCCs and FF-iPDLSCs could acquire their specific morphologies under the culture conditions used in this study. FF-iNCCs exhibited a stellate-shaped morphology, whereas FF-iPDLSCs had a spindle-shaped morphology (Fig. 1A). FF-iNCCs and FF-iPDLSCs had morphologies similar to OF-iNCCs and OF-iPDLSCs, respectively. Next, to confirm the differentiation stage of iPSCs, we analyzed gene and protein expression patterns of iPSC markers (NANOG and OCT3/4) and NCC markers (Nestin and p75NTR). The expression levels of iPSC markers were significantly lower in OF/FF-iNCCs and OF/FF-iPDLSCs than in OF/FF-iPSCs (Fig. 1B). The expression levels of NCC markers were significantly higher in OF/FF-iNCCs than in other cell groups. Additionally, we analyzed protein expression levels of NANOG and Nestin in each cell type by immunofluorescence (Fig. 1C and D). NANOG expression was higher in OF/FF-iPSCs, while Nestin expression was higher in OF/FF-iNCCs compared to other cell types. These results suggested that the differentiation protocol used in this study could promote the differentiation of FF-iPSCs into FF-iNCCs, similar to the protocol for OF-iPSCs.
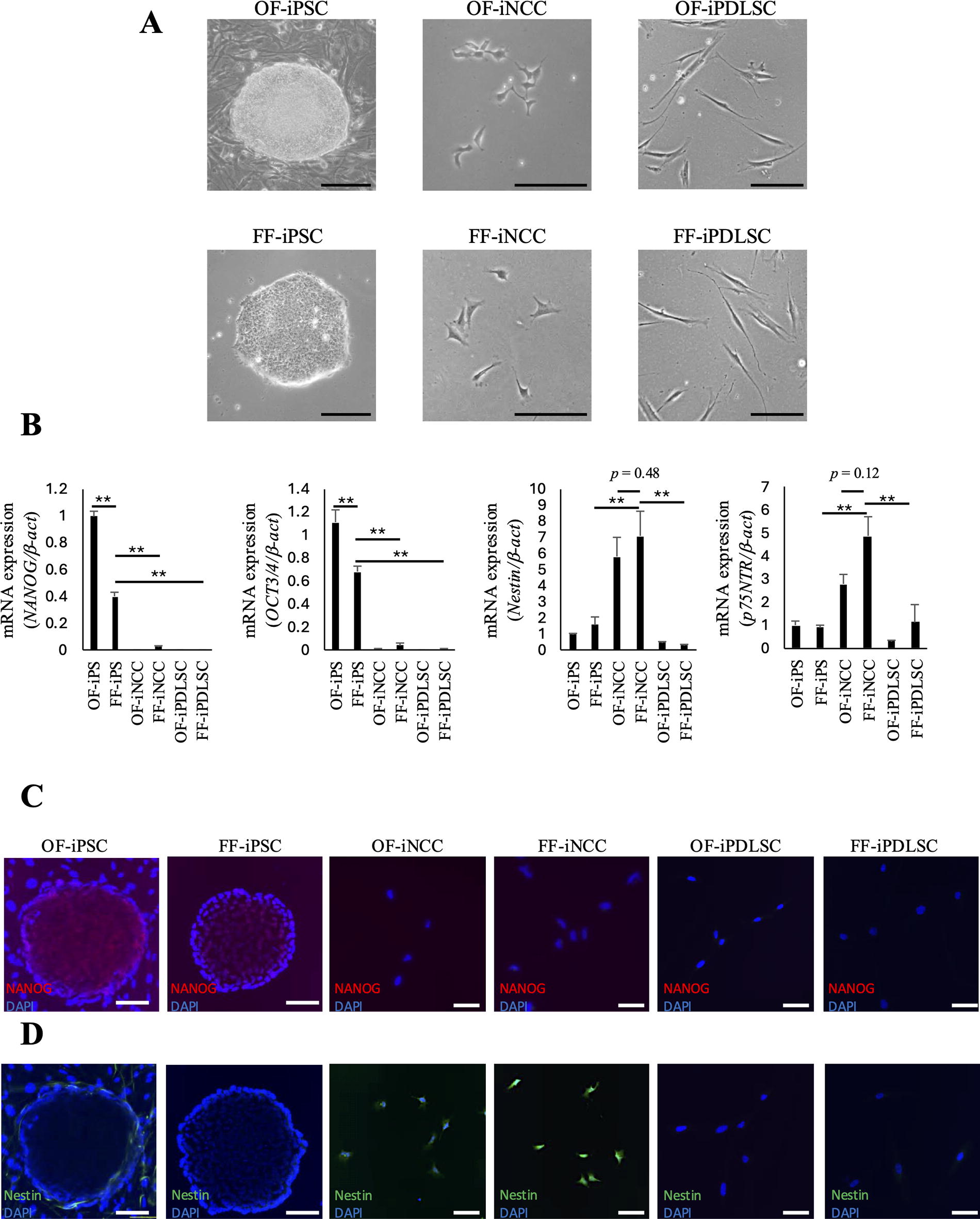
Expression of iPSC and NCC markers in OF/FF-iPSCs, OF/FF-iNCCs, and OF/FF-iPDLSCs.
MSC marker expression and proliferation ability in OF/FF-iPDLSCs
We analyzed the expression patterns of MSC markers in OF/FF-iPDLSCs by flow cytometry. Both cell types showed high expression levels of the MSC markers CD90, CD105, and CD146. Intriguingly, we found that the proportions of CD105 and CD146 were higher in FF-iPDLSCs than in OF-iPDLSCs (Fig. 2A). Almost no CD34-positive and CD45-positive cells were observed in either cell type (Fig. 2B). Additionally, we measured proliferation ability in OF/FF-iPDLSCs using a WST-1 assay. There was no significant difference in proliferation ability between cell types (Fig. 2C).
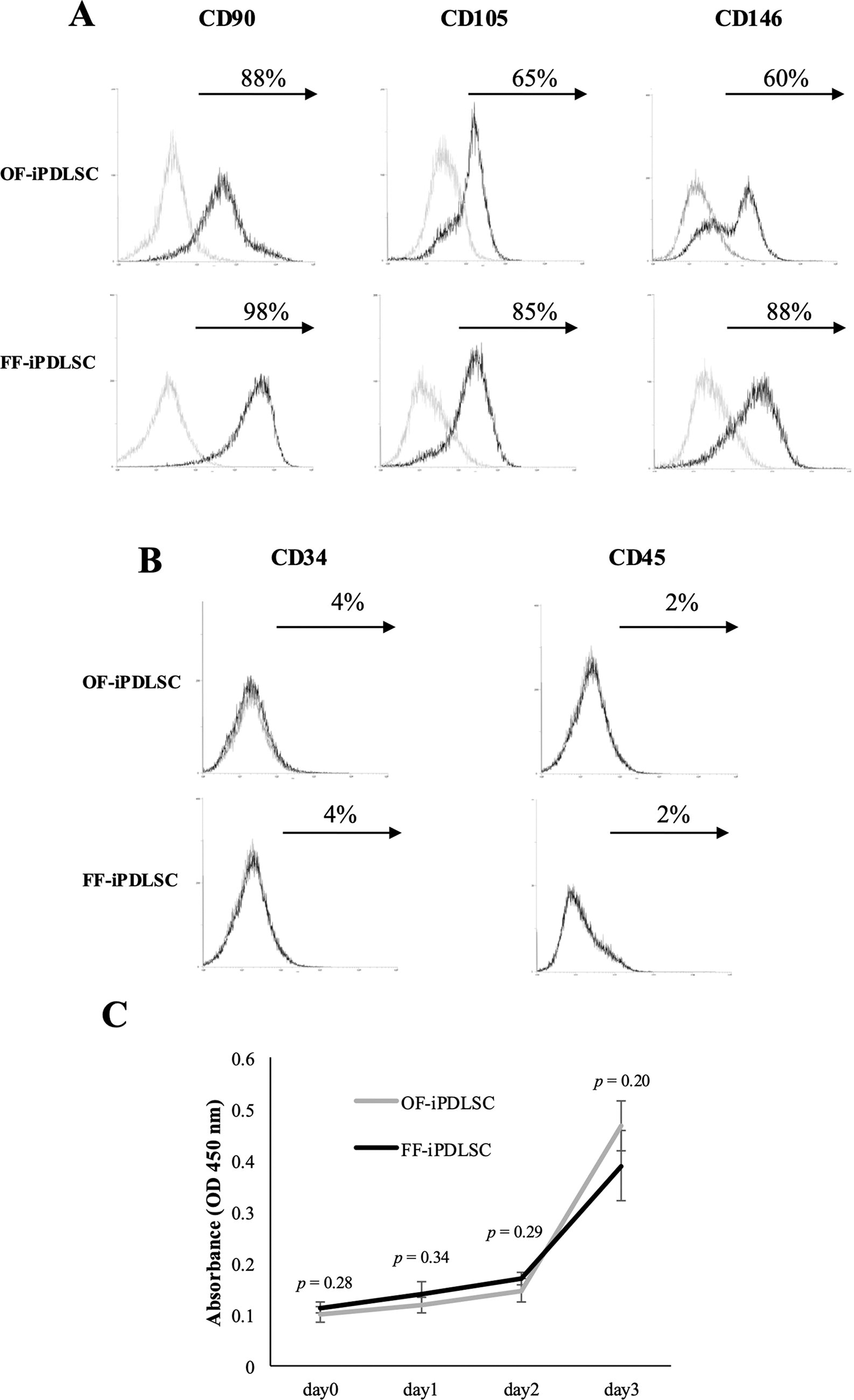
Expression of MSC markers and proliferation ability in OF/FF-iPDLSCs.
Multipotency of OF/FF-iPDLSCs
First, OF/FF-iPDLSCs were cultured in osteoblast differentiation medium and stained with Alizarin Red S to visualize mineralization (Fig. 3A). Both OF-iPDLSCs and FF-iPDLSCs showed significant staining compared to the control, but there was no difference between these two cell types (Fig. 3B). RT-PCR analysis revealed the expression of osteogenesis-related markers (BSP, BMP2, and sclerostin [SOST]) in each cell type and there was no significant difference between the cell types (Fig. 3C). Next, OF/FF-iPDLSCs were cultured in adipocyte differentiation medium. We stained these cells with Oil Red O and observed fatty lipids (Fig. 3D). Compared with OF-iPDLSCs, FF-iPDLSCs formed more fatty lipids (Fig. 3E). The expression levels of adipocyte-related markers (lipoprotein lipase [LPL], peroxisome proliferator-activated receptor-γ [PPARγ], and ADIPSIN) were higher in FF-iPDLSCs than in OF-iPDLSCs (Fig. 3F). These results suggested that FF-iPDLSCs had properties similar to OF-iPDLSCs in vitro.
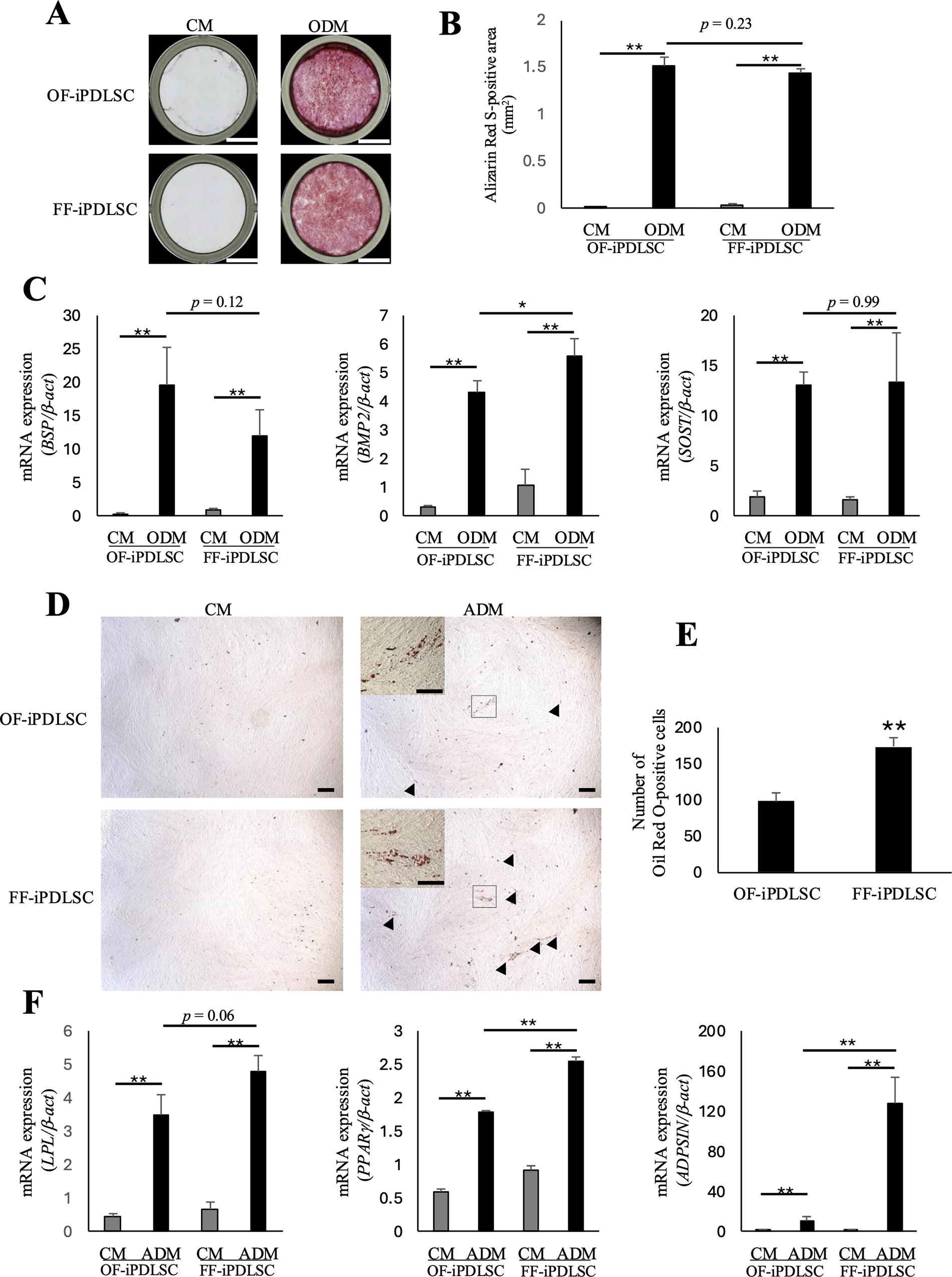
Multipotency of OF/FF-iPDLSCs.
Expression of PDL-related markers in OF/FF-iPDLSCs
To determine whether FF-iPDLSCs have characteristics similar to OF-iPDLSCs, we analyzed the expression levels of PDL-related markers (OPG, PLAP1, COL1, Mohawk [MKX], and fibrillin-2 [FBN2]) in OF/FF-iPDLSCs via RT-PCR (Fig. 4A). The expression levels of PDL-related markers were significantly higher in FF-iPDLSCs than in OF-iPDLSCs. The protein expression levels of OPG and COL1 also were higher in FF-iPDLSCs (Fig. 4B).

Expression of PDL-related markers in OF/FF-iPDLSCs.
Periodontal tissue formation of FF-iPDLSCs in vivo
Finally, we evaluated the periodontal tissue formation ability of FF-iPDLSCs in vivo. Eight weeks later after transplantation of FF-iPDLSCs, we observed fiber-like tissues connecting the β-TCP scaffolds (Fig. 5A). Fiber-like tissues (observed by Masson’s trichrome staining) exhibited COL1, COL3, and OPG expression (Fig. 5B–D). Immunostaining was performed using an antihuman OCN and OSX antibody, which identifies human bone/cementum; OCN and OSX-positive cells were observed around the scaffolds (Fig. 5E and F). In addition, immunostaining was performed using an anti-NuMA antibody that specifically stains human nuclei; this assay confirmed that FF-iPDLSCs were localized to the transplant site (Fig. 5G). We also confirmed co-localization of COL1 and NuMA (Supplementary Fig. S1) and found that COL1 was expressed not only in mouse cells but also in iPDLSCs. These results suggested that FF-iPDLSCs generated periodontal tissue-like structures in vivo. In control group, these tissues were not evident (Fig. 5I) and no expression of NuMA was observed (Fig. 5J).
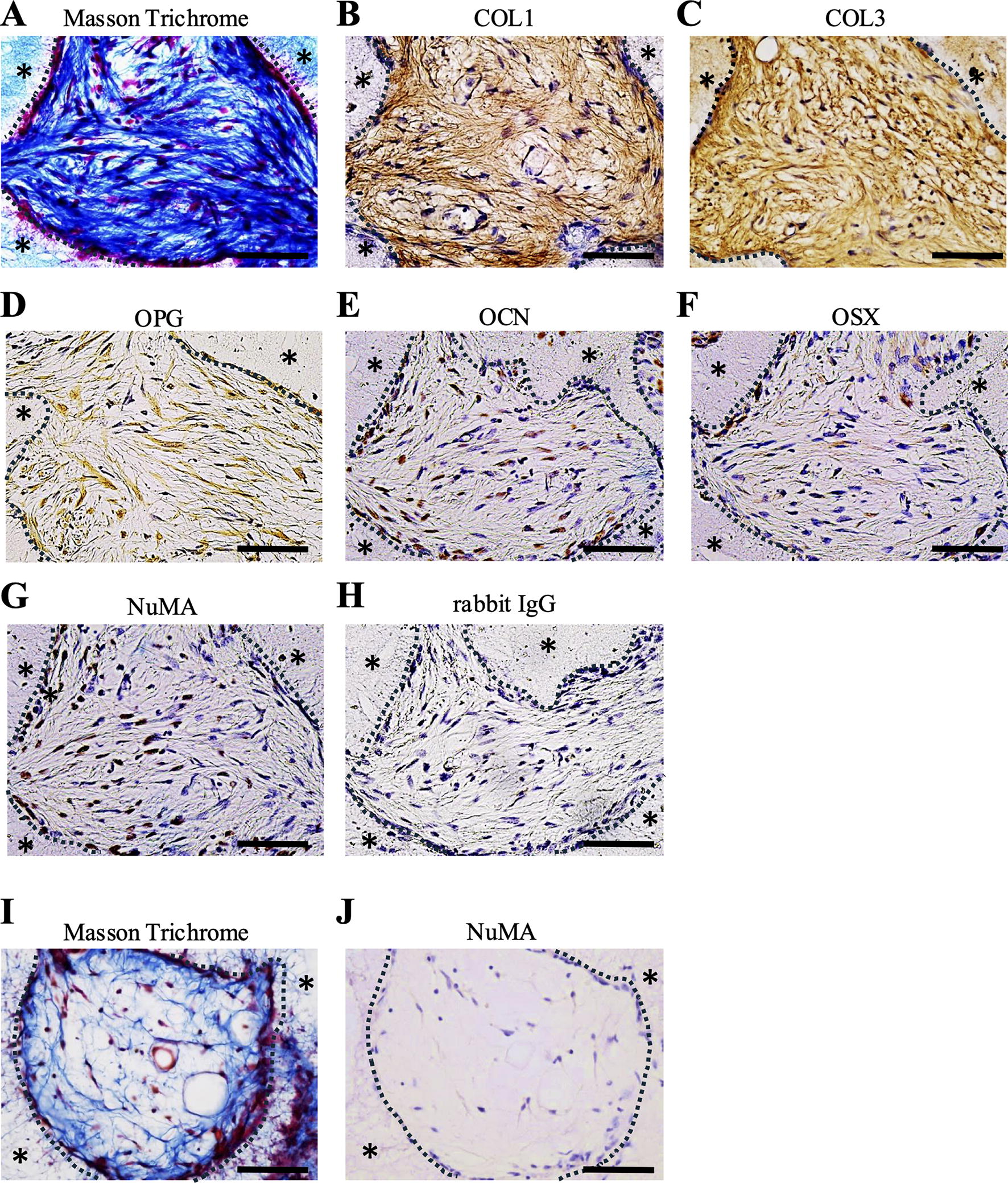
Differentiation potential of FF-iPDLSCs in vivo.
Discussion
In this study, we aimed to obtain FF-iPSC-derived PDLSCs that can be used in clinical applications. We found that FF-iPDLSCs exhibited characteristics similar to OF-iPDLSCs in vitro and formed PDL-like structures in vivo.
NCCs are stellate-shaped cells that express neural crest markers, such as p75NTR and Nestin. 15 –18 NCCs have also reported to differentiate into various cell types, such as craniofacial bone, Schwann, and smooth muscle cells. 19 In our previous study, we could not differentiate OF-iPDLSCs directly from OF-iPSCs. 12 Those results suggested that it was important to differentiate iPSCs into iNCCs, which could then be used for differentiation into iPDLSCs. In this study, FF-iPSC-derived FF-iNCCs had a stellate morphology and expressed neural crest markers similar to OF-iNCCs. These findings suggested that the iNCCs required for iPDLSC differentiation could also be obtained from FF-iPSCs.
The MSC markers CD105 and CD146 are involved in regulating MSC pluripotency and proliferation. 20 –22 A previous study of mouse dermal fibroblasts revealed that, compared with CD105-negative cells, CD105-positive cells had a greater capacity for differentiation into osteoblasts and adipocytes. 23 In addition, CD146 was regarded as a key marker of PDLSCs during their initial characterization, 3 and it has been reported that CD146-positive cells display multipotency and proliferation in various cell types, such as bone marrow MSCs and umbilical cord MSCs. 24 –26 FF-iPDLSCs, which abundantly expressed CD105 and CD146, demonstrated multipotency and proliferation; these results were consistent with previous reports. Moreover, the ability to differentiate into adipocytes was stronger among FF-iPDLSCs than among OF-iPDLSCs. These results might be related to the higher expression levels of CD105 and CD146 in FF-iPDLSCs than in OF-iPDLSCs.
PDL-related markers such as OPG, PLAP1, COL1, MKX, and FBN2 are known to be major components of PDL tissues. 27 –30 FF-iPDLSCs expressed higher levels of these markers compared with OF-iPDLSCs. Those results suggested that FF-iPDLSCs successfully differentiated into PDLSCs in vitro, similar to OF-iPDLSCs.
In this study, we focused on whether iNCCs differentiated from iPSCs in a FF condition could differentiate into PDLSC-like cells. However, we used HPDLC-derived ECM and serum medium in iPDLSC differentiation process. Using serum-free medium is crucial for safety and reliability in clinical applications, therefore completely xeno-free differentiation of iPDLSC is essential. We have previously hypothesized that the ECM derived from HPDLCs contains PDLSC-inducing factors. Thus, we have performed comprehensive analysis of HPDLCs and succeeded in identifying FBN2, one of the PDLSC-inducing factors. 31 In the future, we plan to identify PDLSC-inducing factors other than FBN2 and establish a method to induce FF-iPDLSCs without using HPDLCs and serum medium.
Previous reports have shown that OCN and COL1 are expressed during subcutaneous transplantation of PDLSCs. 32 –34 Cementoblasts and osteoblasts have been reported to express OCN and OSX and these results supported our results. 35,36 In our previous studies, we have demonstrated that COL3 and OPG, as well as COL1, are PDL markers. 37,38 Thus, it was suggested that FF-iPDLSCs formed bone/cementum- and PDL-like tissues in vivo and FF-iPDLSCs might be useful for periodontal tissue regeneration. However, to evaluate the periodontal tissue regeneration ability of FF-iPDLSCs, it is necessary to transplant these cells into injured periodontal tissue. Previous studies reported that scaffold materials such as collagen hydrogel, poly lactic-co-glycolic acid, and apatite-coated silk fibroin were used when transplanting cells into periodontal tissue. 39 –41 In addition, growth factors such as b-FGF, platelet-rich fibrin, and the bone morphogenetic factor 6 were suggested to promote periodontal tissue regeneration. 42 –45 In the future, it will be necessary to identify scaffolds and growth factors suitable for FF-iPDLSCs, then transplant these components into models of periodontal tissue injury.
Conclusions
In conclusion, we found that FF-iPDLSCs had properties similar to OF-iPDLSCs in vitro and formed periodontal tissue-like structures in vivo, suggesting that PDLSCs can differentiated induced from FF-iPSCs. These results may lead to the development of novel periodontal tissue regeneration therapies.
Footnotes
Acknowledgments
We thank Drs. Obata, Yoshida, Kaneko, Kinoshita, Kadowaki, Nagano, Safwan, Tashita, Toraya, Kidsen, and Bara for their great support in the preparation of this work and we appreciate the technical assistance from The Joint Use Laboratories, Faculty of Dental Science, Kyushu University. We also thank Michelle Kahmeyer-Gabbe, PhD, from the Edanz Group (![]() ) for editing a draft of this article.
) for editing a draft of this article.
Author Disclosure Statement
The authors declare that there is no conflict of interest regarding the publication of this article.
Funding
This work was financially supported by Grants-in-Aid for Scientific Research (Project Nos. JP21K09876, JP22H03270, JP23K09171) from the Japan Society for the Promotion of Science.
Supplementary Material
Supplementary Data S1
Supplementary Figure S1