
Abstract
Immunotherapy utilizes immune cells to target cancer and improves treatment outcomes with few side effects. Despite the effectiveness of immunotherapy, the limited availability of monocytes, which are essential for the differentiation of antigen-presenting cells, remains a major challenge. In this study, we developed a technique for inducing monocytes from hematopoietic stem and progenitor cells by using a serum-free (SF) medium supplemented with optimal concentrations of serum substitutes and cytokines. Three key serum substitutes, namely lipids, ascorbic acid, and β-glycerophosphate, were identified through factorial design screening, with their concentrations optimized through steepest ascent path analysis. Iscove’s modified Dulbecco’s medium was identified as the optimal basal medium. Long-term culturing confirmed the successful induction of CD14+CD16+ and CD14+CD16− monocytes. Functional assays validated the efficacy of this technique with comparable gene expression, cytokine secretion, phagocytosis ability, and T-cell stimulating ability between SF and serum-containing cultures. Under SF conditions, high expression levels of CD16 were detected, indicating the broad range of potential applications of CD16+ monocytes. Overall, this technique represents a feasible SF alternative for monocyte generation, with potential benefits for immunotherapy.
Introduction
Immunotherapy is a technique that involves utilizing immune cells for the treatment of cancer, and it is associated with several benefits, such as reduced invasiveness, minimal side effects, and low recurrence rates. 1 –3 This technique is extensively used in the management of cancer and involves utilizing antigen-presenting cells (APCs), such as dendritic cells (DCs), to initiate immune responses. Monocytes, identified as precursors to DCs and macrophages, play a key role in the differentiation of APCs, increasing the effectiveness of immunotherapy. 4,5 Despite the positive effects of monocytes, the differentiation of APCs, particularly in ex vivo cultures, is often impeded by apoptosis, which in turn reduces the number of viable cells. 6 To solve this problem, the number of progenitor cells should be increased. Nevertheless, monocytes constitute only 2%–8% of all leukocytes in human blood, representing a limited population of 2–8 × 105 monocytes per liter. As a result of cell loss during cell separation and isolation, obtaining a sufficient number of monocytes for immunotherapy can be a challenging process, potentially resulting in fewer than 104 cells. 7 In this study, we explored the feasibility of inducing the differentiation of monocytes from serum-free (SF) expanded hematopoietic stem and progenitor cells (HSPCs) by using an effective SF expansion method. We utilized the proliferative capacity of HSPCs during their differentiation into monocytes as a promising approach to address the shortage of monocytes required for the production of APCs.
In the adult human body, monocytes differentiate from HSPCs within the bone marrow and subsequently join peripheral blood in an immature state. 8 Compared with other types of blood cells, monocytes exhibit stronger endocytic capabilities. After briefly circulating for 2–3 days, monocytes migrate to peripheral tissues, where they undergo maturation, which is characterized by an increase in cell size, lysosomal content, and mitochondrial content. Upon activation, both monocytes and tissue macrophages release various cytokines, including interferons and interleukins, to coordinate immune responses. 8,9 Under inflammatory conditions, monocytes proliferate and participate in antigen uptake, subsequently process and present antigen, and promote B cell production of antibodies. 10,11 Despite being classified as blood cells, monocytes predominantly exert their functions within tissues and organs.
Monocytes typically express CD14 and CD16 as their primary antigens. Depending on these antigens, monocytes are divided into three subtypes: nonclassical monocytes (CD14+CD16++), intermediate monocytes (CD14++CD16+), and classical monocytes (CD14++CD16−). Classical monocytes constitute 80%–95% of the total population of monocytes, and they exhibit anti-inflammatory properties. Intermediate monocytes constitute 2%–11% of the total population of monocytes, and they exhibit proinflammatory properties. Among all three types of monocytes, nonclassical monocytes represent the smallest fraction, accounting for 2%–8% of the total population of monocytes. 11 –17 During the maturation process of monocytes, the expression of CD16 is typically upregulated, accompanied by a reduction in cellular dimensions and phagocytic efficacy. Notably, CD16+ monocytes, which have proinflammatory properties, demonstrate increased reactivity upon Toll-like receptor activation, which in turn increases the secretion of tumor necrosis factor (TNF), IL-1β, and IL-10. 10,18 These cells tend to accumulate in older individuals and under chronic inflammatory conditions.
In this study, we evaluated various serum substitutes and optimized their concentrations through factorial design (FD) combined with a steepest ascent path (SAP) technique. Our goal was to develop an optimal SF induction medium capable of generating a large number of functional monocytes from HSPCs. After being cultured for an extended period, an SF medium capable of differentiating expanded CD34+ HSPCs into CD14+ monocytes, as evidenced by previous studies, 19 –21 was developed under serum-containing (SC) media. After 10 days of culture, the expression of CD16 was monitored. A growth curve analysis revealed a plateau in cell growth by day 14, coinciding with an increase in the number of CD16+ cells. By day 21, the number of CD14+CD16+ cells peaked at approximately 40% and remained stable thereafter. Characterization of maturation revealed changes in the proportions of CD16+ cells and their endocytic activity. Specifically, older cells exhibited lower latex endocytosis by day 28 compared with day 14. Overall, our data confirmed the efficacy of the proposed SF monocyte induction medium in inducing HSPCs to produce a large number of monocytes. This medium can be used as a key source for the production of DCs and for expanding their potential applications in immunotherapy.
Material and Method
Purification of HSPCs
The sample collection procedure of cord blood was approved by the Institutional Review Board of Taoyuan General Hospital, Ministry of Health and Welfare, Taiwan (approval no. TYGH106078). Before any procedures were performed, written informed consent was obtained from each mother. Mononuclear cells were isolated from cord blood and purified through Ficoll-Paque density gradient centrifugation (Amersham Biosciences, Amersham, UK). Fresh HSPCs were isolated using a CD133 microbead kit on a VarioMACS device (Miltenyi Biotec) in accordance with the manufacturer’s instructions.
Monocyte differentiation medium
Two primary media were separately used for the expansion of HSPCs and the induction of monocytes. An SF medium previously developed for HSPC expansion was used. 22 This medium contained Iscove’s modified Dulbecco’s medium (IMDM) supplemented with serum substitutes including human serum albumin (HSA), transferrin, insulin, and 2-mercaptoethanol. It also contained a cytokine cocktail comprising a stem cell factor (SCF), thrombopoietin (TPO), an Fms-like tyrosine kinase 3 ligand (Flt-3L), IL-3, IL-6, a granulocyte colony-stimulating factor, and a granulocyte macrophage colony-stimulating factor (GM-CSF).
To achieve monocyte induction, a medium based on IMDM supplemented with either serum or serum substitutes was used. During the differentiation phase, a cytokine cocktail comprising 100 ng/mL GM-CSF, 14 ng/mL IL-3, 39 ng/mL Flt-3L, 12 ng/mL TPO, 79 ng/mL macrophage colony-stimulating factor (M-CSF), and 89 ng/mL SCF was added to the two media. The optimal serum substitute cocktail was found to be the cocktail that contained lipid mix (cat. no. L5146, Sigma-Aldrich. Containing cholesterol, methyl ester, polyoxyethylenesorbitan monooleate and
Experimental design of the SF medium formula
A hybrid approach involving fractional FD and an SAP technique was used to determine the optimal concentration of serum substitutes required to induce the production of monocytes from expanded HSPCs. To formulate a polynomial equation, the results obtained from FD were subjected to a regression analysis in Design-Expert software (Stat-Ease, Minneapolis, MN, USA). This equation was used to elucidate the correlation between each substitute and the resultant cell count, with a focus on CD14+ monocytes (cells/mL):
Following the coefficients derived from fractional FD, the concentration of each serum substitute was systematically increased in proportion according to the coefficients and difference of concentration. Experiments were conducted along this gradient until the cell yield reached a plateau or its maximum.
To develop an SF monocyte induction medium, serum substitutes such as lipids, ascorbic acid, β-glycerophosphate, hydrocortisone, and α-tocopherol were evaluated using FD. Subsequently, the concentration ratios of these substitutes were optimized using an SAP technique.
Flow cytometry
Induced CD14+ monocytes derived from expanded HSPCs were washed using Dulbecco’s phosphate-buffered saline (D-PBS; HyClone, Logan, UT, USA). Cell surface markers CD14 and CD16 were examined using flow cytometry (Accuri C6; BD Biosciences, Franklin Lakes, NJ, USA). Fluorescent monoclonal antibodies specific for CD14 and CD16 were purchased from Miltenyi Biotec. Monocytes were selected in accordance with the criterion of CD14+ cell expression.
Cell morphology
Scanning electron microscopy (SEM) was used to determine the cell morphology of the HSPCs-derived monocytes. After culture induction at designated time intervals, the induced monocytes were fixed using 4% glutaraldehyde (Sigma-Aldrich, St. Louis, MO, USA) and subjected to stepwise dehydration with an ethanol/water solution in the presence of progressively increasing concentrations of alcohol. Finally, the cells were subjected to gold sputter coating for 100 s and examined under a scanning electron microscope operated at 10 kV.
Endocytosis analysis
At various stages of induction, cells were collected and plated at a density of 2 × 105 cells/mL on 24-well plates (1 mL/well). After 1 mg/mL of dextran-fluorescein isothiocyanate (dextran-FITC; Sigma-Aldrich) or fluorescent latex beads (Sigma-Aldrich) was added, the mixture was incubated in an RPMI medium for 1 h at 37°C in the presence of 5% CO2. After incubation, the cells were stained with CD14-phycoerythrin (Miltenyi Biotec). Endocytic activity was evaluated by quantifying the uptake of dextran-FITC and latex beads in the CD14+ cells through flow cytometry.
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qRT-PCR) was used to compare the relative gene expression levels of six inflammatory cytokines. After RNA was reverse-transcribed into cDNA, cDNA amplification was conducted using RT-PCR with a KAPA SYBR Fast Master Mix (Thermo Fisher Scientific, Waltham, MA, USA). Gene expression levels were normalized to the level of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), an internal housekeeping gene. Table 1 lists the sequences of the primers used in this study.
Primer Sequences of Monocyte-Related Genes
IL, Interleukin; TNF, tumor necrosis factor.
Cytokine secretion
Monocytes typically regulate immune function by secreting cytokines. In this study, the levels of IL-10, TNF-α, and IL-12p70 secreted by monocytes were measured using a human inflammatory cytokine cytometric bead array kit (BD Biosciences). After differentiation for 7, 14, and 28 days, the secretion profiles of these cytokines were evaluated using flow cytometry in accordance with the manufacturer’s protocol.
Mixed leukocyte reaction
To evaluate the allogeneic stimulatory potential of distinct monocyte populations, induced monocytes were collected on days 7, 14, and 21 after differentiation. CD3+ T cells were isolated from cord blood mononuclear cells using CD3 microbeads (Miltenyi Biotec Gmbh) and the VarioMACS Separator. Then, CD3+ T cells were stained with carboxyfluorescein succinimidyl ester (CFSE, Sigma-Aldrich) to serve as the target cells. After CD3+ T cells stained with CFSE (Sigma-Aldrich) were washed with D-PBS, they were cocultured with these induced monocytes (at monocyte to T cell ratios of 1:1, 1:2, and 1:5) in an RPMI medium containing 10% fetal bovine serum (FBS) at 37°C for 4 days in the presence of 5% CO2. Finally, all cells were collected, and the ratio of proliferating CD3+ T cells to the total population of T cells was determined using flow cytometry.
Statistical analysis
All experiments were independently conducted at least three times, and the results are presented as the mean ± standard error. A p value less than 0.05 was considered statistically significant, as determined by a paired-samples t-test, with significant values represented by an asterisk.
Results
Monocyte induction from HSPCs-serum substitutes screening
Synthesizing an appropriate SF medium is essential in clinical applications. In a previous study, we synthesized an SF medium (HIT2) containing four serum substitutes suitable for ex vivo HSPC expansion: 1.5 g/L HSA, 60 μg/mL transferrin, 4.39 μg/mL insulin, and 25.94 μM 2-mercaptoethanol.
22
We also synthesized a cytokine-optimized medium for monocyte differentiation from HSPCs under SC conditions, which contained IMDM supplemented with 20% FBS and a cytokine cocktail (100 ng/mL GM-CSF, 89 ng/mL SCF, 79 ng/mL M-CSF, 39 ng/mL Flt-3L, 14 ng/mL IL-3, and 12 ng/mL TPO).
19
In the present study, we systematically evaluated 19 serum substitutes (biotin, folic acid,
Groups with high CD14+ cell performance were identified in single-addition experiments (data not shown). After screening, five serum substitutes were identified as potentially essential for the generation of monocytes: hydrocortisone, lipids, ascorbic acid, α-tocopherol, and β-glycerophosphate. After the effects of these substitutes were evaluated, their optimal concentration ratios were determined using two-level fractional FD (Table 2). The resulting FD data were analyzed using Design-Expert software, which generated a formula indicating the correlation between the number of CD14+ cells and the effects of each substitute:
Matrix of 25-1 Fractional Factorial Design and Experimental Results of Serum Substitutes for CD14+ Cell Induction from Hematopoietic Stem and Progenitor Cells
The initial cell seeding density was 1 × 105 serum-free expanded HSCs/mL in IMDM supplemented with 1.5 g/L human serum albumin, 4.39 μg/mL insulin, 60 μg/mL transferrin, and 25.94 μM 2-mercaptoethanol.
+1: the concentration of adding serum substitute was indicated in the in parentheses; −1: no added serum substitute.
Cell density at day 7 (n = 5).
IMDM, Iscove’s modified Dulbecco’s medium.
Both hydrocortisone and α-tocopherol exhibited negative effects, as well as negative ecoefficiency, when combined with other cocktail substitutes for the generation of CD14+ monocytes. By contrast, lipids, ascorbic acid, and β-glycerophosphate exhibited high potential, with positive ecoefficiency, for increasing the production of CD14+ monocytes. Finally, the optimal cocktail ratio was refined using an SAP technique for concentration optimization.
Monocyte induction from HSPCs-concentration optimization of serum substitutes
Fractional FD was used to determine the concentration ratios of lipids, ascorbic acid, and β-glycerophosphate from the coefficients of Equation (1). An SAP technique was used to determine the optimal concentrations of these components in relation to the HIT2 criteria under SF conditions. This technique involved an initial concentration of 0 mM for each substitute, progressing over nine steps to a maximum concentration of 100 mM for β-glycerophosphate. Table 3 presents the concentrations of serum substitutes and the corresponding CD14+ cell numbers.
Concentrations of Serum Substitutes with Steepest Ascent Path, Total Cell, and CD14+ Monocyte Growth
Value in the parenthesis is the standard deviation; the initial seed density was 1 × 105 serum-free expanded CD34+ cells/mL in IMDM supplemented with 1.5 g/L human serum albumin, 4.39 μg/mL insulin, 60 μg/mL transferrin, and 25.94 μM 2-mercaptoethanol.
Cell density at day 7 (n = 5).
As shown in Table 3, the number of CD14+ cells uniformly increased with increasing concentrations of serum substitutes, until it reached a critical value at step 5 with 0.063% lipids, 0.063 mM ascorbic acid, and 30 mM β-glycerophosphate. After step 5, the number of CD14+ cells decreased, indicating the presence of an optimal concentration range.
To reexamine the effects of hydrocortisone and α-tocopherol on the induction of CD14+ monocytes from HSPCs under SF conditions, these components were added at the concentrations identified at step 5 (Fig. 1 and Supplementary Fig. S1). The results indicated that hydrocortisone and α-tocopherol were associated with significantly lower production of CD14+ cells compared with the production levels observed at step 5 (p < 0.05), which involved the same concentrations of lipids, ascorbic acid, and β-glycerophosphate.

Culture conditions for the production of CD14+ monocytes from HSPCs. The initial number of HSPCs was 1 × 105. The number of induced monocytes was counted on day 7 in IMDM supplemented with six cytokines (GM-CSF, Flt-3L, TPO, M-CSF, SCF, and IL-3) and an optimal serum substitute cocktail at step 5 (1.5 g/L HSA, 4.39 μg/mL insulin, 60 μg/mL transferrin, 25.94 μM 2-mercaptoethanol, 0.063% lipids, 0.063 mM ascorbic acid, and 30 mM β-glycerophosphate). The negative effects of hydrocortisone (1 μM) and α-tocopherol (4 μg/mL) on monocyte generation were examined: (+) addition, (−) no addition. *Significant difference at p < 0.05 compared with other groups (n = 5). Flt-3L, Fms-like tyrosine kinase 3 ligand; GM-CSF, granulocyte macrophage colony-stimulating factor; HSA, human serum albumin; HSPCs, hematopoietic stem and progenitor cells; IL-3, Interleukin-3; IMDM, Iscove’s modified Dulbecco’s medium; M-CSF, macrophage colony-stimulating factor; SCF, stem cell factor; TPO, thrombopoietin.
In summary, our findings confirmed that the elimination of hydrocortisone and α-tocopherol, as identified by FD, had no effect on the production of CD14+ cells. In IMDM supplemented with HIT2, lipids, ascorbic acid, and β-glycerophosphate played a key role in the production of CD14+ cells. The optimal concentrations of serum substitutes for the induction of monocytes from HSPCs were determined to be 1.5 g/L HSA, 60 μg/mL transferrin, 4.39 μg/mL insulin, 25.94 μM 2-mercaptoethanol, 0.063% lipids, 0.063 mM ascorbic acid, and 30 mM β-glycerophosphate.
Monocyte induction from HSPCs-basal medium comparison
Generally, the presence of a serum substitute cocktail capable of producing CD14+ cells underscores the importance of selecting an optimal basal medium. In this study, we compared seven basal media to determine their compatibility with monocyte induction and serum substitute cocktails consisting of HIT2, lipids, ascorbic acid, and β-glycerophosphate (Fig. 2 and Supplementary Fig. S2). The results indicated that, in terms of the number of CD14+ cells produced with each basal medium, IMDM had the highest efficacy in promoting the production of monocytes. After a modified SF medium was synthesized for CD14+ cell induction, functional assays were conducted to ensure that the generated CD14+ cells exhibited the typical characteristics of functional monocytes. These assays provided valuable insights into the functionality and effectiveness of the induced monocytes, confirming their suitability for subsequent therapeutic applications.
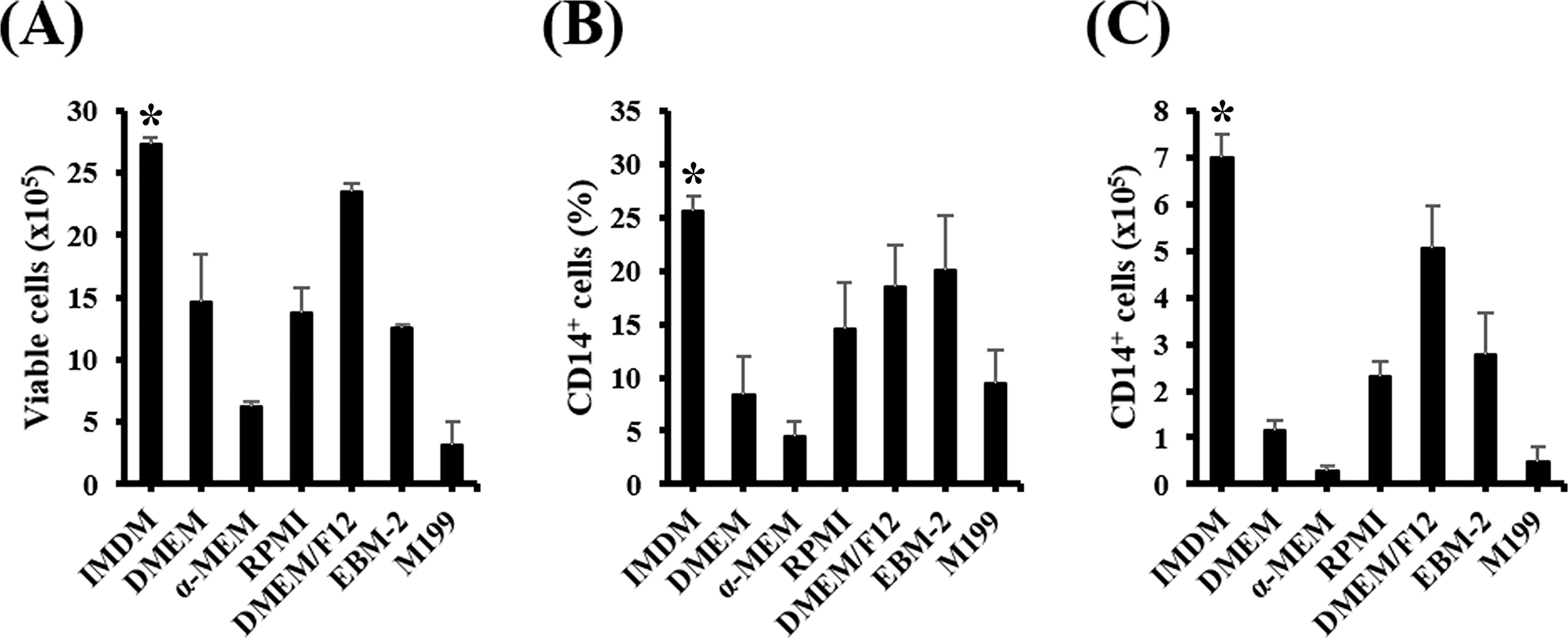
Comparison of the effects of different basal media on monocyte induction from HSPCs. In total, 1 × 105 HSPCs were cultured in different basal media supplemented with six cytokines (GM-CSF, Flt-3L, TPO, M-CSF, SCF, and IL-3) and an optimal serum substitute cocktail at step 5 (1.5 g/L HSA, 4.39 μg/mL insulin, 60 μg/mL transferrin, 25.94 μM 2-mercaptoethanol, 0.063% lipids, 0.063 mM ascorbic acid, and 30 mM β-glycerophosphate). After 7 days of culture induction,
Long-term culture of monocyte induction
To evaluate the efficacy of our monocyte induction medium, a long-term culture was conducted to monitor the simultaneous expression of the monocyte-presenting markers CD14 and CD16. A culture was initiated using 1 × 105 HSPCs/mL seeded on a 24-well plate (1 mL optimal monocyte induction medium per well). The medium was changed twice per week, and the cell density was adjusted to 1 × 105 cells/mL every week. Cell numbers were counted and compared to those obtained with an SC medium, and the culture was maintained until no further cell expansion was observed (Fig. 3).
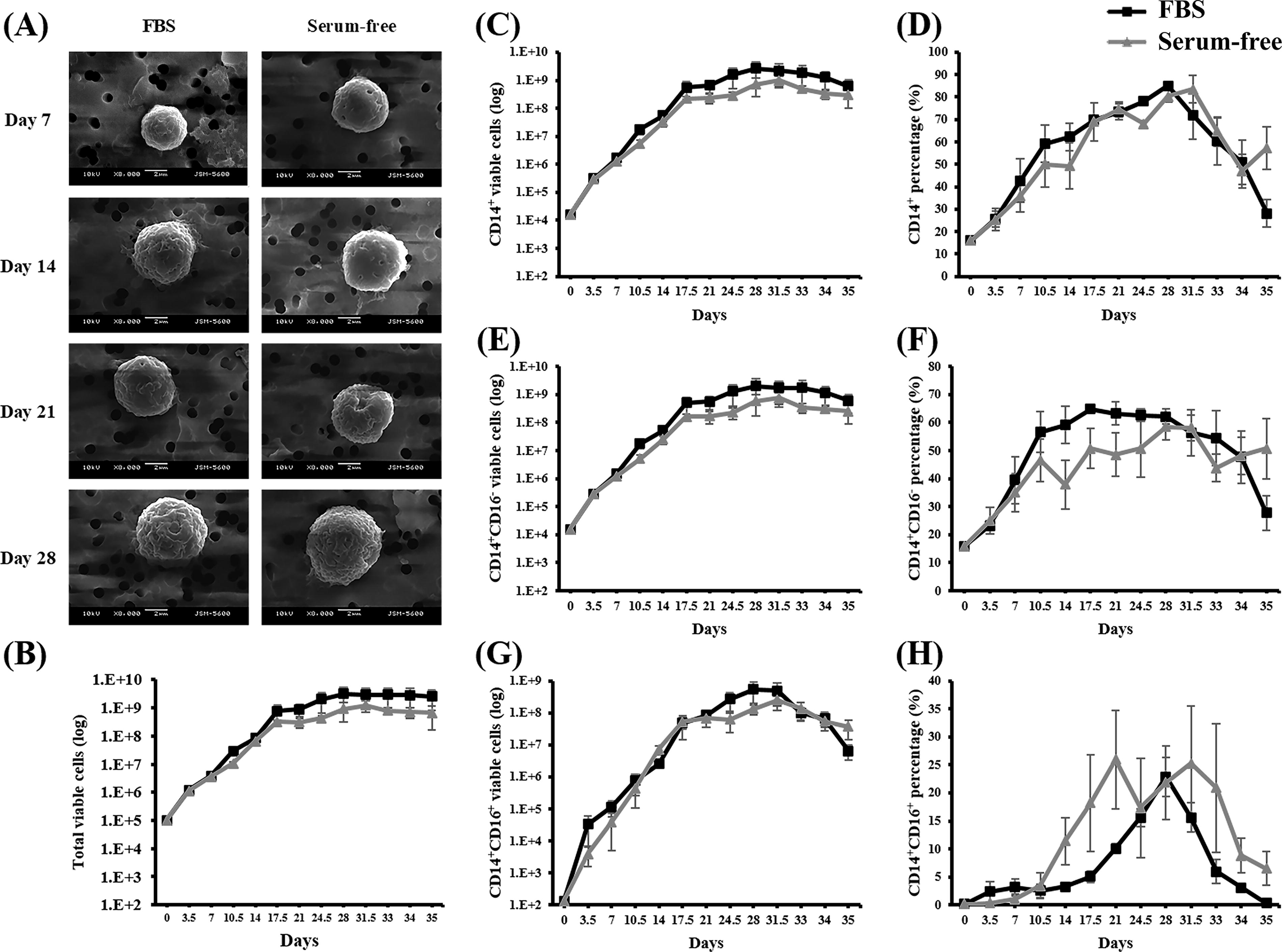
Long-term growth kinetics of monocytes induced from HSPCs in optimal SF and SC induction media. Monocytes were induced from 1 × 105 expanded HSPCs under SC (FBS group, 20% FBS, black lines) and SF (optimal SF monocyte induction medium, gray lines) conditions. The media were changed twice per week, and the cell density was adjusted to 1 × 105 cells/mL every 7 days. The induced cells were evaluated at the indicated time points.
According to the total cell counts, the proposed medium significantly promoted cell expansion, reaching over 2000 times the initial seeded cell count between days 17.5 and 21 (Fig. 3B). However, the rate of growth began to decrease thereafter, followed by a stationary phase. The maximum number of viable cells was recorded on day 31.5, which was followed by a substantial increase in cell death. Consequently, the culture was terminated on day 35, during the cell death phase.
Analysis of the expression ratios of CD14 and CD16 revealed the uniform induction of CD14 expression for 28 days (Fig. 3C and D). Before day 17.5, a significant increase was observed in the CD14+CD16− subset, followed by stabilization (Fig. 3E and F). By contrast, the CD14+CD16+ subset became evident by day 14, peaking on day 21 (Fig. 3G and H). These results indicated that the optimal monocyte induction medium effectively induced the differentiation of HSPCs into monocytes, mimicking the in vivo differentiation process ex vivo. In addition, it is worth noting that all growth kinetics and CD14/CD expressions of the induced cells cultured under SC and SF conditions were highly similar (no significant difference, p > 0.05). These results demonstrated that serum substitute cocktails (HIT2, lipids, ascorbic acid, and β-glycerophosphate) can replace the role of serum in monocyte differentiation.
After the induced cells were cultured for 7, 14, and 28 days in an SC or SF medium, their morphology was examined using SEM (Fig. 3A). The results indicated that these two conditions were associated with similar spherical cell shapes, with sizes increasing over time. Wrinkled structures were observed on the cell surfaces. All cells exhibited a uniform morphology, suggesting complete monocyte differentiation after culture induction.
Endocytosis ability
To provide insights into the mechanisms of cellular uptake, the endocytic capabilities of dextran-FITC for small molecules and latex beads for large molecules were evaluated. The results of the dextran-FITC assay indicated uniform endocytic capabilities across the SC and SF cultures, regardless of culture duration. The two groups exhibited similar percentages and mean fluorescence intensities (MFIs) for the uptake of small molecules, indicating constant performance under various conditions (Fig. 4A, C, and D, no significant difference, p > 0.05). By contrast, variations were observed in the uptake of larger molecules, as evidenced by our latex bead assays (Fig. 4B, E, and F). During the early stages (day 7 and day 14) of the long-term culture, the cells cultured under SF conditions exhibited marginally higher MFIs compared with those cultured under SC conditions (p < 0.05). However, by day 28, the monocytes induced under SC conditions exhibited slightly high MFIs (p < 0.05). Unlike small molecules, the two groups exhibited a similar pattern of increased endocytosis at first, followed by a decrease during the later stages of culture.
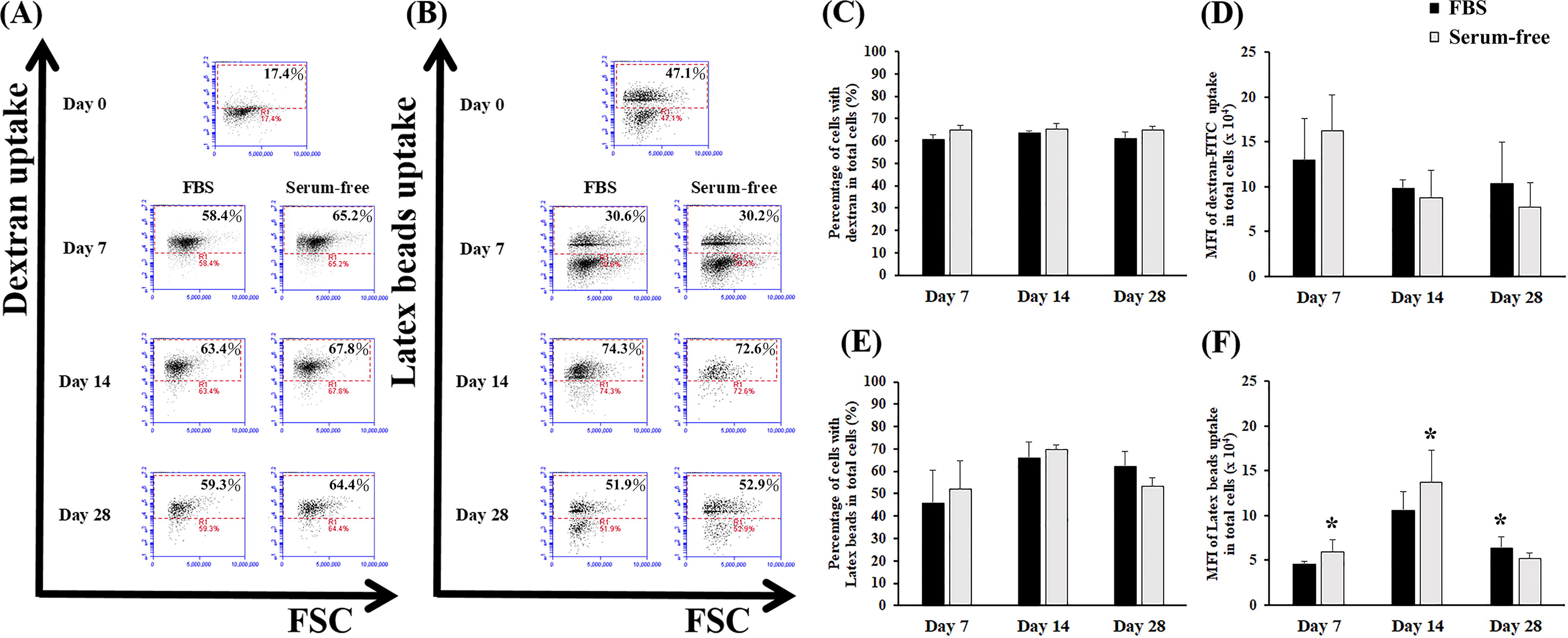
Endocytosis of induced CD14+ monocytes in the optimal SF induction medium and SC induction medium. Monocytes were induced from expanded HSPCs under SC (FBS group, 20% FBS, black bars) and SF (optimal SF monocyte induction medium, gray bars) conditions. After induction, the cells were collected at the indicated time points and treated with (A, C, D) dextran-FITC or (B, E, F) latex beads for endocytic analysis. Representative fluorescence expression of cells with
Taken together, our findings indicate that induced monocytes cultured under SF and SC conditions exhibit similar endocytic capabilities for small molecules. Nevertheless, slight differences were observed in the uptake of larger molecules between the two conditions. Overall, the uniform endocytic capabilities observed in induced monocytes under SF and SC conditions indicate that the proposed serum substitute cocktail can be used as an alternative to serum, with similar efficacy in inducing monocyte function.
Quantitative real-time polymerase chain reaction
Six monocyte-related genes, namely TNF-α, IL-1β, IL-6, IL-8, IL-10, and IL-12, were examined using qRT-PCR and compared to GAPDH under SC and SF conditions after 14 days of induction (Fig. 5). Given that sufficient marker expression and endocytosis were observed, expanded HSPCs were induced to functional monocytes after a minimum of 2 weeks of differentiation. On day 14, the cells were analyzed for inflammatory cytokines. Compared with the SC conditions, the SF conditions were associated with higher expression of IL-6 and IL-12, which typically have proinflammatory properties. However, no significant difference was observed in TNF-α, IL-1β, IL-8, or IL-10.

qRT-PCR analysis of monocytes induced from HSPCs in the optimal SF induction medium and SC induction medium. Monocytes were induced from expanded HSPCs under SC (FBS group, 20% FBS) and SF (optimal SF monocyte induction medium) conditions. After 14 days of induction, the cells were collected at the indicated time points and subjected to a qRT-PCR analysis to determine the expression levels of monocyte-related genes (IL-1β, IL-6, IL-8, IL-10, IL-12, and TNF). The expression of each targeted gene was normalized to that of expanded HSPCs on day 0. *Significant difference at p < 0.05 compared with the corresponding SC group under the same conditions (n = 3). TNF, tumor necrosis factor; qRT-PCR, quantitative real-time polymerase chain reaction.
Cytokine secretion
Inflammatory cytokine levels were measured on days 7, 14, and 28 of monocyte induction. Cultures were initiated with HSPCs treated with either an SC or an SF monocyte induction medium. Subsequently, the cells were transferred to a cytokine-free medium, and the concentrations of IL-10, IL-12p70, and TNF-α were determined (Fig. 6). Similar levels of IL-12p70 and TNF-α were observed under the SC and SF conditions, with the exception of TNF under the SC conditions on day 7. By contrast, the levels of IL-10 levels, which are correlated with the expression of CD16, were initially high under the SC conditions, and they increased under the SF conditions after 14 days of culture. Consistent with the ratio of CD14 and CD16 observed in the long-term induction culture, these findings indicate that the SF culturing method may delay the maturation of monocytes derived from HSPCs.
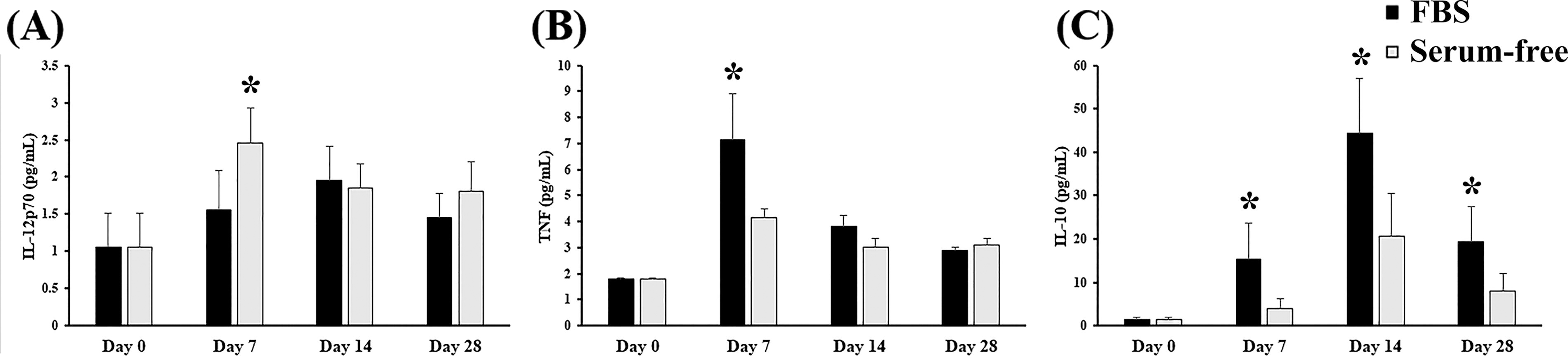
Cytokine secretion in monocytes induced from HSPCs in the optimal SF induction medium and SC induction medium. Monocytes were induced from 1 × 105 expanded HSPCs under SC (FBS group, 20% FBS, black bars) and SF (optimal SF monocyte induction medium, gray bars) conditions. The media were changed twice per week, and the cell density was adjusted to 1 × 105 cells/mL every 7 days. After induction, the cells were collected at the indicated time points, and the amounts of
Mixed lymphocyte reaction
Monocytes trigger T-cell proliferation, with increased T-cell counts indicating enhanced stimulation capabilities. The CFSE assay, which is used to identify halved MFI peaks, serves as an indicator of increased cell proliferation (Fig. 7).

Mixed lymphocyte reaction of monocytes induced from HSPCs in the optimal SF induction medium and SC induction medium. Monocytes were induced from expanded HSPCs under SC (FBS group, 20% FBS) and SF (SF group, optimal SF monocyte induction medium) conditions. After induction, CD14+ monocytes were collected and isolated using anti-CD14 microbeads on a VarioMACS separator at the indicated time points. Subsequently, the isolated CD14+ monocytes were co-cultured with allogenic CFSE-stained CD3+ T cells at ratios of 1:1, 1:2, and 1:5 for mixed leukocyte reactions. The initial CD3+ T-cell density was 1 × 104 cells/mL. After 96 h of coculturing, all cells were harvested, and proliferating CD3+ T cells were identified using flow cytometry.
In this study, direct cell counting revealed that the proliferative capacity of the monocytes increased with the duration of HSPCs differentiating into monocytes until day 21 (Fig. 7B). Compared with the SC conditions, the SF conditions were associated with higher potential for stimulation (p < 0.05). However, by day 28, the stimulation potential under the two conditions decreased. These findings were supported by a CFSE assay (Fig. 7A), suggesting that the monocytes derived from HSPCs effectively stimulated T-cell proliferation during the initial 21-day induction period. Despite their initial activity, by the end of the 28-day culture period, their capacity to promote T-cell expansion decreased. Nevertheless, both induced monocytes from SC and SF media supported the expansion of T cells, as evidenced by the CFSE assay.
Discussion
In this study, we developed an optimized SF monocyte induction medium by using IMDM. This medium contains six cytokines (GM-CSF, M-CSF, SCF, IL-3, Flt-3L, and TPO) and seven serum substitutes (HSA, insulin, transferrin, 2-mercaptoethanol, lipids, ascorbic acid, and β-glycerophosphate) used to replicate the functions of serum in the production of monocytes. Our goal was to facilitate the differentiation of expanded HSPCs into monocytes. In total, 23 potential serum substitutes were thoroughly screened to evaluate their efficacy in monocyte induction, considering both cell numbers and the proportion of CD14+ cells. Lipids, ascorbic acid, and β-glycerophosphate were identified as the most effective candidates, and their optimal concentrations required for successful monocyte induction were determined.
Serum albumin serves as a carrier for lipids and fatty acids in the human body. When the body requires energy or other resources, adipocytes release fatty acids into the bloodstream. This process is essential in detoxification and in maintaining pH balance. 23,24 Insulin is a soluble protein and cytokine that plays a pivotal role in regulating cell metabolism. It enhances the function of many cells and is produced by the beta cells of the islets of Langerhans. Insulin is primarily involved in the metabolism of sugar and the maintenance of blood sugar balance. It is also used in the treatment of diabetes. 23,25 Transferrin is the primary iron-containing protein in plasma. It promotes the expansion of many cell types and facilitates the transport of iron released from the degradation of erythrocytes and digestion in the alimentary canal. 23 2-Mercaptoethanol is an organic compound that contains ethylene glycol and ethanedithiol functional groups. This compound serves both as a reducing agent for disulfide bonds and as an antioxidant. It also aids in mitigating oxygen toxicity, thereby promoting cell expansion. 26,27 Cholesterol, free fatty acids, and phosphoric acid combine to form lipids, which are fundamental components for all cells. Cells typically utilize lipids as a major source of energy or as precursors for biosynthesis. 28,29 Ascorbic acid, commonly referred to as vitamin C, serves as both an antioxidant and a pH regulator within physiological environments. It also plays a pivotal role in modulating the activation and expansion of immune cells, such as T cells. 28 β-Glycerophosphate is a serine—threonine phosphate enzyme that utilizes serine as a source of phosphate for cell growth and recombinant protein synthesis. Its addition to media helps reduce precipitation at high pH levels. 30 β-Glycerophosphate and Lys phosphatidylcholine are fundamental components of metabolic pathways and serve as major intermediates in lipid metabolism. Lysophosphatidylcholine either activates or enhances the chemotactic response of monocytes. 31
CD16, also referred to as FcγRIII, is a receptor expressed on the surface of various immune cells, including monocytes. Its expression is usually upregulated in response to inflammatory signals, including those from the cell culture environment. One potential pathway for the upregulation of CD16 expression involves the activation of Toll-like receptors on monocytes. When lipids, ascorbic acid, and β-glycerophosphate combine, they presumably create a microenvironment that either upregulates the expression of CD16 or exerts a synergistic effect on monocyte differentiation and CD16 expression. CD16+ monocytes represent a mature stage of myeloid differentiation with a microphage-like and DC-like transcription program, whereas CD16− monocytes closely resemble a common myeloid precursor, 32 suggesting a developmental relationship between different subsets of monocytes. These different subtypes represent sequential stages of monocyte differentiation. 33
Serum, commonly used as a cell culture supplement, promotes the generation of CD14+ cells. The development of a SF medium for monocyte induction is a crucial advancement in cell culture technology. Traditional SC media can introduce variability and potential contaminants, hindering reproducibility and scalability. SF media offer several advantages, including the defined composition, which allows for precise control of the culture environment, facilitating optimization and standardization. SF media could also reduce batch-to-batch variation by eliminating serum, which can vary in composition, ensures greater consistency in experimental outcomes. For scalability, SF media are better suited for large-scale production, making them essential for industrial applications. In other side, it could also reduce the risk of contamination by removing a potential source of biological contaminants, SF media improve the safety and reliability of cell culture processes. In this study, our growth curves and CD14 and CD16 ratios indicated that all cells exhibited similar growth patterns under optimal SF conditions. Although the SC and SF conditions were associated with similar CD14 ratios, the SC conditions induced a larger number of CD16− cells during the early stages of long-term culture induction. However, by day 28, this ratio decreased, with the CD14+CD16− cells reaching similar or lower levels compared with those observed under the SF conditions. By contrast, the CD14+CD16+ cells demonstrated opposite trends, with the SF conditions maintaining a high ratio of double-positive cells until day 28, leading to a crossover on the growth curves. Despite these findings, the SF conditions demonstrated considerable potential in inducing CD14+CD16+ cells at any given time point. In terms of the production of CD14+CD16+ cells, the SC conditions appeared to induce a more mature phenotype compared with the SF conditions. This maturity was reflected in the cells’ endocytic capability, with increasing CD16 expression correlating with phagocytic activity. Consequently, the induced monocytes exhibited a lower phagocytic capability under the SC conditions than under the SF conditions. In addition, monocyte maturity was associated with an increased secretion of inflammatory cytokines. Although the cells cultured under the SC conditions initially secreted a larger number of cytokines than those secreted under the SF conditions, in some instances, the cells cultured under the SF conditions caught up after long-term culture induction, suggesting a potential delay in the maturation process of monocytes when cultured in the absence of serum.
Under the SF conditions, the induced monocytes demonstrated higher endocytic activity and co-stimulation capabilities than those observed under the SC conditions. As an alternative to serum, the cells cultured under the SF conditions exhibited high T-cell costimulation activity with increasing monocyte to T-cell ratios. It is worth noting that serum contains many complex biomolecules, such as various types of hormones, growth factors, binding proteins, cytokines, and antibodies. These biomolecules may have different effects on cell behavior, such as cell proliferation, differentiation, migration, and function. To avoid interference from serum, a SF and cytokine-optimized medium was developed for monocyte differentiation from HSCs in this study. The reason why the performance of mixed lymphocyte reaction of induced monocytes under SC condition lower than that under SF condition may be due to some biomolecules contained in the serum. Certain biomolecules, such as prostaglandin E1, IL-16, IL-18, and VACM-1 have been demonstrated to show the inhibitive effects on mixed lymphocyte reaction. 34 –37 According to the secretion patterns of cytokines, the majority of cytokines were expressed at similar levels under the SC and SF conditions, with the exception of IL-6, which was upregulated in the monocytes induced under the SF conditions during PCR, consistent with its proinflammatory properties. Generally, IL-6 plays a pivotal role in the activation of the immune system by modulating the activity of various immune cells. Our findings jointly indicate that the SF culturing method can generate functional monocytes capable of catching and presenting antigens, which are crucial for the generation of functional T cells. Thus, the SF culturing method not only maintains but also presumably enhances the immunological function of monocytes, making it a promising alternative to traditional SC culturing methods.
In a previous study, two distinct subsets of monocytes were identified in a mouse model: Ly6ChighCCR2highGR1+CX3CR1low, which resembles human CD16− monocytes, and Ly6ClowCCR2lowGR1−CX3CR1high, which resembles human CD16+ monocytes. Compared with the Ly6ChighCCR2highGR1+CX3CR1low subset, the Ly6ClowCCR2lowGR1−CX3CR1high subset exhibited a substantially more mature phenotype. 33,38,39 According to the results of a previous transcriptome analysis, CD16− monocytes are typically derived from a common granulocyte—macrophage precursor. Subsequently, they differentiate into CD16+ monocytes, which exhibit a more mature phenotype closely resembling that of macrophages and DCs. 32 CD16+ monocytes exhibit high expression levels of transcripts encoding cathepsin L. This high expression contributes to phagocytic—endocytic proteolysis, which in turn facilitates antigen presentation by DCs. 40 According to the literature, monocytes cultured in optimal SF media have considerable potential to differentiate into DCs, thereby serving as APCs in immunotherapy. In addition, CD16− and CD16+ monocytes may induce the differentiation of Th1 and Th2. 41 Specifically, CD16+ monocytes express IL-12Rβ1, which facilitates the polarization of Th1, whereas CD16− monocytes express IL-6 and IL-13, which facilitate the polarization of Th2. 32 In this study, we observed a slight increase in the level of IL-12p70 when we used an SF method, which in turn generated a large number of CD16+ cells. However, we also observed a high level of IL-6. Therefore, to validate the function of cells cultured under SF conditions, further research is required to confirm whether serum substitutes and cytokine cocktails exhibit synergistic effects.
Conclusion
In this study, we successfully developed an effective technique for the induction of expanded HSPCs into monocytes in an SF medium enriched with a specific cytokine cocktail and selected serum substitutes. According to our findings, the monocytes cultured under the SF conditions exhibited a similar function to that of monocytes cultured in conventional SC media, which are commonly used in monocyte cultivation. These findings underscore the efficacy and potential of SF culturing in generating functional monocytes suitable for various applications. Notably, our optimized SF culturing method increased the expression of CD16 on the surface of monocytes. Overall, this novel technique can expand the potential applications of monocytes, particularly in scenarios in which traditional monocyte culturing methods may be inadequate or may be associated with limitations. Culturing monocytes under SF conditions offers a promising alternative with similar functionality, thereby laying the foundation for further advancements in biomedical research.
Ethics Approval and Consent to Participate
The study was approved by the institutional review board of Taoyuan General Hospital, Ministry of Health and Welfare, Taiwan with the IRB number TYGH106078. The study population consisted of 10 cord blood collection from pregnant women, and all of whom provided written informed consent.
Footnotes
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
Funding Information
This work was supported in part by the
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2