
Abstract
Exposure to high-dose radiation often results in hematopoietic acute radiation syndromes, leading to early mortality, while current therapies for patients exposed to lethal radiation doses are limited. Mesenchymal stem cell-derived extracellular vesicles (MSC-EVs) have shown promise in tissue repair and regeneration but have not been well investigated for mitigating high-dose radiation damage. We previously demonstrated that human or murine MSC-EVs can reverse bone marrow injury caused by mild or moderate radiation. The current study evaluated the therapeutic potential of human MSC-EVs in mice exposed to high-dose total body irradiation (TBI). Mice were exposed to 0, 700, or 950 cGy TBI and subsequently received daily intravenous MSC-EV injections (1 × 109 particles) for 3 days postirradiation. We evaluated survival rates, peripheral blood recovery, bone marrow engraftment, and bone marrow gene expression profiles at various intervals following treatment. MSC-EV administration significantly enhanced survival, with 70% of treated mice surviving 120 days after 950 cGy TBI exposure, compared with 0% survival in untreated controls by day 30. Although early peripheral blood recovery was not observed within 14 days, MSC-EV treatment facilitated substantial recovery at 3 months postirradiation, with significant increases in red blood cell, platelet, white blood cell, and hemoglobin levels, despite white blood cell and hemoglobin levels remaining slightly below normal. Furthermore, the engraftment capacity of bone marrow stem cells was significantly improved. The changes in hematopoietic-related gene expression presented at 14 days postirradiation returned to normal levels by 120 days in MSC-EV-treated mice. These results highlight the potential of MSC-EVs as a therapeutic strategy for high-dose radiation injuries by promoting hematopoietic recovery and improving survival. Our future research will focus on elucidating the radioprotective mechanisms and investigating their integration with existing therapies.
Introduction
Widespread radiation exposure from a large scale radiological or nuclear incident represents a constant threat. Individuals exposed to radiation from terrorist activity, radiation accidents, or radiological warfare will have a variety of acute and chronic organ injuries. Acute radiation exposure (ARE) can lead to hematopoietic, gastrointestinal, and neurovascular syndromes, as well as damage to other tissues and organs. Among these syndromes, hematopoietic-acute radiation syndrome (H-ARS), is a major cause of mortality after exposure to high doses of radiation. This is primarily due to severe injuries to hematopoietic stem cells, resulting in pancytopenia and subsequently leading to infection and/or hemorrhage. 1,2
The current therapies for patients exposed to potentially lethal doses of radiation are limited. Several radiation mitigators have been studied, but only Granulocyte Colony-Stimulating Factor (G-CSF) has been approved by the FDA for the treatment of ARS. Although potentially effective, hematopoietic cellular transplants are not generally available in a timely manner. The results with G-CSF and other mitigators at present appear suboptimal. 3 As a result, there is a growing need for effective rescue and treatment measures in the event of radiation emergencies.
Extracellular vesicles (EVs) are small membrane-bound particles secreted from various cells throughout the body. EVs consist of two major types of vesicle populations: microvesicles, which are shed from the plasma membrane, and exosomes, formed through reverse budding of multivesicular bodies. 4,5 EVs carry many bioactive molecules, including proteins, DNA, lipids, mRNA, and noncoding RNA such as miRNAs, which are derived from parental cells and play a critical role in cell communication. 6,7 EVs derived from stem cells have been shown to have a significant ability to repair damaged tissues. 8,9 In particular, mesenchymal stem cell-derived EVs (MSC-EVs) have shown a significant ability to promote tissue repair and regeneration in various conditions, including myocardial ischemia-reperfusion injury, liver, kidney, and respiratory disorders. 10 –15 Currently, there are 1,135 clinical trials registered on clinicaltrials.gov using autologous and allogeneic MSCs to treat various categories of human diseases and medical conditions. However, clinical data remain inconclusive regarding the advantages of autologous versus allogeneic MSCs. 16,17 MSC-EVs are considered an acellular biological product with several advantages over MSCs including low immunogenicity, long half-life, in vivo stability, and efficient delivery capacity. 18,19 Presently, 15 clinical trials are underway evaluating allogeneic MSC-EVs for various disease categories, with preliminary data indicating minimal toxicity concerns. We previously demonstrated that human or murine MSC-EVs would completely or partially reverse bone marrow injury elicited by exposure to mild or moderate radiation in vitro or in vivo through the inhibition of apoptosis and DNA damage. The entire MSC-EV population, including both microvesicles and exosomes, was more effective in reversing radiation-induced injury than either microvesicles or exosomes alone. 20 The goal of this study is to further test whether we could reverse marrow stem cell damage and improve survival in mice exposed to high or lethal doses of radiation in an in vivo mouse model by administering human MSC-EVs.
Materials and Methods
MSCs culture and EV isolation
Human bone marrow-derived MSCs were purchased from Lonza (Catalog# PT-2501, Walkersville, MD, USA) cultured, and expanded in Minimum Essential Medium (MEM) α media (Fisher Scientific) with 15% MSC-qualified fetal bovine serum (Fisher Scientific), in an incubator at 37°C with 5% CO2. When the cells reached about 80%–90% confluent in T-175 flasks, the flasks were rinsed with Phosphate-Buffered Saline (PBS) two times and cultured with Roswell Park Memorial Institue (RPMI) 1640 media (Fisher Scientific) for 24 h. The conditioned culture media was then harvested and centrifuged with WX Ultra Centrifuge with Sorvall AH-629 rotor (Thermo Fisher Scientific) at 2,000g for 30 min to remove cell debris and large particles, followed by 1,00,000g for 70 min at 4°C to isolate the MSC-EVs. The pelleted MSC-EVs were then washed with PBS with an additional 70 min centrifuge cycle at 1,00,000g. The MSC-EV pellet, containing both microvesicle and exosome populations, was resuspended in PBS with 1% Dimethyl Sulfoxide (DMSO) and stored at −80°C until further use.
EV characterization
The size distribution and concentration of EVs were analyzed using the NanoSight NS500 (Malvern Instruments, Malvern, UK) equipped with a syringe pump for accurate sample delivery. Nanoparticle tracking analysis (NTA) was performed with NanoSight NTA v3.4 software (Malvern Pananalytical Ltd, Worcestershire, UK), following previously established protocols. 20,21 All samples were analyzed in triplicate to ensure that the results were reliable and reproducible.
EVs were further characterized by transmission electron microscopy (TEM) as previously described. 20,21 Briefly, 2% paraformaldehyde was used to fix the MSC-EV samples, then placed onto Formvar-carbon-coated electron microscopy grids, and further fixed with 1% glutaraldehyde and contrasted with 4% uranyl acetate. The EV samples were then visualized under a transmission electron microscope (FEI Morgagni 268).
The presence of EV-specific markers was confirmed by Western Blot, including CD81 (System Biosciences# EXOAB-CD81A-1), CD9 (System Biosciences # EXOAB-CD9A-1), CD63 (System Biosciences # EXOAB-CD63A-1), TSG101 (Thermo Fisher #MA1-23296), PDCD6IP (Thermo Fisher # 50-167-7058), beta actin (Sigma #A1978-200UL), heat shock protein 70 (HSP70) (Santa Cruz #sc-59560), and albumin (ALB) (Santa Cruz #sc-58688).
Animal experiments
All animal studies were approved by the Institutional Animal Care and Use Committee at Rhode Island Hospital. The 6–8-week-old male C57BL/6 and B6.SJL mice were purchased from Jackson Laboratory. C57BL/6J mice were exposed to total body irradiation (TBI) at doses of 0, 700, and 950 cGy, with the 950 cGy dose administered as a split dose of 475 cGy over 3 h (Gammacell 40 Exactor, Cesium 137 source irradiator, 0.94–0.96 Gy/min). For the MSC-EV treatment group, mice were injected with 1 × 109 human MSC-EVs by tail vein injection per day for 3 days at 1-day postirradiation and the control group mice were injected with vehicle only. Peripheral blood samples were collected by tail vein bleeding into Ethylendiaminetetraacetic acid (EDTA)-coated tubes, and complete blood counts were performed on HemaTrue Veterinary Hematology Analyzer (Heska Corp., Loveland, CO, USA). In this study, we used a competitive transplant mode to determine the engraftment capacity of bone marrow stem cells. The recipient (host) mice received 950 cGy in two split fractions (475 cGy/fraction) 3 h before transplantation. The bone marrow cells from donor mice (CD45.2+; C57BL/6) and congenic competitor healthy mice (CD45.1+; B6.SJL) were mixed at a 1:1 ratio and injected into the tail vein of recipient mice (CD45.1+; B6.SJL). Donor chimerism in peripheral blood was evaluated using BD LSR II flow cytometer (BD Biosciences) with antibodies against CD45.1, CD45.2, B220, CD3, CD11b, and GR-1 (BD Biosciences). The donor engraftment rate was determined as the ratio of CD45.2 (donor) cells to the total number of CD45.1 and CD45.2 cells.
Realtime PCR
Total RNA was isolated from whole bone marrow cells (WBMCs) using the miRNeasy mini kit (Qiagen) according to the manufacturer’s protocol. Reverse transcription was conducted with the high-capacity cDNA transcription kit (Life Technologies) under the following conditions: 10 min at 25°C, 120 min at 37°C, and 5 min at 85°C, using a 9800 Fast Thermal Cycler (Applied Biosystems). Pre-amplification reaction was then conducted with the TaqMan Preamp Master Mix Kit (Life Technologies) according to the manufacturer’s instructions. The reaction conditions included an initial step of 10 min at 95°C, followed by 14 cycles of 15 s at 95°C and 4 min at 60°C. The diluted pre-amplified DNA sample was mixed with TaqMan Universal PCR Master Mix and the specific primers (Thermo Fisher Scientific) listed in Table 1 and analyzed on a ViiA 7 real-time PCR instrument using QuantStudio™ software (Applied Biosystems). Relative gene expression across samples was analyzed using the 2−ΔΔCT method with ABI ExpressionSuite 1.1Software (Applied Biosystems). β2 microglobulin (B2M) was used as the endogenous control gene and the control group sample was designated as the reference for calculating the relative expression of each target gene cross the samples. Genes with CT values greater than 35 were considered to be undetectable expressions. TaqMan primers were purchased from Applied Biosystems.
Gene List of TaqMan® Assays (Thermo Fisher Scientific)
Statistical analysis
Prism 10.0 (GraphPad Software, Inc) was used for statistical analyses including statistical comparisons between groups (student t-tests or ANOVAs with multi-comparison tests) and Kaplan–Meier survival curve analysis with the log-rank (Mantele-Cox test.) A P value of <0.05 was considered statistically significant. Gene expression heatmap and principal component analysis (PCA) were performed using SRplot for data visualization and graphing. 22
Results
Human MSC-EV characterization
MSC-EV characterization was evaluated following the MISEV2018 criteria. 23 The concentration and size distribution of human MSC-EVs were analyzed by NanoSight 500 (NanoSight, Malvern, UK) according to the manufacturer’s instructions (Fig. 1A). The average MSC-EV size was determined to be 205.2 ± 99.3 nm. TEM (FEI Morgagni 268) was used to further confirm the size and morphology of EVs (Fig. 1B). Western blot analysis demonstrated the presence of surface epitope proteins CD9, CD63, and CD81, as well as cytosolic EV proteins PDCD6IP, TSG101, HSP70, and β-actin, in MSC-EVs, while the negative control protein ALB was not detected (Fig. 1C).
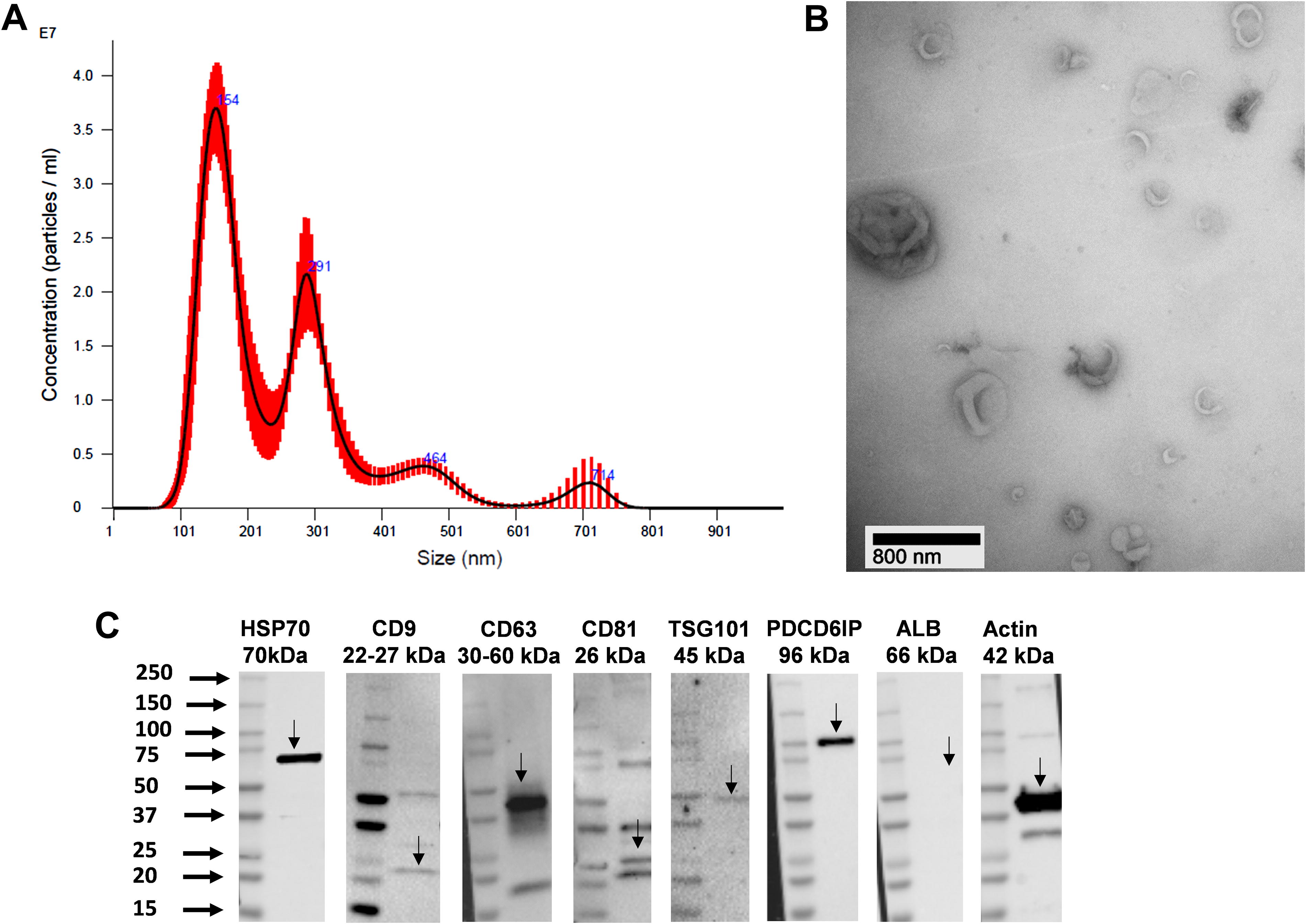
Characterization of MSC-EVs:
Human MSC-EVs increase survival of mice exposed to a lethal dose of TBI
We first investigated the potential of MSC-EVs to protect against lethal TBI in C57BL/6J mice. C57BL/6J mice were exposed to 950 cGy, with a split dose of 475 cGy over 3 h. At 24 h postirradiation, the treatment group received tail vein injections of 1 × 109 human MSC-EVs per day for 3 days, while the MSC-EV untreated control group received vehicle only. At 30 days postirradiation, none of the mice in the MSC-EV untreated control group survived, with all mice (n = 10 mice) dying between 12–27 days postirradiation, including 7 of 10 within the first 2 weeks. This resulted in a 0% survival rate at day 30 post 950 Gy TBI in C57BL/6J mice (Fig. 2). However, the MSC-EV treated group exhibited a 70% survival rate extending to 120 days postirradiation. Only three mice in the treatment group died 14, 15, and 16 days after radiation, respectively.
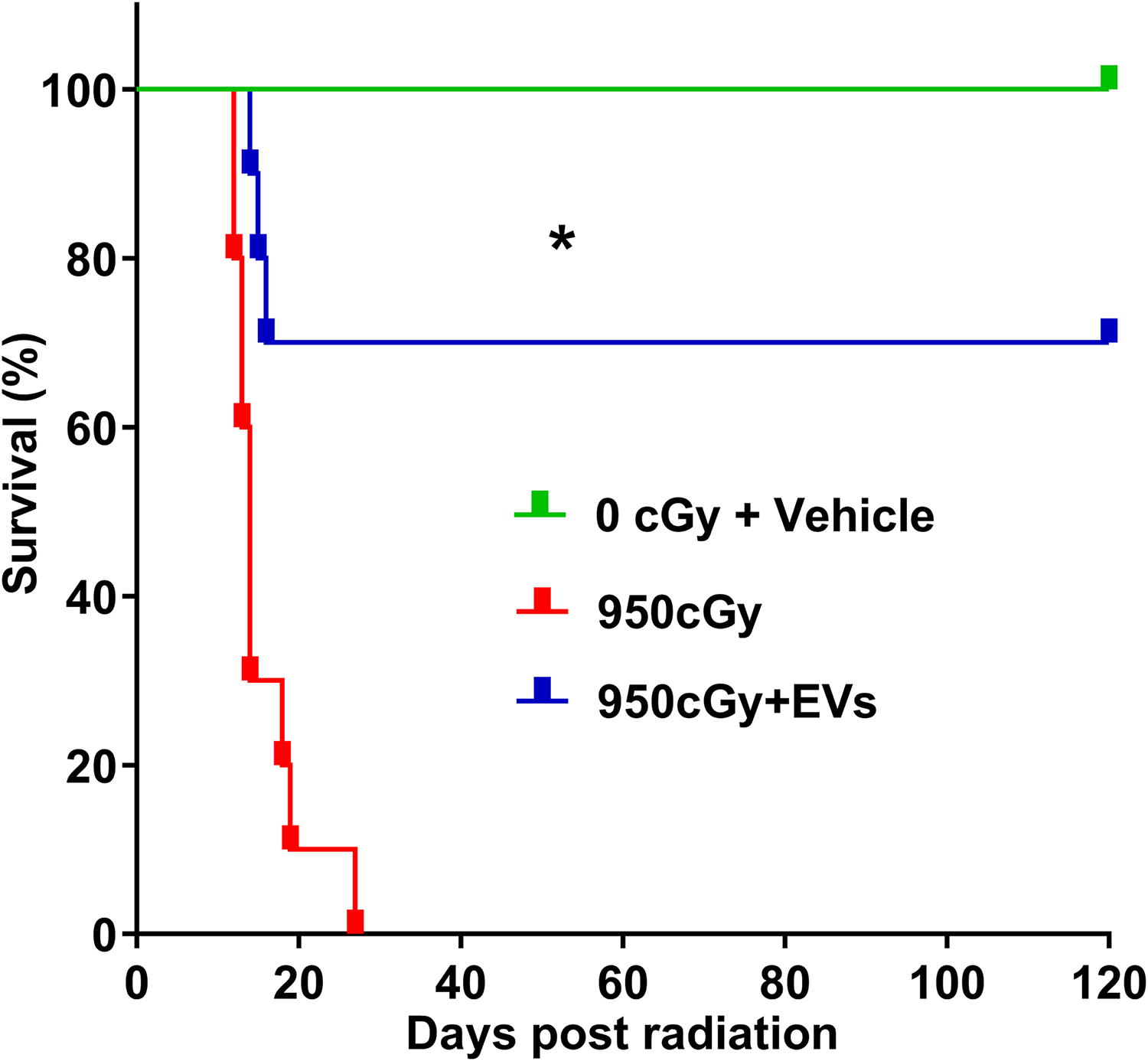
Kaplan–Meier survival curve showing the survival of mice exposed to 950 cGy total body irradiation with or without MSC-EV treatment. Differences in survival were determined by log-rank (Mantel-Cox) analysis, n = 10 mice/group, *P < 0.001, 950 cGy versus 950 cGy + EVs group.
Effects of MSC-EVs on reversal of peripheral blood and bone marrow cells from mice exposed to radiation
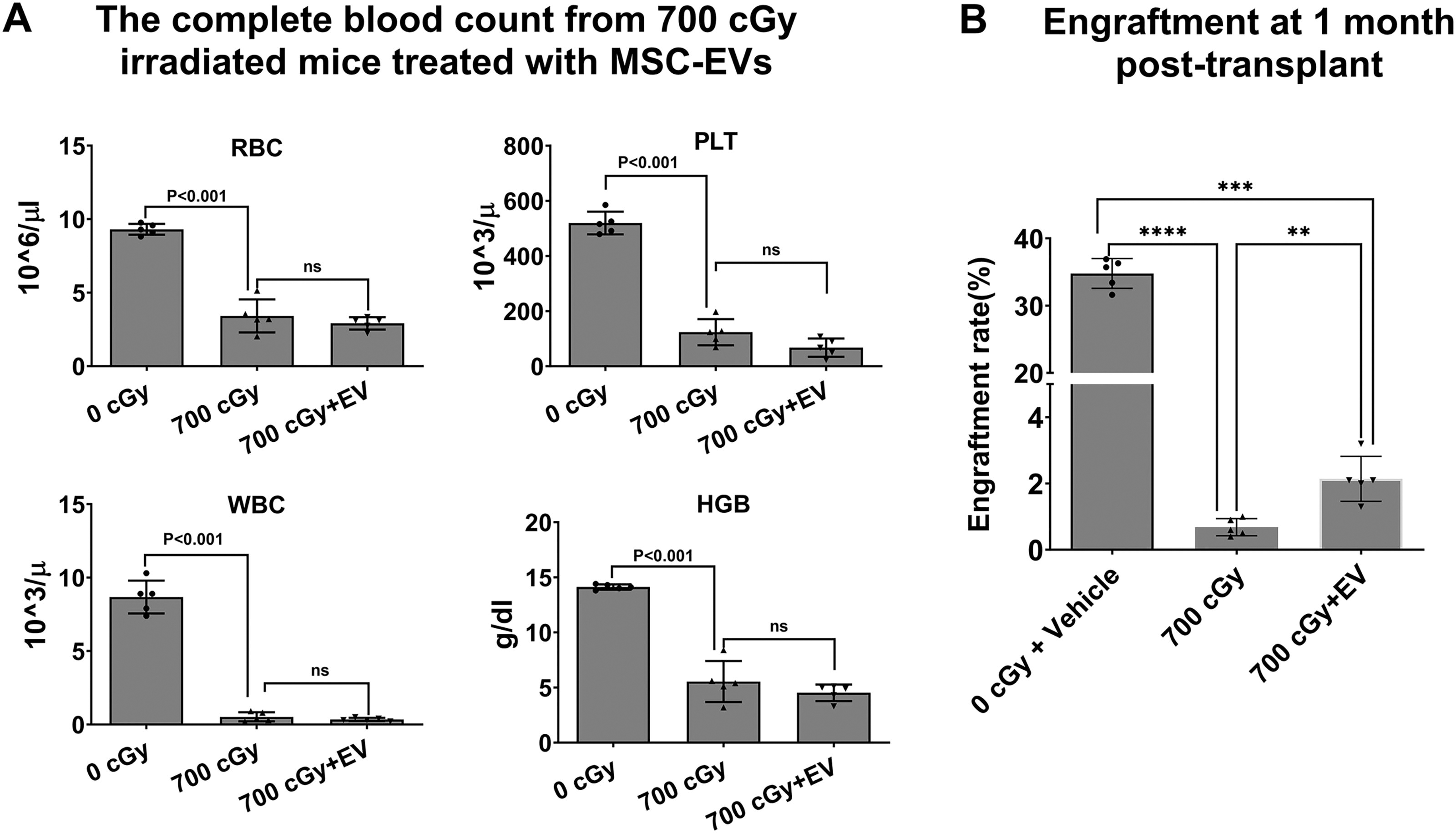
We next evaluated the effect of MSC-EVs on the reversal of high dose of 700 cGy radiation injury in bone marrow stem cells. C57BL/6 mice received 0 or 700 cGy TBI. At 24 h postirradiation, the mice received daily intravenous infusions of 1 × 109 MSC-EVs for 3 days. MSC-EV untreated control mice received the vehicle only. The peripheral blood counts from the mice by tail vein bleeds were analyzed by HemaTrue Veterinary Hematology Analyzer at 14 days postirradiation. There was a significant reduction in white blood cell (WBC), Red blood cell (RBC), hemoglobin (HGB), and platelet (PLT) in mice exposed to 700 cGy radiation compared with nonirradiated mice (Fig. 3A). However, no significant peripheral blood cell restoration was observed in EV-treated mice (N = 5 mice per group, analyzed using ordinary one-way ANOVA with multiple comparisons). To further evaluate the restorative effects of MSC-EVs on the stem cell compartment, we performed a bone marrow transplant study where we harvested bone marrow from irradiated hosts and infused them into lethally irradiated mice. Two million WBMCs from C57BL/6 mice exposed to 700 cGy TBI, with or without MSC-EV treatment at 14 days postirradiation from the above experiment, were cotransplanted with an equal number of WBMCs from healthy B6SJL mice into lethally irradiated B6SJL recipients. Mice treated with MSC-EVs showed significantly higher engraftment rates (2.14% ± 0.68%) than untreated controls (0.68% ± 0.26%) at 1 month post-transplantation (Fig. 3B).
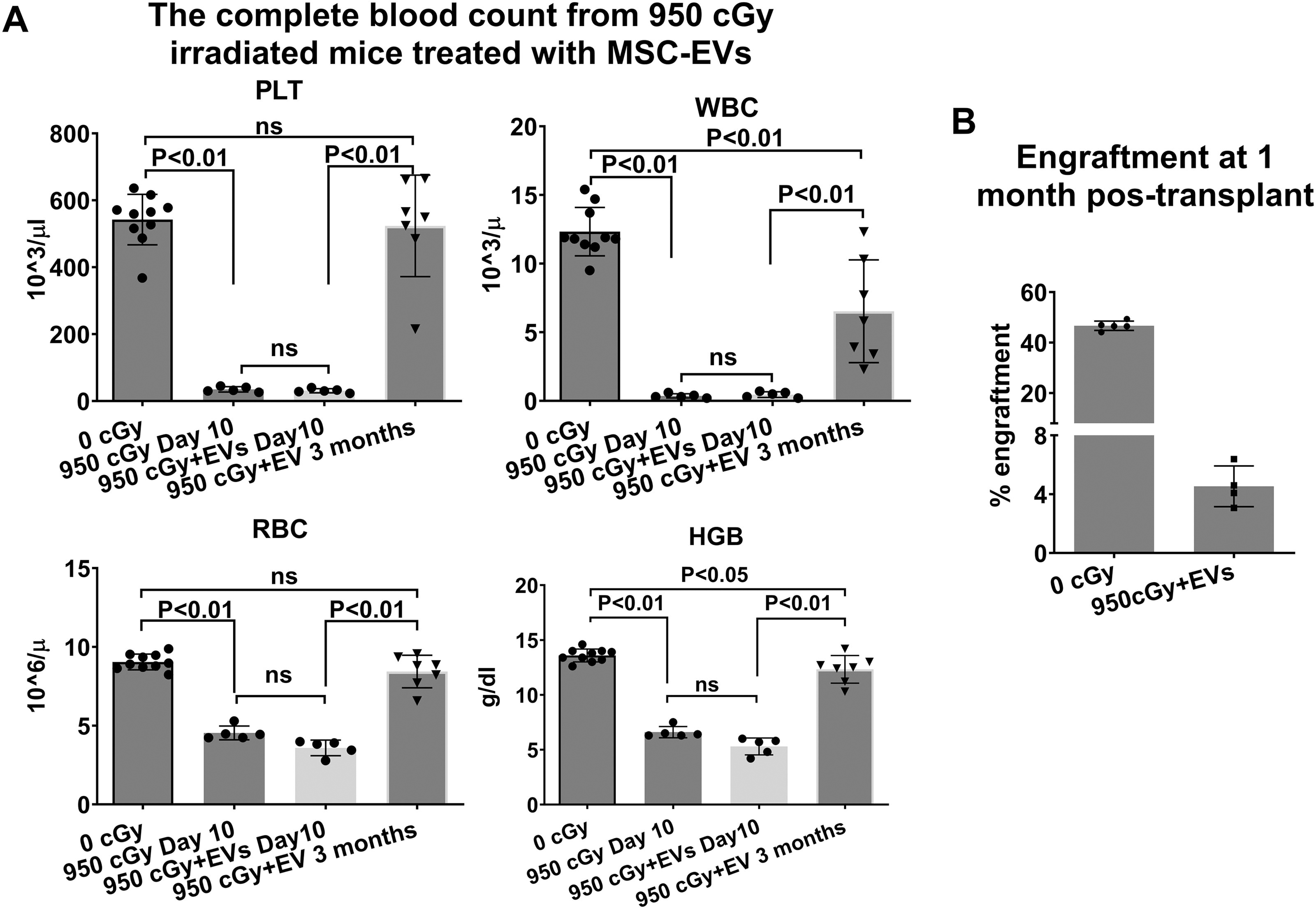
We then further evaluated the effect of MSC-EVs on the reversal of radiation injury in mice exposed to 950 cGy with a split dose of 475 cGy in 3 h. MSC-EV-treated mice received 1 × 109 human MSC-EVs daily for 3 days starting 24 h postirradiation, while controls received vehicle only. We also observed peripheral blood counts significantly reduced, at ten days after radiation with/without EV treatment and there was no significant peripheral blood cell restoration in EV-treated mice compared with EV-untreated mice. However, the peripheral blood cell counts including RBC, PLT, WBCs, and HGB significantly recovered after 3 months of EV treatment although WBCs and HGB were still lower than the normal control (Fig. 4A, Table 2). Because all untreated mice died by 30 days postirradiation, no data were available for the untreated mice at this 3-month time point. We were unable to evaluate the engraftment capacity of bone marrow cells from mice 10 days post-950 cGy radiation, with or without EV treatment, due to insufficient bone marrow stem cell (BMSC) collection for competitive transplantation during this period (only about one million WBMCs were collected from a mouse). However, we observed an engraftment rate of 4.53% ± 1.39% of WBMCs from MSC-EV-treated mice 3 months after 950 cGy radiation at 1 month post-transplant (Fig. 4B).
Peripheral Blood Counts at 10 Days and 3 Months Postirradiation a
Data are shown as the mean ± SD, n = 5–10 mice/group. Data were analyzed using Student’s t-test.
P < 0.05 versus 0 cGy control group.
P < 0.05 versus 0 cGy control group.
P < 0.05 versus 0 cGy control group.
versus 950 cGy Day 10 group: WBC, P = 1.00; PLT, P = 1.00; RBC, P = 0.15; HGB, P = 0.09.
MSC-EVs promote gene expression recovery in BMSCs from lethally irradiated mice
We further analyzed the expression of 30 marrow stem cell-related genes in WBMCs from mice with or without EV treatment at 14 and 120 days postexposure to 0 and 950 cGy radiation using real-time PCR analysis. The differential expression genes (DEGs) were identified using cutoff values of |log2FoldChange(log2FC)| ≥ 1.0 and P < 0.05 (t-test). Upregulated DEGs were defined as log2FC ≥ 1 and downregulated DEGs as log2FC ≤ −1. Fifteen DEGs were identified between groups receiving radiation with or without EV treatment at 14 days postirradiation and nonirradiated control mice (Table 3). Within the 15 DEGs, the gene expression of CXCL12, CD90, and Nestin was upregulated, while 7 genes including SATB1, RUNX1, PBX1, CKIT, CD34, CD33, and S100A9 were downregulated in mice at 14 days postirradiation with/without EV treatment compared with the nonradiation control mice. The remaining four genes (PECAM, MPO, SPI1, and CD105) showed significant alterations in EV-treated mice but exhibited similar alteration trends in the EV untreated group without matching the criteria for DEG compared with the nonirradiated control group. Using the same criteria, no DEGs were found between mice with and without EV treatment at 14 days postirradiation except the gene CD3E was upregulated by radiation but came down to normal on day 14 with EV treatment. We observed very similar gene expression patterns between the nonirradiated control mice group and the EV treatment group at 120 days post-EV treatment, with only MCAM and CXCR4 upregulated at day 120 post-EV treatment, compared with nonirradiated control mice. The differential expression of these 30 genes in mice was then illustrated in the heatmap across these five experimental groups (Fig. 5A): A (0 cGy day 14), B (950 cGy day 14), C (950 cGy + EVs day 14), D (950 cGy + EVs day 120), and E (0 cGy day 120). Dark orange squares represent higher relative expression, while yellow squares indicate lower expression. Each column corresponds to a sample mouse within the respective groups, while rows represent individual genes. The expanded view of the DEG heatmap (Fig. 5B) presents the significant expression differences among groups. Next, we performed the PCA on the gene expression profiles of all 24 mice. It showed that the nonirradiated control mice clustered closely with EV-treated mice at day 120 postirradiation. They were clearly separated from the mice at day 14 postirradiation with or without EV treatment (Fig. 5C).
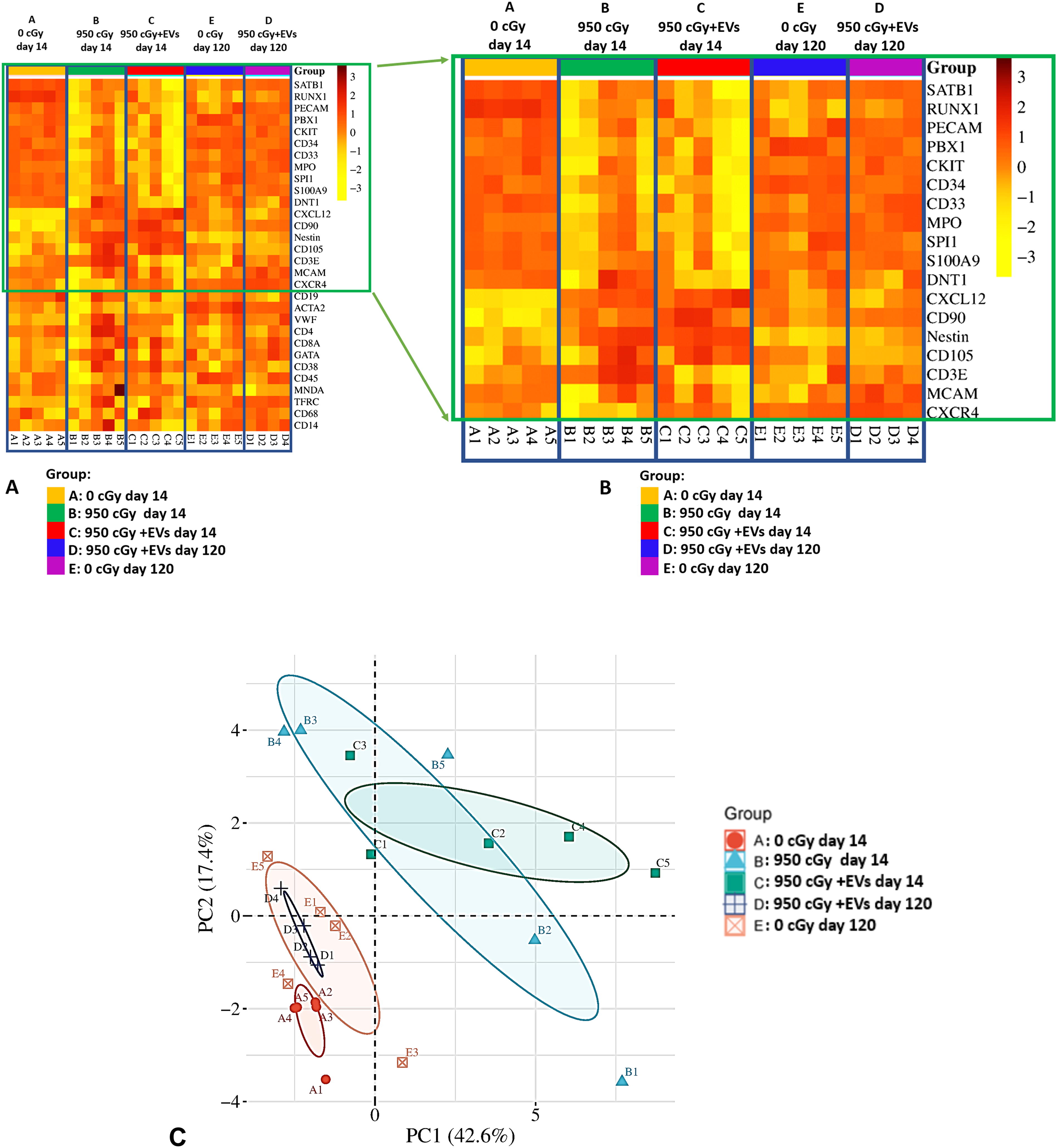
Effect of MSC-EV Treatment on Gene Expression in BMSCs from Irradiated Mice a
↑ and ↓ indicate gene upregulation and downregulation, respectively, defined as log2FC ≥ 1 or log2FC ≤ −1 with a P value of <0.05 (t-test) in two-group comparisons. N = 5 mice per group, except for the 950 cGy + EVs day 120 group (n = 4).
Discussion
In our previous studies, we assessed the therapeutic capacity of human MSC-EVs on the reversal of radiation injury in C57BL/6J mice exposed to mild or moderate TBI. 20 We observed that there was a significant but modest WBC and granulocyte count restoration at 3 weeks postirradiation, albeit not completely. Similarly, the injection of MSC-EVs also significantly increased engrafting stem cell capacity in the marrow from irradiated mice compared with the irradiated mice treated with vehicle control. In addition, we found that the MSC-EV population, containing both microvesicles and exosomes, was better at reversing radiation-induced injury than either population alone. 20 We also evaluated the biodistribution of MSC-EVs and their potential homing to the injury site after intravenous injection into C57BL/6J mice exposed to TBI. We observed that MSC-EVs were increasingly distributed in injured hematopoietic tissues such as the spleen and marrow after radiation. 21 In this study, we further evaluated whether human MSC-EVs could reverse marrow stem cell damage and improve the survival of mice exposed to high or lethal doses of radiation in an in vivo mouse model.
Our data showed that treatment with human MSC-EVs significantly improved the survival of mice exposed to high-dose TBI. MSC-EV treated mice maintained a 70% survival rate at 120 days postirradiation. However, the untreated mice died between 12–27 days postirradiation. Moreover, MSC-EV treatment significantly improved peripheral blood cell recovery in mice with 950 cGy TBI 3 months post-treatment. However, MSC-EV treatment did not result in immediate restoration of peripheral blood counts within the first 14 days in mice subjected to 700 and 950 cGy TBI. These findings align with our previous works and others, which showed that MSC-EVs or epidermal growth factor enhanced peripheral blood recovery approximately 3 weeks postirradiation but not within the first 2 weeks. 20,24 Similarly, Satyamitra M, et al.’s study showed that G-CSF treatment did not increase peripheral blood recovery between the vehicle- and G-CSF-treated groups until 14 days postirradiation. 25 This suggests that the effects of MSC-EVs on peripheral blood cell recovery require a longer duration, and the beneficial effects of MSC-EVs may not be immediate but instead primarily support long-term hematopoietic recovery. The lack of early recovery may be due to the severe radiation toxicity, leading to the loss of hematopoietic stem cells and differentiation potency. 26 It requires a longer time for MSC-EVs to reverse these effects, facilitating the regeneration of hematopoietic stem cells and rebuilding the bone marrow microenvironment.
The competitive transplantation assays further support that MSC-EVs enhance the function of hematopoietic stem and progenitor cells. BMSCs from MSC-EV treated mice exposed to 700 cGy TBI showed significantly higher engraftment rates at 1 month post-transplantation than those from untreated mice. These findings indicate that MSC-EV treatment not only promotes survival but also enhances the functional capacity of bone marrow cells in mice exposed to high-dose whole-body irradiation, consistent with previous studies that show the therapeutic benefits of MSC-EVs in enhancing survival. 24,27
Our gene expression analyses found that radiation exposure resulted in significant alterations in bone marrow cell gene expression 14 days postirradiation compared with nonirradiated mice. However, MSC-EV treatment did not significantly restore these alterations, except for the gene CD3E, which was upregulated by radiation but was restored to normal on day 14 post-EV treatment.
Significantly altered genes, including SATB1, RUNX1, PBX1, CKIT, CD34, CD33, and S100A9, were downregulated in the radiation alone and radiation + MSC-EV groups compared with the control group at 14 days postirradiation exposure, while CXCL12, CD90, and Nestin were upregulated. These genes play critical roles in hematopoietic stem cell maintenance and differentiation. SATB1 gene is crucial for the maintenance of HSC self-renewal, 28 RUNX1 gene is a key regulator of developmental hematopoiesis, 29 PBX1 gene regulates the balance between self-renewal and differentiation of hematopoietic stem cells, 30 CD34 is the most widely recognized marker for hematopoietic stem cells, 31,32 CKIT is associated with stem cell populations, 33,34 while CD33 is more relevant to myeloid differentiation, 35 and S100A9 gene promotes cellular senescence of bone marrow stromal cells via TLR4 and NLRP3 inflammasome formation. 36 The downregulation of SATB1, RUNX1, and PBX1 suggests impaired self-renewal capacity and altered developmental hematopoiesis, while the decrease in CKIT, CD34, and CD33 expression reflects a reduction in stem and progenitor cell populations, as well as changes in myeloid lineage commitment. In addition, we found upregulation of CXCL12, CD90, and Nestin gene expression after 14 days of radiation exposure. CXCL12 also known as stromal cell-derived factor 1 (SDF-1), is crucial for maintaining hematopoietic stem and progenitor cells (HSPCs) in a quiescent state and regulating cell proliferation, differentiation, and HSC mobilization, contributing to wound healing and tissue repair. 37 –42 It has been reported that the CXCL12 expression is increased after irradiation therapy and attracts CXCR4-expressing inflammatory cells that exacerbate acute normal tissue injury. 41 CD90 participates in MSC self-renewal and differentiation. The activation of CD90 expression has been observed in inflammation, wound healing, tumor development, and radiation. 43 –46 Nestin is expressed in a subset of bone marrow MSCs that contributes to HSCs maintenance, tissue growth, and regeneration. 47,48 Studies have shown that radiation induces Nestin expression in brain progenitor cells and may play a role in the remodeling and repair processes following brain irradiation injury, and radioresistance. 49 –51 As these genes are functionally associated with inflammation, tissue repair, and MSCs in supporting the hematopoietic niche, the upregulation of these three genes in bone marrow cells may be related to the response to radiation exposure.
CD3E, a component of the T-cell receptor complex essential for T-cell development and activation, 52,53 was increased in response to radiation but returned to normal levels with MSC-EV treatment by day 14. The initial upregulation of CD3E likely reflects inflammation and immune response following radiation exposure. Excessive inflammation and immune activation can exacerbate tissue damage in the bone marrow after radiation exposure. The restoration of CD3E expression after MSC-EV treatment indicated that MSC-EVs play an immunomodulatory role, helping regulate the immune response by reducing inflammation and preventing further damage. This aligns with the known anti-inflammatory properties of MSC-EVs, which have been shown to modulate immune responses and promote tissue repair by delivering anti-inflammatory microRNAs and cytokines. 54,55
By 120 days post-treatment, in MSC-EV-treated mice, we observed that the expression levels of genes altered at day 14 postirradiation had returned to levels similar to those of nonirradiated controls. Only two genes, MCAM and CXCR4, were upregulated at 120 days post-treatment compared with the nonirradiated controls, while the others showed no significant differences. CXCR4 is a chemokine receptor essential for the homing, maintenance, and retention of HSPCs within the bone marrow microenvironment. Studies have shown that CXCR4 supports HSPC quiescence and promotes bone marrow regeneration during radiation-induced damages. Overexpression of CXCR4 increases the migration ability of stem cells and enhances their therapeutic efficacy in damaged tissues. 56 –58 MCAM (CD146) is associated with cell adhesion, inflammation response and MSC cells. 59,60 CD146+ MSCs have been demonstrated to possess various unique properties in regenerative medicine, including homing ability, cell proliferation, and immunomodulation. Our data suggest that MSC-EVs may facilitate long-term recovery of bone marrow homeostasis by promoting the restoration of normal gene expression patterns in hematopoietic stem and progenitor cells.
Conclusion
In summary, our data indicated that MSC-EV treatment significantly extended the survival of mice exposed to high-dose radiation and helped recover the peripheral blood cell counts, gene expression, and function of bone marrow cells recovered from radiation injury. This highlights the potential of MSC-EVs as a therapeutic option for ARS, particularly under high-dose radiation exposure, where the current treatment options are limited. However, our study had several limitations. We observed significant improvements in survival and long-term hematopoietic recovery with MSC-EV treatment, but the underlying mechanisms remain unclear. Future studies should focus on identifying the specific components of MSC-EVs responsible for their radioprotective properties. In addition, we did not observe significant restoration of peripheral blood counts or gene expression at early time points postirradiation. This limitation may reduce the immediate therapeutic potential of MSC-EVs during the acute phase of radiation injury, particularly after high-dose TBI. Combining MSC-EV therapy with mitigation agents, such as G-CSF, may enhance overall therapeutic efficacy.
Footnotes
Acknowledgments
The authors acknowledge the use of services from the Extracellular Vesicle Core at the COBRE Center for Stem Cells and Aging, Rhode Island Hospital.
Data Availability Statement
The data associated with this study are available from the corresponding author upon reasonable request.
Declaration of Generative AI Use in Scientific Writing
During the preparation of the article, the authors used ChatGPT-4 to improve the readability. Following its use, the authors reviewed and edited the content as needed and take full responsibility for the final article.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was supported by NIH/NIGMS (1P30GM145500 and 1P20GM119943) and the Academic Assessment Research Award from Brown Physicians Incorporated.