
Abstract
Therapies aimed at manipulating microvasculature require the ability to generate both blood and lymphatic vessels. Adipose-derived stromal vascular fraction (SVF), consisting of endothelial cells, progenitor cells, pericytes, smooth muscle cells, fibroblasts, and immune cells, has emerged as a heterogeneous cell composition able to promote blood vessel formation and growth, but whether SVF forms lymphatic vessels remains unknown. The objective of this study was to evaluate whether SVF can form lymphatic vessels. SVF was isolated from C57BL/6 mouse inguinal adipose tissue, characterized for prevalence of blood (PECAM+) and lymphatic (Prox1+, Podoplanin+, LYVE-1+) endothelial cells and cultured with avascular mouse mesentery tissues for up to 9 days. The presence of lymphatic endothelial cells in SVF is supported by the percentages of PECAM+ cells that are also positive for lymphatic markers. By day 1 after SVF seeding, cells established PECAM+ segments, and by day 3 cell clusters with segment extensions were observed. At later time points, segments established network of blood vessels. In parallel, a subset of structures positive for lymphatic marker labeling and characterized by a rounded shape (termed “blebs”) connected with nearby SVF-derived blood vessel and were changing shape over time. Our findings provoke a new research area focused on the ability for SVF to form lymphatic vessels.
Introduction
Regenerative medicine therapies aimed to promote microvasculature development require the ability to generate both blood and lymphatic vessels. 1 Adipose-derived stromal vascular fraction (SVF), consisting of endothelial cells, endothelial and mesenchymal progenitor cells, pericytes, smooth muscle cells, fibroblasts, and immune cells, represents an attractive cell composition 2 –4 for such therapies. 5 –8 Clinical trials have used SVF therapy to treat ischemic disease, 9,10 diabetic foot ulcers, 11 –13 skin flap transplantation, 14 scar healing, 15,16 osteoarthritis, 17,18 chronic wounds, 19 –21 and pulmonary fibrosis. 22,23 To understand the mechanism of SVF’s ability to improve tissue function and more specifically promote the growth of functional microvascular networks, it will be essential to define the fate and distribution of the cells of SVF in the tissue after delivery, their effects on host tissues, and contribution to microvascular remodeling.
The emerging interest in SVF and other adipose tissue-derived cell mixtures 24 –26 is challenged by the basic science questions and the lack of studies that characterize SVF capabilities. Consider the publications related to microvascular remodeling and the growth of blood and lymphatic vessels. A PubMed search for “stromal vascular fraction + angiogenesis” results in less than 250 articles since the discovery of SVF in 2001. 27 In vitro studies have established that cells of SVF undergo self-reassembly into blood microvessels. 28 –31 Furthermore, implanted SVF form patent microvessels. 32 –35 While it has been proven that SVF promotes formation of new vessels, the contribution of SVF on angiogenesis of the host microvascular network is not clear. Recent results from our group offered temporal and spatial insights related to the enabled evaluation of SVF effects on angiogenesis of host microvascular networks versus SVF neovascularization, 29 yet our understanding of how SVF-derived vessels integrate with host networks and whether SVF-derived vessels are functional remains unclear.
We also do not know about the ability of SVF to influence lymphatic vessel growth as well as prevalence of lymphatic endothelial cells in SVF. A PubMed search for “stromal vascular fraction lymphatic” resulted in 20 articles, and a search for “stromal vascular fraction lymphangiogenesis” resulted in 1 article. To explore this knowledge gap, the objective of this study was to evaluate whether SVF can form lymphatic vessels. SVF seeded ex vivo on avascular mouse mesenteric tissues. We decided to use mouse mesenteric tissues in part because the thin connective tissues in mouse, termed “mesenteric windows,” are avascular, 36 and any vessel structures observed will be considered SVF derived. Our results suggest that SVF contains lymphatic endothelial cells and forms lymphatic vessel-like structures.
Materials and Methods
Animal use
All animal experiments were approved by the University of Florida’s or the Wake Forest University’s Institutional Animal Care and Use Committee. Adult female C57BL/6 mice (Jackson Laboratory or Charles River Labs; 18–25 g) or adult female C57BL/6-Tg(CAG-EGFP)131Osb/LeySopJ mice (C57BL6-GFP mice; Jackson Laboratory; 18–25 g) were euthanized via CO2 suffocation and cervical dislocation or via cervical dislocation post 5% isoflurane inhalation for approximately 30 min. Adult male Wistar rats (Inotiv; 350–395 g) were anesthetized by intraperitoneal injection of ketamine (80 mg/kg body weight) and xylazine (8 mg/kg body weight) and used for the in vivo SVF transplantation study. Immediately after mesentery and/or fat harvest, rats were euthanized by intracardiac injection of single dose of 0.2-mL Beuthanasia.
Mouse mesentery tissue harvest
Mouse mesenteric tissues were harvested and cultured according to a previously established protocol. 36 Rat mesenteric tissues were harvested and cultured according to a previously established protocol. 29 To harvest mesenteric tissues, the abdominal region was shaved and depilated, treated with 70% isopropyl alcohol and iodine, followed by incision along the linea alba. The mesentery was exteriorized onto a plastic stage using cotton tip applicators to first remove the cecum and subsequently the ileum and jejunum. Mesenteric tissues were excised, rinsed in sterile 0.9% NaCl, and immersed in minimum essential media (MEM; Gibco, Catalog #11095080) containing 1% penicillin–streptomycin (PenStrep; Gibco, Catalog #15140122) at 37°C.
Adipose tissue harvest
A scalpel was used to make a Y-shaped incision into the skin from the bottom of the of the linea alba to each of the inner thighs along the midline, stopping above the knee. The skin was removed from the subcutaneous adipose and muscle layers using the scalpel and microscissors. Hemostats were used to hold the skin back as the separation advanced to the coxal region. The center skin flap of the pelvic region was removed in a similar fashion. The inguinal fat pad was removed in a continuous strip from one hip and leg to the other and placed into sterile DPBS at 37°C. Then, fat was diced into fine pieces and weighed. This adipose harvesting method was used for both mice and rats.
SVF isolation
Adipose tissue was incubated in 0.15% collagenase type 1 (Thermo Fisher, Catalog #17100017) in DPBS at 37°C on a shaker set to 150 RPM for 30 min at a ratio of 2 mL per 1 g of tissue. Collagenase solution was then pushed through a 0.22-µm filter (Thermo Fisher, Catalog #9740113) into the 50-mL conical tube containing the diced fat. In the event that the fat weighed less than 2.5 g, 5 mL was used to ensure enough volume was in the conical tube containing the diced adipose. The collagenase–adipose mixture was then transferred to a shaker set to 150 RPM and digested for 30 min at 37°C. After digestion, the mixture was centrifuged at 600g for 10 min to pellet the cells. The top layer of undigested fat was transferred to a tube containing fresh collagenase and transferred back to the incubator for a second digestion. Cells pelleted from both digestions were resuspended in 10% fetal bovine serum (FBS)/DPBS, filtered through a 200-µm cell strainer (pluriSelect, Catalog #435020003), pelleted by centrifugation at 200g for 5 min, and resuspended in 5% FBS/DPBS. Cell suspension was filtered through a 70-µm filter (Alkali Scientific, Catalog #MT4070) and centrifuged at 200g for 5 min. The media was removed, and cells, representing SVF and blood-derived non-SVF cells, were resuspended in 10% FBS/DPBS and counted with a NucleoCounter® NC-200™ (ChemoMetec).
Flow cytometry characterization of lymphatic endothelial cell presence
SVF cells were stained with fixable viability dye for 30 min, washed twice with DPBS, incubated in blocking serum for 15 min, and then with PerCP-Cy™5.5 anti-mouse CD31 IgG (PECAM; BD Biosciences, Catalog #562861), APC antimouse Podoplanin IgG (BioLegend, Catalog #127410), and PE anti-mouse LYVE-1 (R&D System, Catalog #FAB2125P) for 30 min. Then cells were washed to remove IgG, incubated with BD Cytofix/Cytoperm Kit (BD, Catalog #554714) for 20 min, and incubated with FITC Prox1 IgG (Novus Biologicals, Catalog #NBP1-30045F) for 30 min. Data acquisition was done on a BD FACS Canto II Analyzer with 3-laser, 8-color configuration (Wake Forest Medical Center Flow Core Facility) and analyzed by FCS Express software. Unstained cells served as negative control, whereas cells stained with single color for each conjugated were used for establishing compensation. Fixable viability dye allowed to exclude dead cells and TER-119 (Thermo Fisher, Catalog #A18452) to exclude erythrocytes.
SVF transplantation and culture on mouse mesentery
Isolated SVF cells were suspended in 10% FBS/MEM at 107 cells/mL. Murine mesentery tissues were spread onto a Isopore membrane filter with pore size 5 µm (Millipore Sigma, Catalog #TMTP04700) on cell culture insert (CellCrown insert; Millipore Sigma, Catalog #Z742381); 5 × 105 SVF cells in (20–40 µL) were plated on mesenteric window, followed by 25-min incubation to allow cells to attach to the tissues. Each CellCrown insert was then inverted and pressed into a well of a 6-well plate so that the tissue was pressed between the bottom of the well and the filter membrane and 3 mL of 10% FBS/MEM was added to the wells on top of the filter. Tissues were cultured in standard incubation conditions (5% CO2, 37°C) for 2, 3, 5, 7, and 9 days.
In vivo SVF transplantation
Male Wistar rats (370–390 g) were anesthetized by intraperitoneal injection of ketamine (80 mg/kg body weight) and xylazine (8 mg/kg body weight). The abdominal surgical field was shaved and treated with 70% isopropanol and iodine. Analgesic injections of Ethiqa XR (0.65 mg/kg body weight) and Bupivacaine (2 mg/kg body weight) were administered subcutaneously on the dorsal side and near the incision site, respectively. After an incision was made on the linea alba, a random section of mesentery was exteriorized using cotton tip applicators and placed onto a plastic stage. Five to seven sequential tissues were selected, and 7–0 monofilament polyamide sutures (Ethicon, Catalog #1696G) were used to mark the first and last tissue; 5 × 105 rat SVF cells were deposited onto each tissue SVF cells and allowed to adhere to the tissues for 20 min, followed by returning exposed mesentery to the peritoneal cavity. The muscle layer was sutured back together using 5–0 monofilament polydioxanone sutures (Ethicon, Catalog #1995G) and the skin was closed using 3–0 monofilament polydioxanone sutures (Ethicon, Catalog #1993G). Rats were then given Meloxicam (5 mg/kg) and 5.0 mL of saline subcutaneously. Meloxicam was administered every 24 h for 72 h, and Ethiqa was administered every 72 h after initial dose. In Sham group, saline alone was applied onto the mesenteric tissues.
SVF fate tracking
To support the SVF-derived origin of the observed lymphatic endothelial cell structures, additional experiments were conducted to either track SVF-derived cells or determine if structures formed in host tissues without the presence of SVF.
Mouse SVF transplanted on rat mesentery
Wistar rat mesentery tissues were spread onto a Isopore membrane filter with pore size 5 µm (Millipore Sigma, Catalog #TMTP04700) on cell culture insert (CellCrown insert; Millipore Sigma, Catalog #Z742381); 5 × 105 isolated C56BL6 mouse SVF cells in (20–40 µL) were plated on mesenteric window, followed by 25-min incubation to allow cells to attach to the tissues.
GFP mouse SVF transplanted on mouse mesentery
C57BL6 mouse mesentery tissues were spread onto a Isopore membrane filter with pore size 5 µm (Millipore Sigma, Catalog #TMTP04700) on cell culture insert (CellCrown insert; Millipore Sigma, Catalog #Z742381); 5 × 105 isolated C57BL6-GFP mouse SVF cells in (20–40 µL) were plated on C57BL6 mouse mesenteric window, followed by 25-min incubation to allow cells to attach to the tissues.
Mouse mesentery tissue cultured without SVF (Sham Control)
C57BL6 mouse mesentery tissues were spread onto a membrane filter with pore size 5 µm (Millipore Sigma, Catalog #TMTP04700) on cell culture insert (CellCrown insert; Millipore Sigma, Catalog #Z742381). Each CellCrown insert was then inverted and pressed into a well of a 6-well plate so that the tissue was pressed between the bottom of the well and the filter membrane and 3 mL of 10% FBS/MEM was added to the wells on top of the filter. Tissues were cultured in standard incubation conditions (5% CO2, 37°C) for 3 or 6 days.
Immunohistochemistry
Following incubation, adipose tissues around mesenteric windows were removed and tissues were mounted on glass slides. Tissues were fixed in methanol at −20°C for 30 min and rinsed in 0.1% saponin (Sigma-Aldrich, Catalog #84510)/PBS three times for 10 min.
Mouse ex vivo tissues
Tissues were incubated with biotinylated rat antimouse PECAM antibody (1:200; BD Biosciences, Catalog #553371) and DAPI (1:3000; Thermo Fisher Scientific, Catalog #D1306), with rabbit polyclonal anti-LYVE-1 antibody (1:100; AngioBio, Catalog #11034) or rabbit anti-Prox1 antibody (1:200; AngioBio, Catalog #11002P) for 1 h at room temperature. Then, tissues were incubated with CY2-conjugated Streptavidin (1:500; Jackson Immunoresearch, Catalog #016220084) or goat antirabbit CY3-conjugated IgG (1:100; Jackson Immunoresearch, Catalog #111585144) for 1 h at room temperature.
Rat in vivo tissues
Tissues were incubated 1 h with biotinylated mouse anti-rat PECAM IgG (1:200; BD Biosciences, Catalog #555026), with rabbit anti-LYVE-1 IgG (1:100; AngioBio, Catalog #11034) or anti-Prox1 IgG (1:200; AngioBio, Catalog #11111585002P) at room temperature, followed by 1-h incubation with CY2-conjugated Streptavidin and goat anti-rabbit CY3-conjugated IgG at room temperature.
Rat ex vivo tissues co-cultured with mouse SVF
Tissues were incubated with biotinylated rat antimouse PECAM antibody (1:200; BD Biosciences, Catalog #553371), with BD OptiBuild™ BUV 615 mouse antirat PECAM antibody (1:100; BD Biosciences, Catalog #751394) for 1 h at room temperature. Then, tissues were incubated with CY2-conjugated Streptavidin (1:500; Jackson Immunoresearch, Catalog #016220084) for 1 h at room temperature.
Antibodies were diluted in buffer comprising of PBS, 0.1% saponin, 2% bovine serum albumin (Jackson Immunoresearch, Catalog #001000162), and 5% normal goat serum (Jackson Immunoresearch, Catalog #005000121). After each antibody incubation, the tissues were washed in 0.1% saponin/PBS (three times for 10 min).
Images of stained mesentery tissues were captured using Nikon Eclipse Ti2 inverted microscope coupled with an Andor Zyla sCMOS camera and quantitatively analyzed with ImageJ (National Institutes of Health).
Quantification of SVF-derived lymphatic structures
A 4× montaged image of a tissue was opened in ImageJ. Per image the following measurements were made: tissue area, the number of PECAM+/LYVE-1+ lymphatic structures, the size of each PECAM+/LYVE-1+ lymphatic structure defined by the circumscribing area, normalized by tissue area, the number of lymphatic structures with mosaic labeling (all cells are PECAM+, but only partially LYVE-1+), the maximum length of each lymphatic structure, and the average width per structure. We also quantified the number and lengths of PECAM+/LYVE-1+ structures attached to at least a PECAM+/LYVE-1− endothelial segment. One data point is the average quantification for one tissue.
Statistical analysis
In the experiments of mouse mesentery tissues cultured with transplanted mouse SVF, analysis of variance and Tukey’s honest significant difference test were used for pairwise comparisons between different time-point groups with P value ≤0.05 considered to be statistically significant. Statistical analysis was performed using Prism (GraphPad Software). Values are presented as mean ± standard error of the mean.
Results
The prevalence of PECAM+ cells in SVF was 67.8 ± 2.2%, whereas prevalence of cells dual positive for PECAM and one of the lymphatic markers was PECAM+/Prox1+: 5.6 ± 2.2%, PECAM+/LYVE-1+: 1.7 ± 2.7%, and PECAM+/Podoplanin+: 8.3 ± 0.5%. A percentage of cells positive for one of the lymphatic markers but negative for PECAM were PECAM−/Prox1+: 3.0 ± 1.5%, PECAM−/LYVE-1+: 0.9 ± 0.2%, and PECAM−/Podoplanin+: 6.2 ± 0.8%. Figure 1 shows representative flow cytometry data for one of the five analysis runs.

Representative flow cytometry analysis of mouse SVF sample. Plots represent quantification of cell counts for viability and lymphatic marker (Prox1, LYVE-1, and Podoplanin) labeling versus PECAM (CD31) labeling. Data are gated with nucleated cells. The vertical and horizontal lines per plot indicate quadrant gates based on unstained, control sample (top row). The bottom row displays the data for a stained sample. Ghost Dye (Cytek) was used as the viability label; the dye binds to necrotic cells with compromised cell membranes, and, therefore, viability is indicated by lower-level labeling. The percentages indicate the percentage of cells that are present in the respective region. For example, for the PECAM versus Prox1, plot 15.10% indicates the percentage of cells positive for both PECAM and Prox1. SVF, stromal vascular fraction.
SVF seeded onto avascular mouse mesentery developed branched networks of PECAM+ vessels over the course of incubation (Fig. 2A–E). While on day 1 only individual cells and cell clusters were observed, by day 3 the tissues displayed sprouting from PECAM+ clusters as well as isolated PECAM+ segments. Over the subsequent several days, the number of connected segments increased to form networks. Temporal dynamics of SVF-derived blood vessel and their network formation were consistent with previously reported by us formation of rat SVF-derived vascular networks on cultured rat mesenteric tissues. 29 Furthermore, circular or oval-shaped structures positive for both PECAM+ and LYVE-1+ were observed (Fig. 2F). These multicellular structures were termed “blebs.” In some cases, these blebs were connected to nearby PECAM+/LYVE-1− endothelial capillary segments (Fig. 2I). Capillaries were defined by cords with diameters less than 10 µm. Connections were confirmed by continuity of PECAM+ labeling in submicron confocal microscopic images (Supplementary Fig. S1). A subset of blebs displayed a mosaic cell phenotype characterized by presence of LYVE-1+ and LYVE-1− cells (Fig. 2G). Bleb elongation was observed with extended incubation, which indicates structural remodeling (Fig. 2E, H, I). The blebs were also observed to be positive for Prox1 (Figs. 3 and 4) and example with DAPI co-labeling confirms the presence of cell nuclei (Fig. 4). Remodeling of the PECAM+/LYVE-1+ structures is further supported by quantitative analysis (Fig. 5). Bleb numbers, phenotype, and morphology changed over the course of incubation. While the average size of blebs was consistent, their numbers and degree of elongation increased and fraction of mosaic structures with positive and negative LYVE-1 cells decreased.
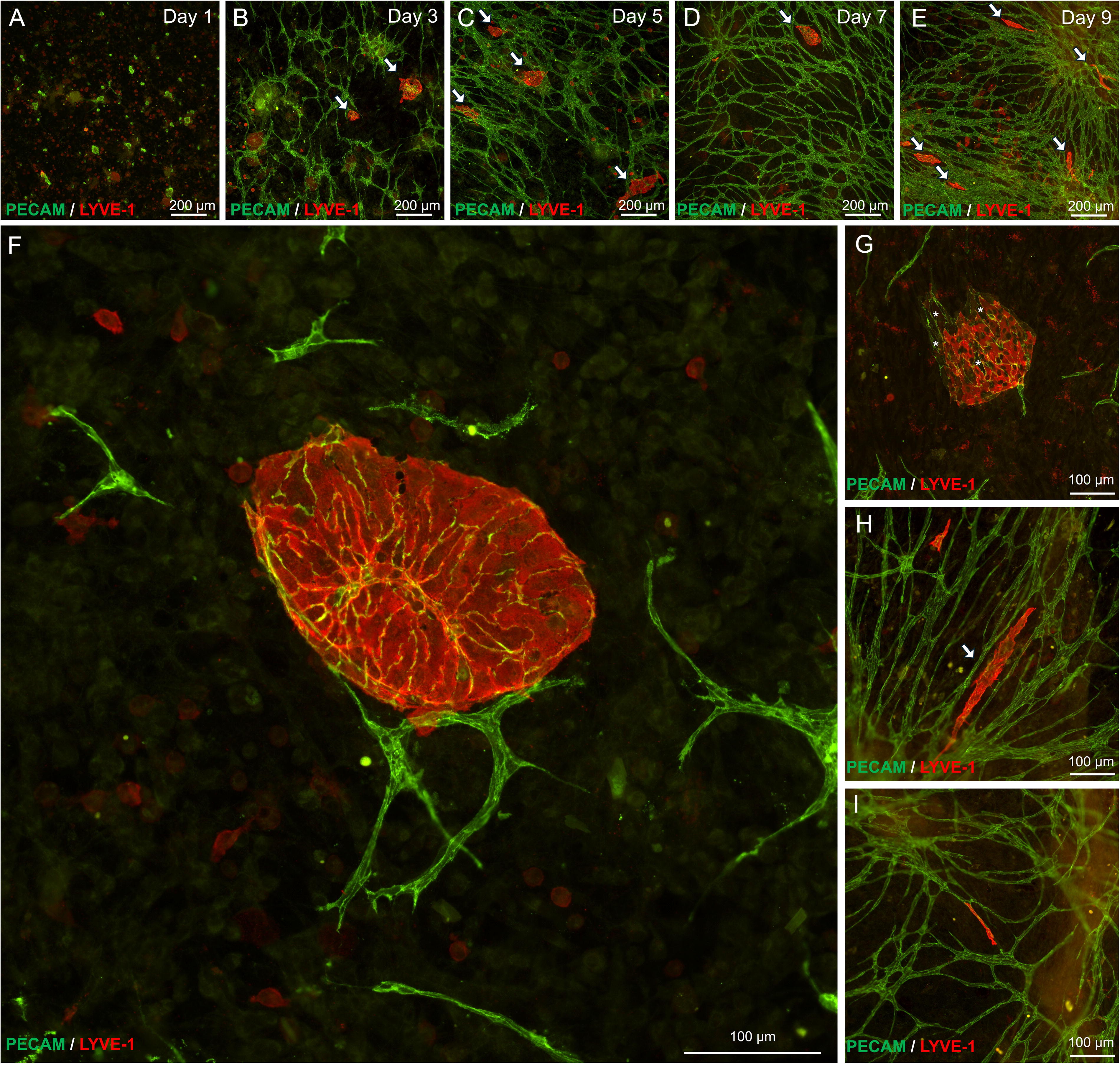
Representative images of SVF-derived blebs. Representative images over the time course of tissue culture
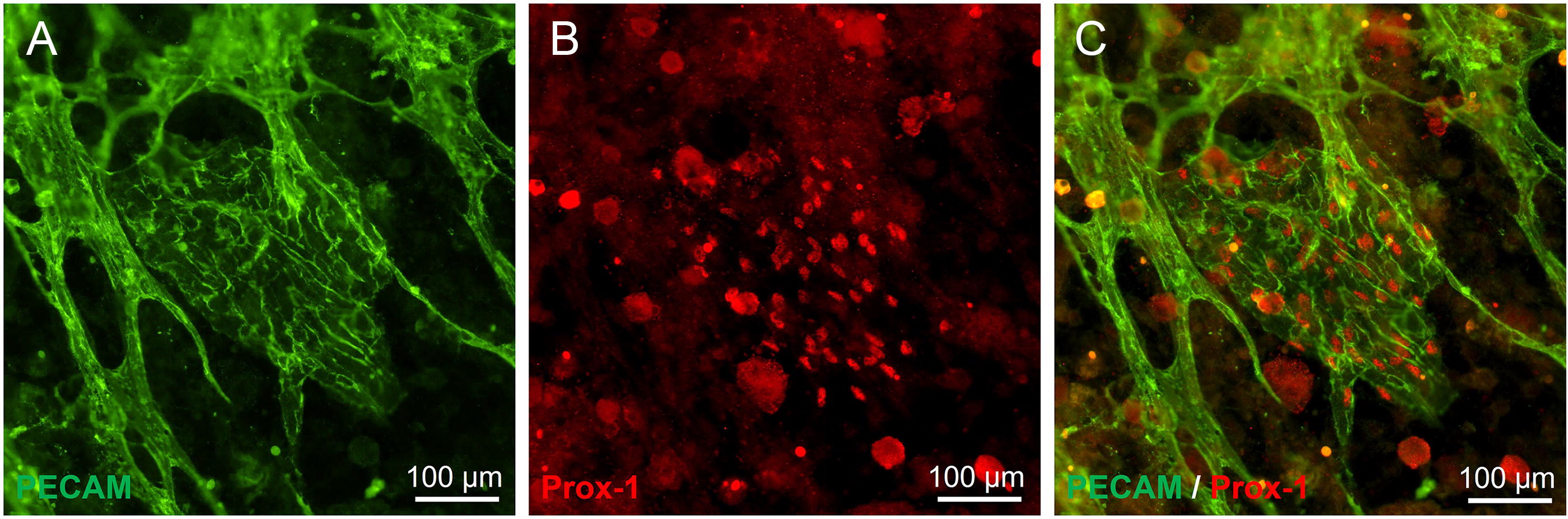
The lymphatic labeling of endothelial cells “blebs.”
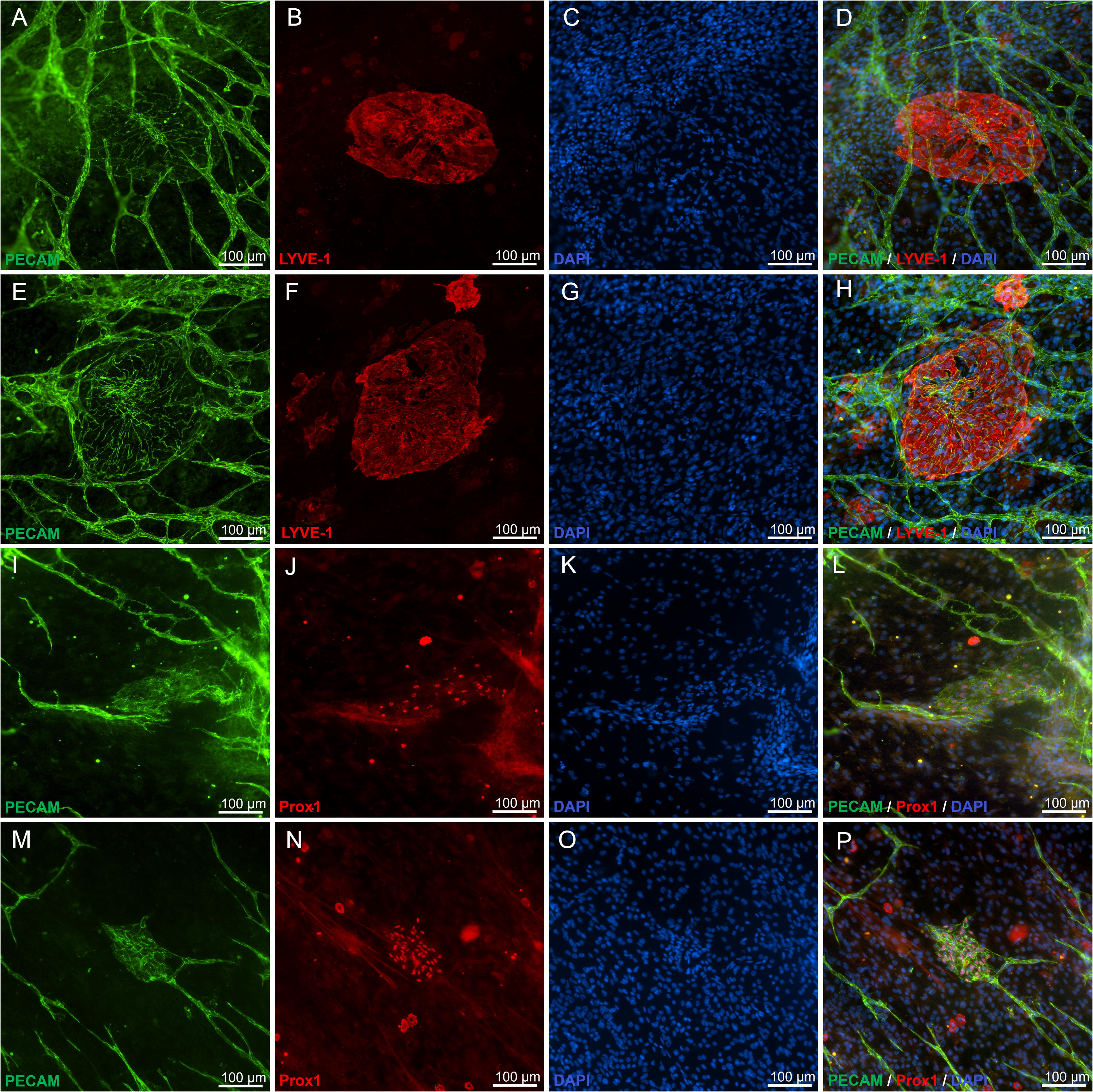
Additional examples of “blebs” labeled for PECAM and LYVE-1 or Prox1. DAPI labeling identifies the nuclei of cells within the 'bleb' structures.
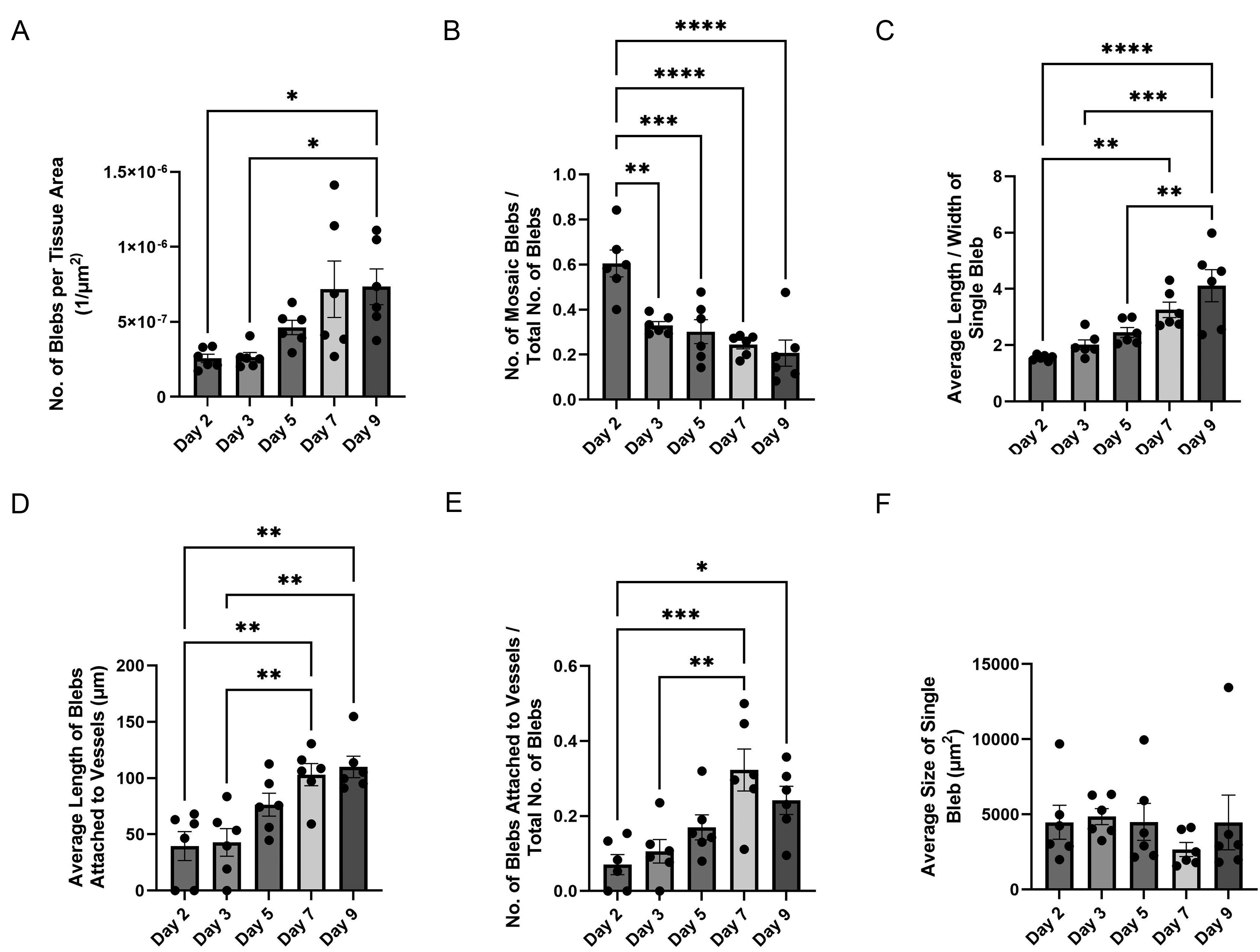
Morphological and phenotypical changes of “blebs” over the time course of tissue culture.
Three additional experiments were conducted to confirm that the lymphatic endothelial cell structures were derived from the donor SVF cells. In one experiment, mouse SVF was transplanted onto rat mesenteric tissues. Post culture, mouse-specific PECAM labeling confirmed that the lymphatic endothelial cell structures were SVF derived (Fig. 6). These observations are supported by a second experiment in which SVF cells isolated from GFP mice were observed to co-localize with LYVE-1+ cell clusters associated with the bleb structures. For the third confirmatory experiment, mouse mesenteric tissues were cultured with serum and without SVF cells. The tissues were void of any PECAM+ vessels.
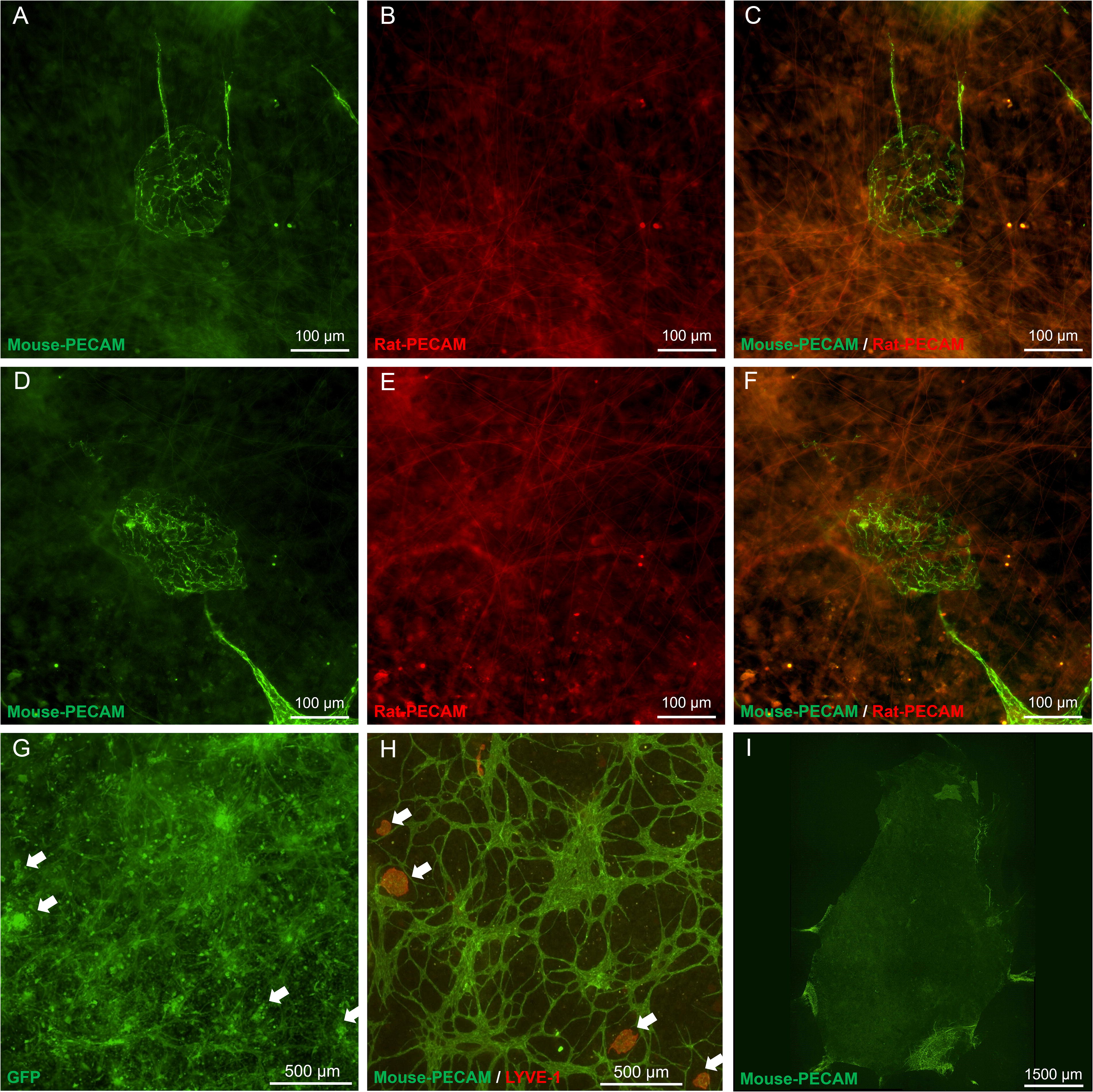
Tracking of endothelial cell “blebs” derived from transplanted SVF. The fate of SVF cells was tracked by transplanting
To verify that the blebs were not an artifact of in vitro culture, an analogous in vivo experiment was conducted. Rat SVF was seeded onto exteriorized rat mesentery tissues. Since rat mesentery has lymphatic vessels, it allowed us to evaluate whether blebs connect with host lymphatic vessels. PECAM+/LYVE-1+ blebs, similar in morphology to those observed in the mouse tissue culture experiments, were observed on day 3 post seeding (Fig. 7). While SVF, used in experiments, was not prelabeled (a limitation of the study), only tissues with seeded SVF established blebs and no blebs were observed sham group (mesentery exteriorization without SVF seeding).
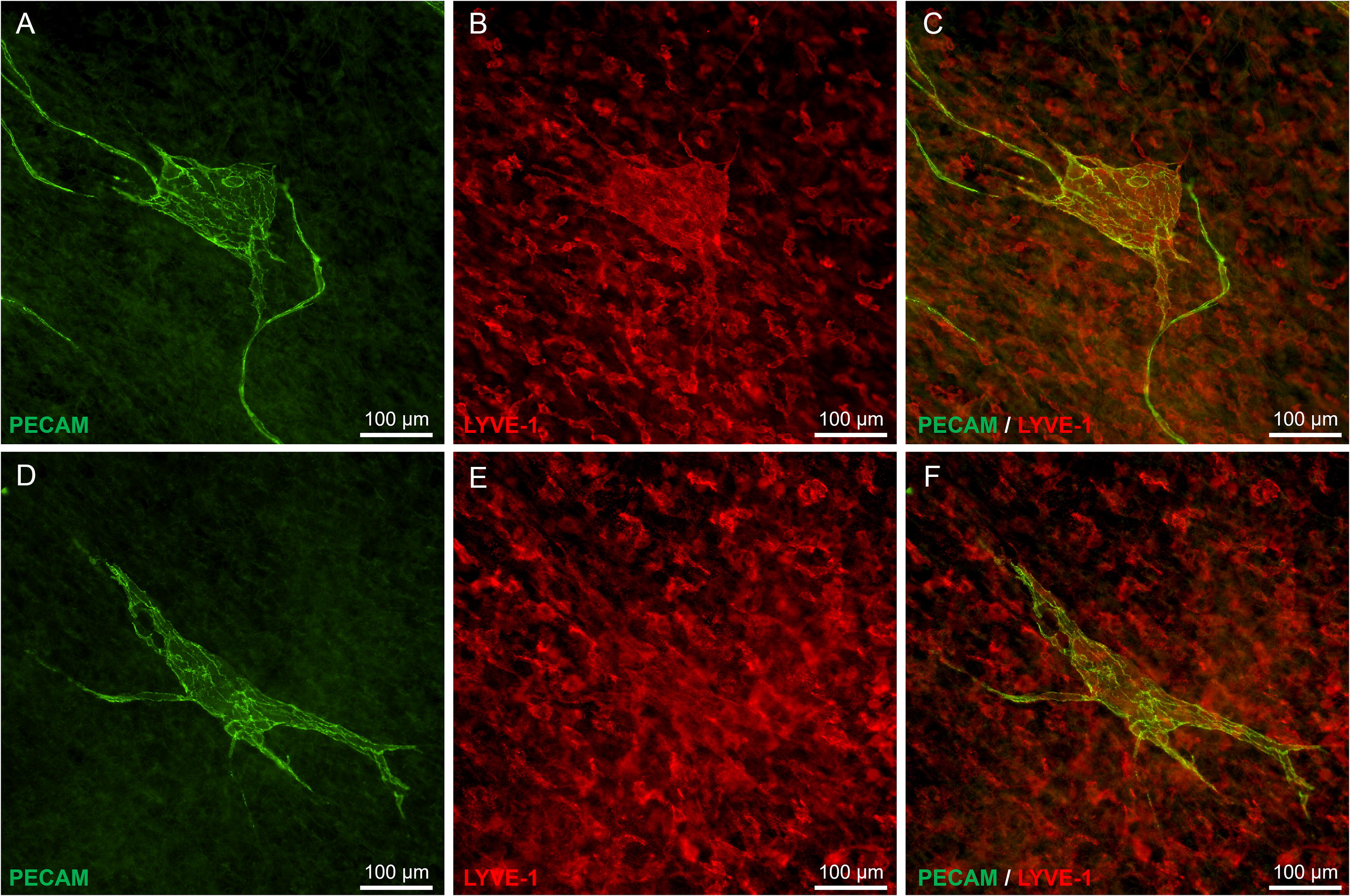
Examples of endothelial cell “blebs” in vivo. Representative images of PECAM+ [
Discussion
The main finding of this study is the identification of lymphatic endothelial cell subpopulations within SVF and formation of lymphatic marker-positive structures from mouse and rat SVF both ex vivo and in vivo. Current observations emphasize a lack of complete understanding of SVF cells fate after transplantation and provide an initial perspective on SVF potential to reorganize into lymphatic vessels. The increased elongation of these structures over the course of incubation and their interconnection with SVF-derived blood endothelial cell cords are interesting observations that motivate follow-up studies to understand whether these structures contribute to the remodeling or regeneration of microvascular networks of both blood and lymphatic vessels. To our knowledge, this is the first report of SVF ability to establish lymphatic endothelial cell-derived structures.
The flow cytometry data support the notion that enzymatic dissociation of adipose tissue preserves a fraction of viable lymphatic endothelial cells, capable of self-organization. Prox1, LYVE-1, and Podoplanin were selected for this study as the most prominent markers of lymphatic endothelial cells. The flow cytometry data, shown in Fig. 1, represent an example for one of the five analysis runs. For the Prox1 analysis data in the figure, the percentage of PECAM+, Prox1+ cells is 15.1%. The corresponding average for the five flow cytometry runs was 5.6% with the raw percentages being 0.3%, 0.9%, 4.2%, 15.1%, and 7.3%. The different percentages across runs suggest that the presence of Prox1 cells can be variable and support the presence of subsets of lymphatic endothelial cells. To confirm that our results could be replicated when controlling IgG isotype labeling, we conducted an additional validation study with three analysis runs. Representative data for one of the runs are shown in Supplementary Fig. S2. The percentage of cells positive for PECAM and one of the lymphatic markers based on the IgG isotype comparisons was PECAM+/Prox1+: 5.1 ± 2.3%, PECAM+/LYVE-1+: 3.5 ± 0.3%, and PECAM+/Podoplanin+: 4.6 ± 0.4%. Interestingly, cells positive for lymphatic marker were either PECAM+ or PECAM−, raising questions related to whether expression of lymphatic markers is limited to endothelial cells and whether lymphatic endothelial cells are heterogeneous for PECAM expression. Along this thinking, our results provoke similar types of questions. What is the percentage of SVF cells that express all three lymphatic markers in SVF? What are the cell types that express lymphatic markers but not PECAM? Additional analysis of our flow cytometry data indicates that the percentage of live cells positive for PECAM and positive for the two lymphatic markers is less than 0.07% and the percentage of live cells negative for PECAM and positive for the three lymphatic markers is less than 0.09% (data not shown). The take-home message from our flow cytometry cell is that SVF contains an initial presence of lymphatic endothelial cells, and this finding can be corroborated with PECAM and LYVE-1 labeling of the cultured mouse tissues with SVF on day 1 before the observation of the bleb structures (Supplementary Fig. S3).
The strength of this study is that activities of mouse SVF were explored on mouse mesentery tissue, which, compared with alternative in vitro or ex vivo models, provides a native tissue environment. Furthermore, its thinness allows for whole mount labeling and en face imaging. As noted in the Introduction, there are only a few reports that include SVF and lymphatics or lymphangiogenesis. In one study by Emont et al., the comprehensive single cell characterizations of SVF suggest the presence of lymphatic endothelial cells based on Prox1 expression with percentages of Prox1 positive cells ranging from 0.4% to 2.6% based on species and fat depot. 37 While comprehensive, the single-cell atlas study does not provide evidence for the fate or function of the lymphatic endothelial cell subpopulation in a remodeling scenario. In the sole example in the literature suggesting that SVF-derived cells can form lymphatic vessels, Vuerich et al. 38 purified SVF cells from different transgenic animals to trace their fate post transplantation into a subcutaneous tissue surrounding an ischemic wound and observed SVF-derived arteries, capillaries, veins, as well as lymphatics. Evidence for lymphatic vessels was based on LYVE-1 labeling of tissue cross sections. In the context of both studies, our flow cytometry results add support for a subpopulation of lymphatic endothelial cells in SVF by showing positive labeling with positive for multiple lymphatic markers, and our documentation of formation of blebs positive for lymphatic markers highlights the importance of en face views and raises questions about potential shape and cellular organization of SVF-derived lymphatic structures.
Study limitations include not exploring the function of the SVF-derived lymphatic structures or whether the structures organize into a network of lymphatic vessels at later time points in vivo. While our analyses focused on the development of SVF-derived structures, consideration of potential functional or dysfunctional implications is important for appreciating therapeutic implications, and our results motivate future characterization studies. Regarding the potential role of the structures in network formation, we speculate that clusters are composed of progenitors of lymphatic endothelial cells. Martinez-Corral et al. discovered that non-venous-derived lymph sacs are clusters of progenitor cells that give rise to lymphatic vessels. 39 Interestingly, the structure of our SVF-derived lymphatic blebs is similar to the lymph sacs. Follow-up studies are needed to determine whether lymphatic blebs represent an initial stage of lymphatic vessel organization.
Mouse mesentery is avascular tissue, 36 which allowed us to assume that vessels formed during culture were SVF derived. Our mouse culture studies, therefore, suggest that the lymphatic endothelial cell bleb structures were indeed SVF derived. The SVF-derived origin of the lymphatic endothelial cell structures is further supported by our follow-up confirmatory observations based on tracking the fate of GFP-positive SVF cells, the labeling of mouse SVF-derived endothelial cells versus host endothelial cells, and stimulation of cultured mouse tissues without SVF (Fig. 6). Altogether, our results support that lymphatic bleb structures are indeed SVF derived and suggest against the possibility that host-derived cells in the mesenteric tissue could be induced into lymphatic endothelial cells.
The in vivo observations suggest support the relevance of our culture results and also support that these structures form in a vascularized host tissue with lymphatic vessels. Rats were used for this verification study since rat tissues are vascularized with both blood and lymphatic microvascular networks. Based on the ex vivo observations, we wanted to determine if similar structures formed in vivo and explore whether the structures interacted with host initial lymphatic networks. Interestingly, while we did observe similar structures, we did not observe any connection of these structures to the preexisting lymphatic networks. Future studies at longer time points are needed to further characterize the fate of the structures.
Optimal incubation conditions for SVF culture on top of live tissues are unknown. Based on our previous work characterizing SVF formation of blood microvessels in cultured rat mesentery tissues, 29 we supplemented the incubation media with 10% serum. Use of specific growth factors could provide insights into whether SVF-derived lymphatic structures can be regulated by the local cues. To test it, additional trial studies were conducted for which incubation media was supplemented with VEGF-C (Supplementary Fig. S4). No change in blebs’ average size or number in response to VEGF-C were detected on day 3 of cultivation; however, VEGF-C increased the fraction of structures with mosaic phenotypes and the number of structures attached to apparent blood endothelial segments. The additional results support the notion that local environment can influence the remodeling of the lymphatic endothelial cell structures. Future experiments are needed to probe the importance of the VEGF-C effects and the potential for other specific factors to regulate the formation of the SVF-derived structures.
Conclusions
Coordination between blood and lymphatic vessels at microvascular level is critical for tissue homeostasis, interstitial fluid flow, and immune cell trafficking. Our study represents an initial step into a new area of research exploring whether SVF contributes to lymphatic vasculature development, as demonstrated by its ability to form lymphatic endothelial structures on mesentery tissue within a native extracellular matrix. The results of this study suggest SVF can form lymphatic endothelial cell structures and motivate future work to understand their fate and function. Overall, the observations additionally emphasize the need to explore the causal mechanisms associated with their formation, the specific involvement of both SVF and tissue resident cell populations, and the potential for the structures to form in other tissues.
Footnotes
Acknowledgment
We would also like to thank all members of Dr. Murfee’s Microvascular Dynamics Laboratory who helped contribute to the initial studies effort that went into producing this publication.
Author Disclosure Statement
The authors declare no other potential conflicts of interest.
Funding Information
Funding for this project was supported by the National Institutes of Health (grant R21HL159501).
Supplementary Material
Supplementary Figures
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.