
Abstract
Spiral ganglion neurons (SGNs) are crucial for transferring auditory signals from cochlear sensory hair cells to the brainstem. However, SGNs are usually damaged in sensorineural hearing loss. Embryonic stem cells (ESCs) have been used to regenerate SGNs, but it is obscure whether ESC-derived neurons can fully resemble SGN subtype features. This study aimed to understand the effect of neurotrophins on the generation of SGN-like cells from ESCs and their subsequent subtype specification. This study utilized a stepwise neuronal generation approach to direct DsRed ESCs toward neural progenitors and eventually SGN-like cells. The derived SGN-like cells expressed multiple neuronal markers, including Tuj1, Map2, and NeuN, indicating maturity. Neurotrophins, including brain-derived neurotrophic factor, neutrotrophin-3, and nerve growth factor, seemed to regulate the generation of mature neurons from ESCs. In addition, derived neuron-like cells expressed the otic protein marker Gata3 and glutamatergic marker VGluT1, suggesting that they are SGN-like the glutamatergic cells. Significantly more SGN subtype marker-positive cells, including Pou4f1, calbindin, and calretinin-positive cells, were observed in the neurotrophin treatment groups. Overall, this study indicates the potential of SGN subtype generation from ESCs, which could be significant for cochlear implant therapy or stem cell-based replacement studies.
Introduction
Spiral ganglion neurons (SGNs) are the primary cochlear afferent neurons in the auditory system, which transfer auditory signals from cochlear sensory hair cells (HCs) to the cochlear nucleus at the brainstem. 1,2 The degeneration of SGNs usually causes sensorineural hearing loss. 3 This injury is typically irreversible since the mammalian cochlear sensorineural tissues, including HC and SGNs, lack the regeneration ability. In clinics, cochlear implants have been used to treat sensorineural hearing loss. However, a healthy amount of SGNs are necessary for the successful application of cochlear implant therapy. Therefore, studies on SGN regeneration also have clinical significance.
Afferent SGNs are glutamatergic neurons and are classified into type I and type II neurons. 4 –8 Type I SGNs constitute 95% of the SGN population, while the remaining 5% are type II SGNs. Type I SGNs are the primary cells that transmit complex sound information to the brain and approximately 10–20 of Type I SGNs innervate a single inner HC mono-synaptically. 9 Type I SGNs are subdivided into Ia, Ib, and Ic subtypes, and calretinin, calbindin, and Pou4f1 are typically expressed in type Ia, Ib, and Ic subtype neurons, respectively. 4,10,11 A previous report suggested that Type II SGNs appear to be involved in both damage detection and pain transmission. 12 Because SGNs are critical for hearing and SGNs cannot regenerate automatically following damage, many studies have been conducted to improve SGN regeneration. Embryonic stem cells (ESCs) have been used to generate SGN-like cells in vitro for stem cell-based replacement. 13 In our and other groups’ previous reports, ESCs have been induced into neuron-like cells expressing several neuronal and glutamatergic proteins. 13,14 However, it remains unclear whether ESC-derived neurons demonstrated SGN subtype features.
Although neurotrophins have been reported to influence auditory neuron survival and neurogenesis during development, 15 –17 their effect on stem cell-derived SGN subtype generation has not been investigated. In this study, we will continue our previous study and apply neurotrophins, including nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), and neurotrophin-3 (NT3), in the ESC induction to evaluate their roles in the generation of SGN-like cells and subsequent SGN subtypes. In addition, since SGNs are the target neurons for cochlear implants, a comprehensive understanding of SGN subtype regeneration is clinically important.
Materials and Methods
Neural differentiation of DsRed ESCs
DsRed ESCs were obtained from the Nagy lab 18 and were maintained in the ESC culture medium (82% DMEM/F12 GlutaMAX, 15% knockout fetal bovine serum [FBS], 1% nonessential amino acids, 1% Na-pyruvate, 1% penicillin/streptomycin, 55 nM 2-mercaptoethanol; all from Invitrogen), and leukemia inhibitory factor (LIF; 1,000 unit/mL, Millipore) in a 5% CO2 and 37°C incubator using our previously published methods. 14 In this study, the culture medium was replenished every 2–3 days to ensure optimal growth conditions for all cultures. Cell cultures were observed daily using phase contrast microscopy (Leica) and were passaged at approximately 70% confluence. DsRed ESCs were dissociated with TryplE (Invitrogen) followed by culturing in LIF-free ESC culture medium overnight. All-trans retinoic acid (Sigma, 10−7M) was added to the culture medium for 6–7 days, and cells were passaged using TryplE, followed by either monolayer or suspension cultures. The monolayer group was cultured in the adherent culture condition with the differentiation medium (49% Neurobasal, 49% DMEM/F12 GlutaMAX (Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12), 1% FBS, 55 nM 2-mercaptoethanol with NGF 20 ng/mL) for 14 days, followed by fixing and immunofluorescence. The suspension group was cultured in suspension medium (48% DMEM/F12 GlutaMAX, 48% Neurobasal, 1% N2, 2% B27, 55 nM 2-mercaptoethanol, 20 ng/mL EGF (Epidermal growth factor) (all from Invitrogen) and 20 ng/mL FGF-2 (Fibroblast growth factor 2; R&D) for 12–14 days for neurosphere formation. Then the floating spheres were collected, mechanically, dissociated and seeded. They were cultured in the differentiation culture medium (49% Neurobasal, 49% DMEM/F12 GlutaMAX, 1% FBS, 55 nM 2-Mercaptoethanol for control or with NGF 20 ng/mL, BDNF 20 ng/mL or NT-3 20 ng/mL for the neurotrophin groups) for 12–14 days, followed by fixing and immunofluorescence.
Immunofluorescence
After fixation with 4% paraformaldehyde for 10 min at room temperature and rinsed with PBS (Phosphate buffered saline), cell samples were treated with PBS containing 5% donkey serum (Jackson ImmunoResearch) and 0.2% Triton X-100 (Sigma) for 30 min at room temperature. Samples were incubated in primary antibodies at 4°C overnight, followed by corresponding secondary antibodies incubation at room temperature for 2 h. Primary antibodies were antibodies specific for Nestin (Developmental Studies Hybridoma Bank, 1:200), Sox2 (Abcam, 1:200), Tuj1 (β-tubulin type III, Sigma, 1:1000), Microtubule-associated protein 2 (Map2, Sigma, 1:1000), Neuronal nuclear protein (NeuN, Sigma, 1:200), Pou4f1 (Sigma, 1:1000), calbindin (Cell signaling, 1:1000), calretinin (Sigma, 1:200), Peripherin (Sigma, 1:200), Tyrosine Hydroxylase (TH, Sigma, 1:500), Gata3 (Sigma, 1:1000), and vesicular glutamate transporter 1 (VGluT1, Sigma, 1:200). Secondary antibodies included Alexa Fluor 488, 405, and 647 conjugated donkey anti-chicken, anti-goat, anti-mouse, anti-rat, and anti-rabbit antibodies (1:500; Jackson Immunoresearch). Samples were observed and imaged by Leica epifluorescence and Leica SPE confocal microscopy.
Quantification study and statistical analysis
Samples for quantitative and statistical analyses were collected from at least three independent experiments, and each group had six samples (n = 6 per group). Cells were analyzed and counted by the ImageJ software (NIH; National Institutes of Health) using the cell counter plugin module. The following standards were used to observe and evaluate neuron-like cells. Tuj1 positive cells with typical neuronal morphology, such as large and round cells with centrally located nucleus. In the quantitative study, a normalized method was used, in which the number of cells in the control group was normalized as 1, and the cell numbers in the other groups were normalized against the number of the control group.
All analyses were performed using GraphPad Prism 8 (GraphPad Software, San Diego, CA, USA) and Microsoft Excel (Microsoft, Redmond, WA, USA). Student’s t-test or one-way analysis of variance (ANOVA) was applied for analysis. Results were considered statistically significant at a level of P < 0.05.
Results
Induction of DsRed ESCs into neuron-like cells
Two different approaches were tested to generate neuronal-like cells with neurite projections. In the first monolayer approach, DsRed ESC colonies were treated with retinoic acid for 6–7 days to guide them toward neural progenitor fate. 19,20 These retinoic acid-treated cells were cultured in the monolayer for 14 days with NGF for neural differentiation (Fig. 1A). In the second approach, DsRed ESC colonies were treated with retinoic acid for 6–7 days as above and then suspended in the suspension medium to form spherical cell clusters (neurospheres) and maintained in suspension for 12–14 days (Fig. 1A). We found that neurospheres expressed general neural stem cell proteins, such as Sox2 and Nestin, when they were cultured for 12 days (Fig. 1B). These neurospheres were then mechanically dissociated and cultured in the medium containing NGF for 14 days to guide neuronal differentiation and maturation. It is observed that DsRed ESC-derived neuron-like cells expressed neuronal markers Tuj1 and Map2 (Figs. 1C and 1D). Neurites extending from the soma can be observed in the neurosphere group whereas neurites were hardly seen in the monolayer approach (Fig. 1D). To mimic the characteristics of typical SGN cells with pseudo-bipolar neurite projections, the neurosphere approach was used to continue the differentiation of DsRed ESC into neuron-like cells in the following experiment.
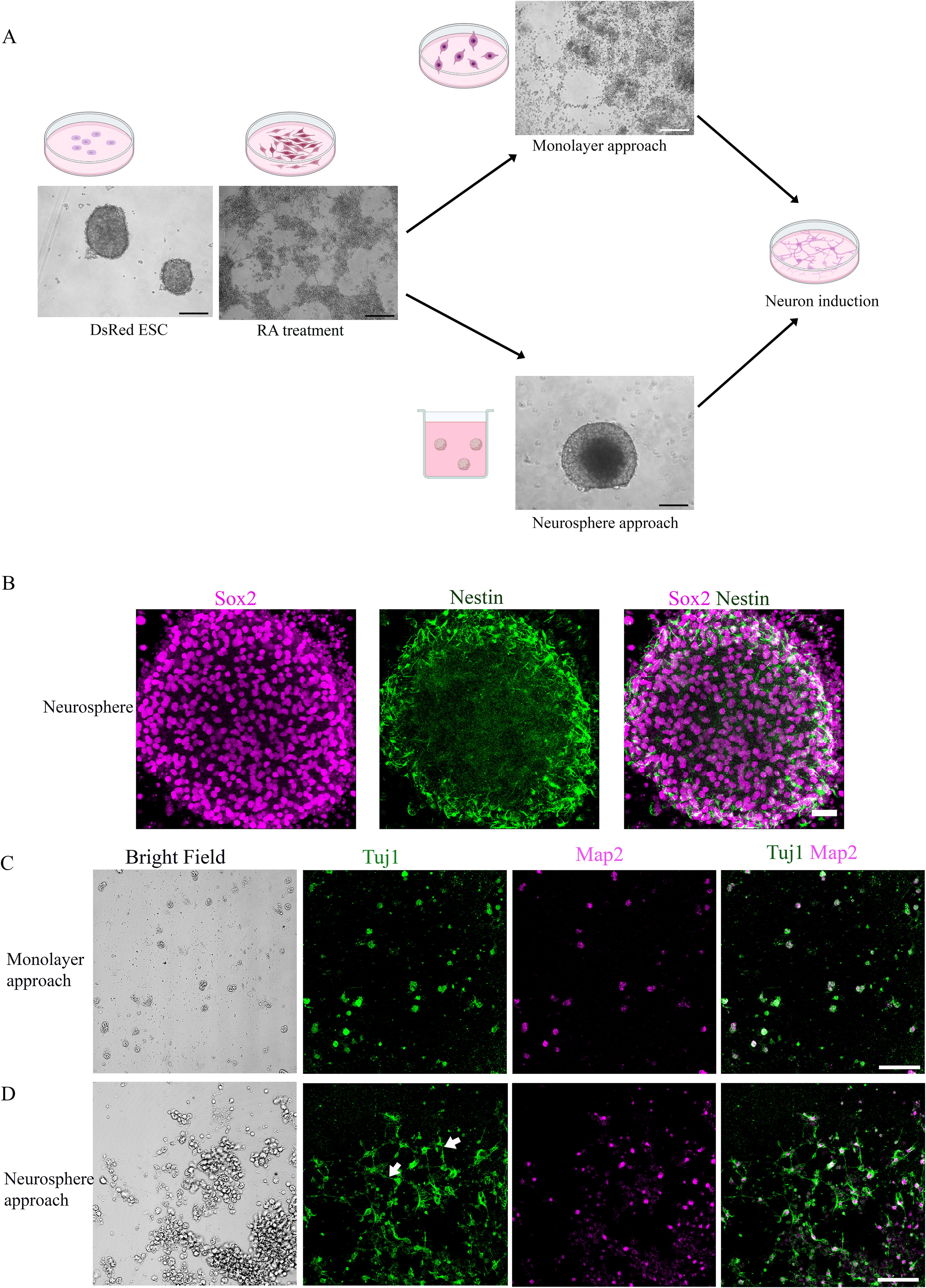
Developing a refined sequential method for generating DsRed-derived neuron-like cells with extended neuronal projections.
Neurotrophin regulates neuron-like cell generation from ESCs
Several previous reports suggested that neurotrophin plays a critical role in the development of SGNs, 21 –23 but it is unclear whether neurotrophin can regulate neuronal differentiation from ESCs. In this study, we examined whether BDNF, NT-3, and NGF treatment would affect neuron-like cell differentiation.
BDNF, NT-3, and NGF were added to the differentiation medium in the treatment groups, whereas the control group only had the differentiation medium without neurotrophins. Cells were maintained for the same period for 12–14 days in all groups and then fixed for immunostaining. Neuron-like cells expressed immature neural markers Tuj1 and mature neuronal markers, including Map2 and NeuN, in the control and treatment groups (Fig. 2A).
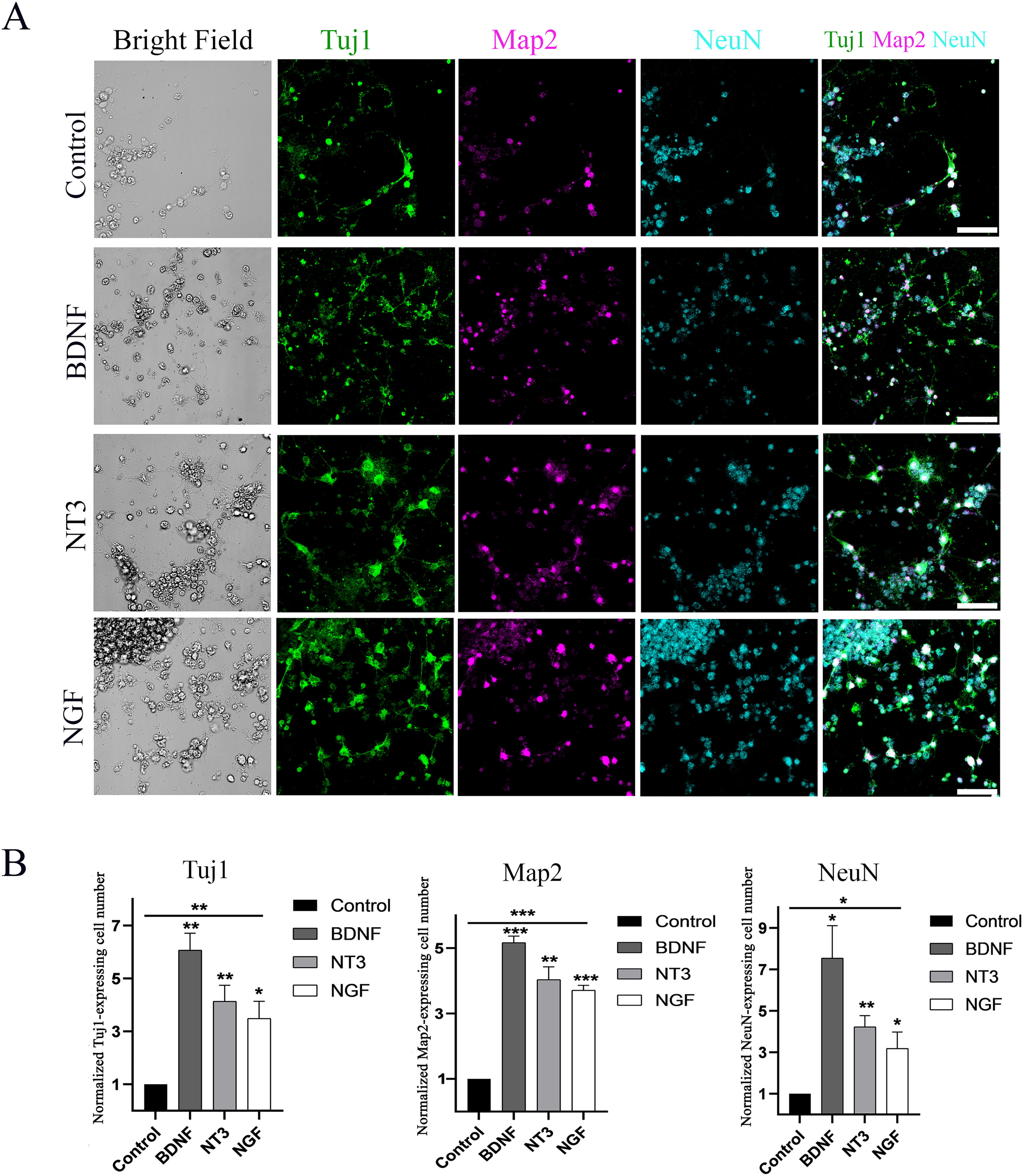
DsRed ESCs derived neuron-like cells express mature neuronal markers.
The quantitative study showed that the normalized Tuj1 positive cell numbers in the control, BDNF, NT-3, and NGF were 1, 6.07 ± 0.63, 4.13 ± 1.05, and 3.49 ± 1.11, respectively (mean ± SEM, Fig. 2B). The neurotrophin treatment groups showed 3 to 6-fold more Tuj1 cells than that of the control group. ANOVA revealed that BDNF, NT-3, and NGF groups had a significantly increased number of Tuj1 positive cells (the P values were 0.001, 0.007, and 0.02). These data suggest that neurotrophins may be able to regulate the generation of Tuj1-positive cells from ESCs.
The normalized Map2 positive cell numbers in the control, BDNF, NT-3, and NGF were 1, 5.16 ± 0.34, 4.03 ± 0.67, and 3.7 ± 0.26, respectively (mean ±SEM, Fig. 2B). The neurotrophin treatment groups showed 3 to 5-fold more Map2 cells than that of the control group. ANOVA revealed that BDNF, NT-3, and NGF groups had significantly large number of Map2 positive cells (the P values were <0.0001, 0.002, and <0.0001). These data suggest that neurotrophins may regulate the generation of Map2-positive cells from ESCs.
The NeuN positive cell numbers in the control, BDNF, NT-3, and NGF were 1, 7.54 ± 2.7, 4.22 ± 0.94, and 3.19 ± 1.36, respectively (mean ±SEM, Fig. 2B). The neurotrophin treatment groups showed 3 to 7-fold more NeuN cells than that of the control group. ANOVA analysis revealed that the BDNF, NT-3, and NGF groups exhibited a significantly increased number of NeuN-positive cells (the P values were 0.02, 0.004, and 0.05). These data suggest that neurotrophins may regulate the generation of NeuN-positive cells from ESCs.
Taken together, the above statistical analysis reveals that BDNF, NT-3, and NGF groups had a significantly increased number of Tuj1, Map2, and NeuN positive cells compared with the control group (P < 0.05, one-way ANOVA, n = 6 samples from 3 independent experiments, Fig. 2B). These data suggest that neurotrophin may play a role in the neuronal differentiation from ESCs.
Induced neuron-like cells express the otic protein GATA3 and the glutamatergic marker VGluT1
Since mature mouse SGNs are glutamatergic neurons, we evaluated the yield of glutamatergic neurons by analyzing the expression of vesicular glutamate transporter 1 (VGluT1) and Gata binding protein 3 (Gata3), a gene regulating auditory system development and function. 24 ESCs were induced into neuron-like cells as above, followed by fixation and immunostaining. It is observed that DsRed ESC-derived neuron-like cells expressed the glutamatergic neural marker VGluT1 (Fig. 3). In addition, DsRed ESC-derived neuron-like cells also expressed Gata3. The expression of VGluT1 and Gata3 was observed in the control and all neurotrophin groups, including BDNF, NT-3, and NGF treatment groups.
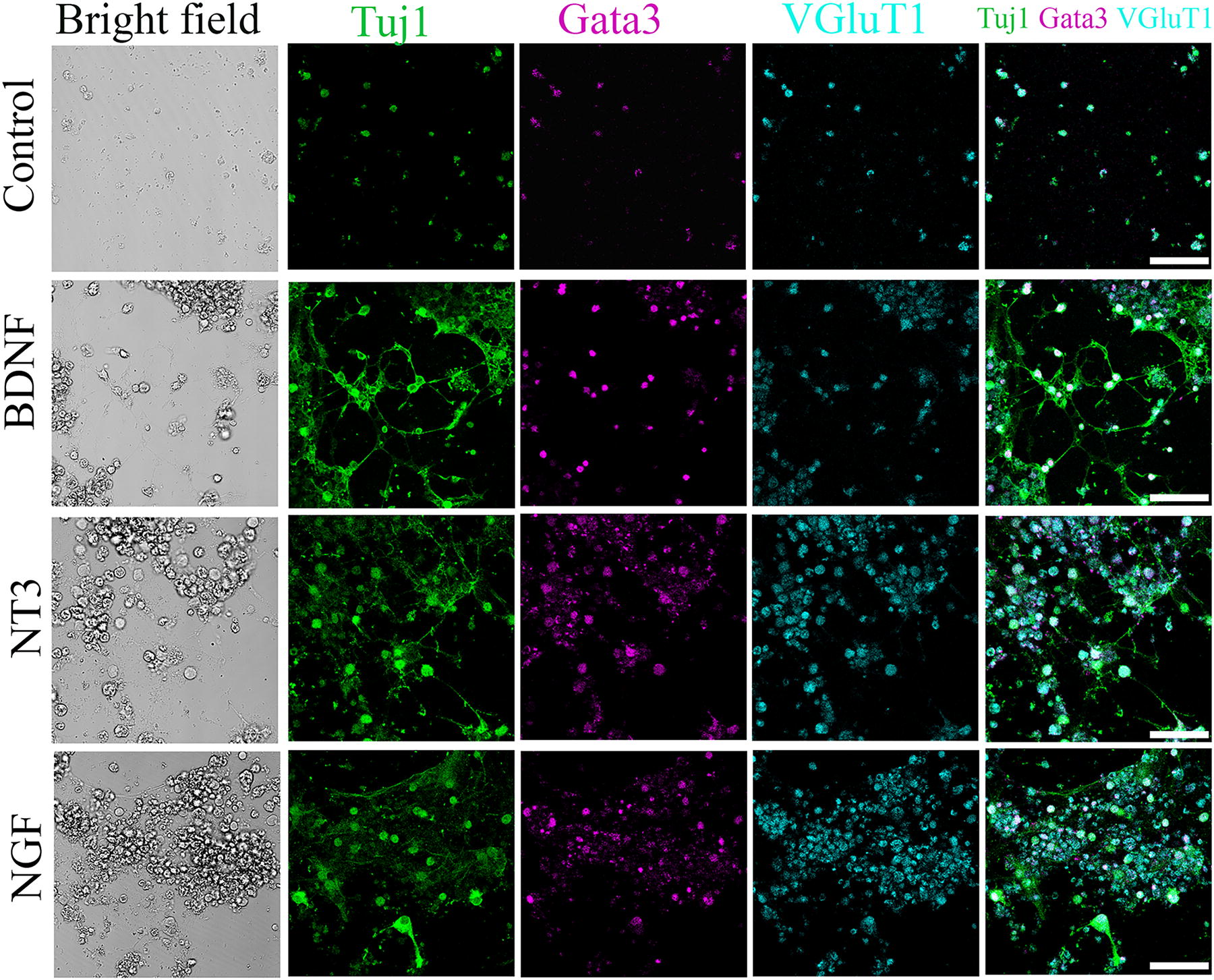
Induced neuron-like cells express key proteins characteristic of SGNs. Neuron-like cells express glutamatergic protein VGluT1 and otic protein Gata3 in the control, NGF, BDNF, and NT3 groups. Scale bar: 50 μm. SGN, spiral ganglion neuron; VGluT1, vesicular glutamate transporter-1.
SGN-like cells express the subtype I and subtype II markers
To evaluate the subtype specification of SGN-like cells, immunofluorescence with antibodies specific for SGN subtype proteins was applied to ESC-derived neuron-like cells. In addition, to determine whether neurotrophins affect SGN subtype specification, BDNF, NT3, and NGF were added to the differentiation medium.
It was found that neuronal-like cells in the control, BDNF, NT3, and NGF groups expressed Pou4f1 (type 1c), calbindin (type 1b), and calretinin (type 1a; Fig. 4A). In the NGF group, SGN-like cells also expressed type II proteins, including tyrosine hydroxylase (TH) and peripherin, suggesting the generation of type II SGNs (Fig. 4B). To examine whether neurotrophins were able to stimulate the generation of SGN subtypes, the number of Pou4f1, calbindin, and calretinin in control, BDNF, NT3, and NGF groups was counted.
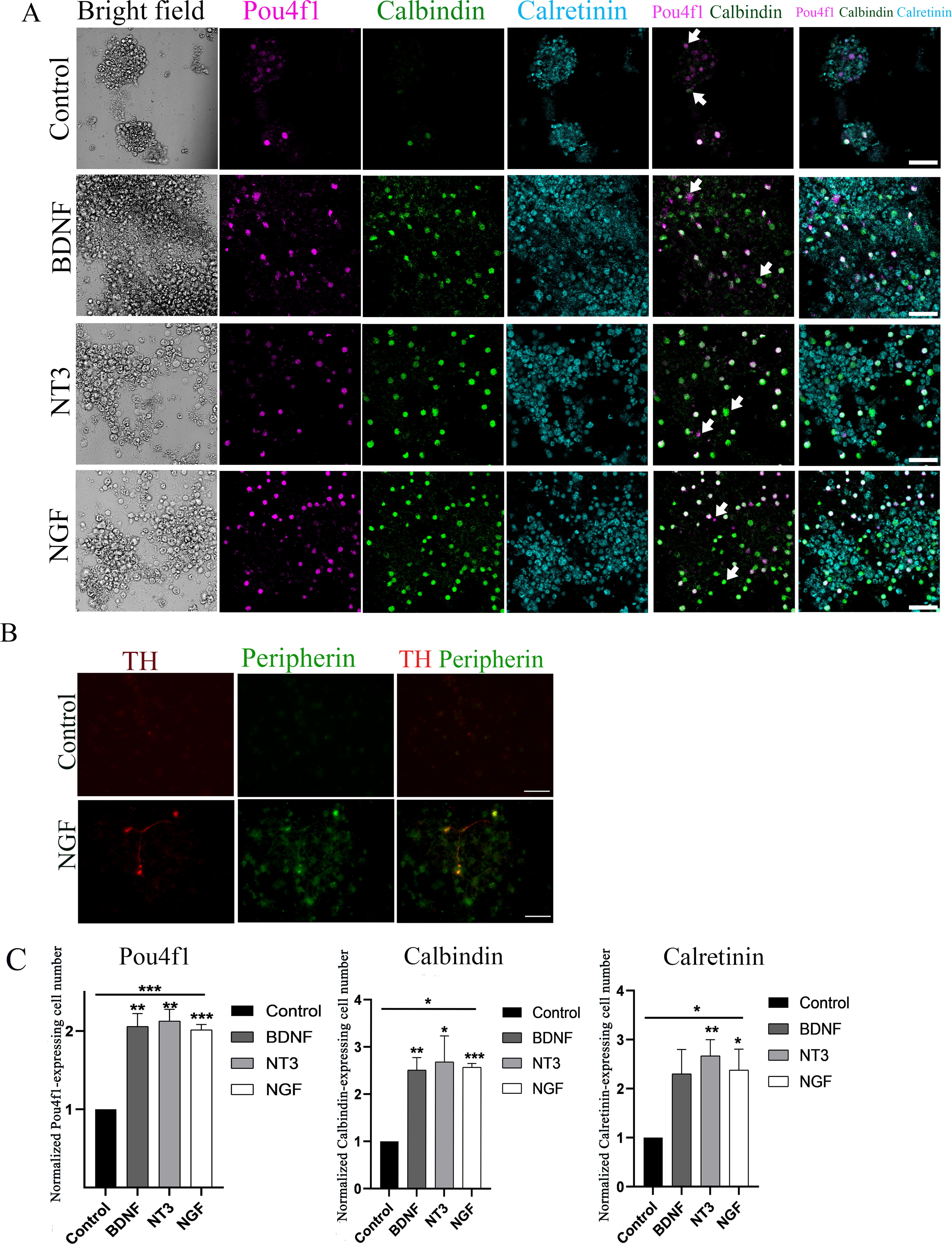
Neuron-like cells express markers for SGN subtypes.
The quantitative study showed that the normalized Pou4f1 positive cell numbers in the control, BDNF, NT-3, and NGF were 1, 2.05 ± 0.28, 2.12 ± 0.25, and 2.01 ± 0.12, respectively (mean ± SEM, Fig. 4C). The neurotrophin treatment groups showed approximately 2-fold more Pou4f1 positive cells than that of the control group. ANOVA revealed that BDNF, NT-3, and NGF groups had significantly increased numbers of Pou4f1 positive cells (the P values were 0.003, 0.002, and 0.0001). These data suggest that neurotrophins may be able to regulate the generation of Pou4f1-positive cells from ESCs.
The normalized calbindin positive cell number in the control, BDNF, NT-3, and NGF were 1, 2.5 ± 0.45, 2.68 ± 0.95, and 2.57 ± 0.13, respectively (mean ±SEM, Fig. 4C). The neurotrophin treatment groups showed approximately 2.5-fold more calbindin cells than that of the control group. ANOVA revealed that BDNF, NT-3, and NGF groups exhibited a significantly large number of calbindin positive cells (the P values were 0.005, 0.04, and <0.0001). These data suggest that neurotrophins may regulate the generation of calbindin-positive cells from ESCs.
The normalized calretinin-positive cell numbers in the control, BDNF, NT-3, and NGF were 1, 2.3 ± 0.86, 2.66 ± 0.57, and 2.38 ± 0.73, respectively (mean ±SEM, Fig. 4C). The neurotrophin treatment groups showed about 2-fold more calretinin cells than that of the control group. ANOVA analysis demonstrated that the BDNF, NT-3, and NGF groups exhibited a significantly higher number of calretinin-positive cells (the P values were 0.061, 0.008, and 0.032). These data suggest that neurotrophins may regulate the generation of calretinin-positive cells from ESCs.
Taken together, the above statistical analysis reveals that compared to the control group, significantly more Pou4f1, calbindin, and calretinin positive cells were observed in BDNF, NT3, and NGF treatment groups (P < 0.05, one-way ANOVA, n = 6 samples from 3 independent experiments), suggesting that neurotrophin treatment may play a role in subtype specification.
Discussion
Based on physiological, anatomical, and transcriptional characteristics, SGNs comprise several subtypes, 6 –8 which have been confirmed by single-cell RNA sequencing studies. 4,10,25 A recent report further documented that the specification of SGN subtypes occurs prior to birth. 8 It is important to note that all these findings stem from normal developmental studies. In response to the need for SGN replacement, a stem cell-based approach has been proposed, and ESCs have been applied to generate SGN-like neurons. 13,14,26,27 For an effective stem cell-based SGN replacement strategy, it is crucial to establish whether stem cells can differentiate into specific SGN subtypes. In this report, we employed DsRed ESCs to tackle this challenge. To direct DsRed ESCs differentiation toward neural progenitors and ultimately SGN-like cells, a sequential neuronal generation approach was utilized in this study, including the generation of neurospheres and induction of SGN-like cells. ESC-induced cells expressed multiple neuronal markers, such as Tuj1, Map2, and NeuN, indicating that they are neuron-like cells. Moreover, ESC-induced neuron-like cells expressed the otic protein marker Gata3 and the glutamatergic marker VGluT1, suggesting that the generated neurons are SGN-like glutamatergic cells. In addition, DsRed ESC-induced SGN-like cells expressed SGN subtype markers, Pou4f1, calbindin, and calretinin, which was enhanced by the application of neurotrophins, including NGF, BDNF, and NT3.
ESCs have been used to generate neuron-like cells, but it is still obscure whether ESC-derived neurons demonstrate mature inner ear neuronal features. In this study, in addition to Tuj1, a marker of immature neurons, mature neuronal markers, such as Map2 and NeuN, 28,29 were also used for the characterization of ESC-derived neuron-like cells. Our results showed that DsRed ESC-derived neuron-like cells expressed mature neuronal markers, including Map2 and NeuN, suggesting the generation of neuron-like cells expressing mature neuronal proteins (Fig. 2). Since adult mouse SGNs are glutamatergic neurons, it is fundamental to determine whether induced neuron-like cells express the glutamatergic protein VGluT1. In addition, SGNs are derived from the proneurosensory tissue, which are suggested in development by the expression of transcription factor networks of Gata3. Accordingly, we evaluated the expressions of VGluT1 and Gata3 in this study. It was observed that neuron-like cells expressed glutamatergic marker VGluT1 (Fig. 3), suggesting that DsRed ESC-derived neuron-like cells may be the glutamatergic neurons. It was also found that DsRed ESC-derived SGN-like cells expressed Gata3, a zinc finger transcription factor expressed in embryonic primary auditory neurons. 30 These data show that ESC-derived neurons express mature, glutamatergic, and otic proteins, which suggests that mature inner ear neurons may be able to be generated from ESCs in vitro.
Neurotrophic factors, such as BDNF and NT3, are known to play a critical role in SGN development 9,21 –23 and have also been shown to prevent SGN degeneration 31,32 and enhance the functional responsiveness of the auditory system. 33 However, it is unclear whether neurotrophins are able to stimulate the induction of SGN-like cells from ESCs in vitro. Further, it remains obscure whether the in vitro specification of SGN subtypes is regulated by neurotrophins. In the current study, compared with the control group, a higher number of neuron-like cells expressing immature and mature neuronal markers, including Tuj1, Map2, and NeuN, were observed in the neurotrophin groups. In addition, we found that ESC-derived SGN-like cells expressed SGN subtype markers, including Pou4f1, calbindin, and calretinin (Fig. 4). Moreover, the normalized number of calretinin, calbindin, and Pou4f1 positive neuron-like cells was significantly increased in the neurotrophin groups (P < 0.05, ANOVA). These data suggest that neurotrophins may promote the differentiation of ESCs into neuron-like and SGN-subtype cells. In addition, the data indicate that neurotrophins may enhance the survival of these newly generated neuron-like and SGN-subtype cells. However, since these cells are continuously produced from ESC-derived NSCs, it remains technically challenging to distinguish between the effects of neurotrophins on stimulating differentiation and promoting the survival of these newly generated cells. The primary aim of this study is to establish proof-of-concept that SGN subtype neuron-like cells can be generated from ESCs. In future studies, we plan to investigate in greater detail whether neurotrophins primarily stimulate the generation of SGN-like cells, maintain their survival, or contribute to both processes.
In summary, stem cells have been explored for SGN replacement; however, previous studies have not yet determined whether specific SGN subtypes can be derived from ESCs. Our study represents a significant initial step in identifying the potential of ESCs to differentiate into neuron-like cells that express markers specific to SGN subtypes. These ESC-derived subtype neuron-like cells could be valuable in SGN replacement research and may ultimately enhance the efficacy of cochlear implants. It is important to note that our current study primarily focuses on the protein expression of these new subtype neurons. Consequently, further detailed studies will be necessary to fully characterize these ESC-derived subtype neuron-like cells in future research.
Footnotes
Author Disclosure Statement
The authors declare that they have no competing financial interests related to the research presented in this article. All research was conducted independently, and no funding sources influenced the study design, data analysis, or interpretation of results.
Funding Information
This study was supported in part by Merit Review Award # I01 RX002100 from the United States (U.S.) Department of Veterans Affairs Rehabilitation Research and Development Service. The contents do not represent the views of the U.S. Department of Veterans Affairs or the United States Government. Anti-Nestin antibodies were obtained from Developmental Studies Hybridoma Bank (DSHB).
Supplementary Material
Supplementary Figure S1