
Abstract
Mesenchymal stem/stromal cells (MSCs) possess immunoregulatory capacity, which is enhanced in an inflammatory environment. Participation of extracellular vesicles (EVs) in this function is proposed, as they can transport various immunoregulatory molecules. However, the impact of the inflammatory microenvironment on the load of the different types of EVs released by these cells is not fully known. Therefore, this work analyzes in detail the temporal effect of IFN-γ, alone or in combination with TNF-α (TNF-α + IFN-γ), on the cargo of immunoregulatory molecules (programmed cell death ligand 1 [PD-L1], CD73, and intercellular adhesion molecule 1 [ICAM-1]) in large extracellular vesicles (L-EVs) released by human bone marrow mesenchymal stem cells (BM-MSCs). The presence of these molecules on the surface of L-EVs was determined by flow cytometry. Our results demonstrate that exposing BM-MSCs to TNF-α + IFN-γ for 24 h increases the percentage of PD-L1+ and CD73+ L-EVs. However, if this stimulus persists, the release of L-EVs with an immunoregulatory phenotype (PD-L1+, CD73+, and PD-L1+CD73+) decreases. The impact of pro-inflammatory cytokines on the transport of ICAM-1 by L-EVs is late, since up to 72 h of treatment with IFN-γ or TNF-α + IFN-γ, the percentage of ICAM-1+ L-EVs increases. In contrast, stimulation with IFN-γ for 72 h favors the release of CD73high and ICAM-1high L-EVs, but this effect also decreases in the presence of TNF-α. Our study generates novel knowledge about the impact of the inflammatory microenvironment on the cargo composition of L-EVs released by BM-MSCs and demonstrates, for the first time, that the prolonged presence of TNF-α reduces the cargo of immunoregulatory molecules in these structures.
Introduction
Mesenchymal stem/stromal cells (MSCs) possess immunoregulatory capacity, which is enhanced in an inflammatory environment. Due to the importance of cell–cell contact in the mechanisms used by these cells to regulate immune cell function, 1 –4 the participation of extracellular vesicles (EVs) as mediators of this interaction has been proposed since they can transport several immunoregulatory molecules, 5,6 including programmed cell death ligand 1 (PD-L1), Cluster of Differentiation 73 (CD73), and intercellular adhesion molecule 1 (CD54/ICAM-1), 3,7 –9 which influence T lymphocytes’ activation, differentiation, and effector function; maturation of dendritic cells; and polarization of macrophages. 3,10
The International Society for Extracellular Vesicles has defined EVs as structures released by cells, bounded by a lipidic membrane, and incapable of replication. It proposes a nomenclature that enables a more comprehensive analysis of the scientific literature, which will be utilized in this work. Thus, small EVs (S-EVs) are considered to have a diameter of less than 200 nm, whereas large EVs (L-EVs) are deemed to be larger than 200 nm. 11
Bone marrow mesenchymal stem cells (BM-MSCs) have been reported to release PD-L1+ EVs. 6 In addition, PD-L1+ S-EVs have been identified in cultures of umbilical cord mesenchymal stem cells (UC-MSCs) and Wharton’s jelly mesenchymal stem cells (WJ-MSCs), and their ability to regulate the immune response has been demonstrated in in vitro assays and preclinical models. 12 –14 Likewise, it has been shown that umbilical cord blood-MSCs (UCB-MSCs) release CD73+ EVs capable of suppressing T cell proliferation. 15 In addition, CD73+ S-EVs released by MSCs from gingival tissue (TG-MSCs) promote polarization toward M2-type macrophages. 16,17
It has been reported that BM-MSCs stimulated with Interferon-gamma (IFN-γ) or a mixture of Tumour Necrosis Factor alpha (TNF-α) plus IFN-γ produce EVs, 6 S-EVs, 16 and ICAM-1-loaded L-EVs. 18 However, to date, no study has analyzed the temporal effect of these inflammatory stimuli on the PD-L1, CD73, and ICAM-1 load on L-EVs released by these cells. Our work generates knowledge about the role of the inflammatory microenvironment in determining the cargo selection of L-EVs released by BM-MSCs. This knowledge is essential to deepen the understanding of the physiology of MSCs and the role of their EVs in the control and resolution of the immune response. In addition to the above, our study also contributes to improving in vitro conditioning protocols to activate MSCs and increase the expression of immunoregulatory molecules, without affecting their physiology.
Materials and Methods
BM-MSC culture and in vitro stimulation
From previously cryopreserved and characterized BM-MSC samples (n = 3), cultures were established and maintained under the previously reported growth conditions. 18 When the cultures reached 90% confluence, the monolayer was washed with phosphate buffered saline (PBS), and fresh medium supplemented with 10% fetal bovine serum (FBS) previously filtered through a 0.2 μm membrane was added. The cells were maintained under one of the following conditions: (1) 5, 10, 20, or 90 ng/mL IFN-γ (Peprotech) for 24, 48, and 72 h; (2) 1.0 ng/mL TNF-α (Peprotech) for 72 h; (3) 10 ng/mL IFN-γ alone or in combination with 1.0 ng/mL TNF-α for 24, 48, and 72 h; and (4) 5 or 90 ng/mL IFN-γ alone or in combination with 1.0 ng/mL TNF-α for 72 h. Cells maintained in culture for the same periods were considered as controls. Finally, the cells were processed by flow cytometry (as described below), whereas the supernatants were used to obtain L-EVs.
L-EV isolation
L-EVs were obtained from 8 mL of supernatant from each experimental condition. Based on the previously reported methodology, 18,19 the supernatants were centrifuged at 500g for 15 min, at 2,000g for 20 min, and at 17,000g for 60 min, at 4°C. The pellets from the last centrifugation were washed and resuspended in 200 µL of PBS for further characterization.
L-EV characterization
Transmission electron microscopy
The morphology of L-EVs was determined using transmission electron microscopy (TEM). Ten microliters of the sample was placed on a copper grid covered with carbon, which was then incubated for 5 min. The grid was subsequently stained with 10 µL of 2% filtered uranyl acetate for 4 min. Excessive solution on the grid was carefully absorbed with filter paper at each step. The preparation was allowed to dry for 15 min and then examined with a JEOL JEM-1200 electron microscope.
Nanoparticle tracking analysis
The size distribution and concentration of nanoparticles were determined on a NanoSight NS300 with software version 3.4 (Malvern Instruments, UK). For this purpose, 1 mL of a 1:10 sample dilution in PBS was used. Each sample was analyzed in triplicate, and three videos, each 60 s long, were collected.
Flow cytometry
Cells
Cells amounting to 1 × 105 were resuspended in 100 μL of PBS with 3% FBS and 1 mM calcium disodium ethylenediaminetetraacetic acid (EDTA) and incubated for 20–30 min with Phycoerythrin (PE)-labeled monoclonal antibodies directed against PD-L1 or ICAM-1. Next, the cells were washed and fixed with FACS Lysing Solution (BD Biosciences). The samples were analyzed using a CytoFLEX LX cytometer (Beckman Coulter), and at least 10,000 events were collected.
L-EVs
The characterization of L-EVs was performed using flow cytometry with the violet side scatter (Violet SSC) configuration on a CytoFLEX LX cytometer (Beckman Coulter), as previously reported. 18 Briefly, the resolution capacity of the equipment was verified using flow cytometry-grade yellow fluorescent nanospheres with diameters ranging from 130 to 1330 nm (Spherotech, USA). To analyze the presence of HLA-I, CD105, CD90, CD73, PD-L1, and ICAM-1, L-EVs were incubated for 30 min at 4°C with monoclonal antibodies coupled to fluorescein isothiocyanate (FITC), PE, allophycocyanin (APC), or brilliant violet 421 (BV421); subsequently, the sample was washed and stored at 4°C for 24 h. 8 × 105 to 1 × 106 events were acquired.
Cell and L-EV data were analyzed using CytExpert 2.0 software, and the percentage of expression and the mean fluorescence intensity (MFI) were obtained.
Statistical analyses
Data are expressed as the mean and standard error of the mean. Statistical analyses were performed using GraphPad Prism 5 software. Comparisons between groups were obtained using the paired t-test or the Mann–Whitney U test. A P value of less than 0.05 was considered statistically significant.
Results
IFN-γ levels differentially affect PD-L1 expression in BM-MSCs
The effect of different concentrations of IFN-γ (5, 10, 20, and 90 ng/mL) on PD-L1 expression was analyzed. We observed that all IFN-γ concentrations increased the percentage of PD-L1+ cells and levels of this molecule at 24, 48, and 72 h (P < 0.05). The rate of PD-L1+ cells is similar in BM-MSCs treated with 5 or 90 ng/mL of IFN-γ at all times evaluated. Notably, treatment with 90 ng/mL for 24 h significantly increases the presence of PD-L1 on the surface of BM-MSCs (P = 0.05) compared with 5, 10, and 20 ng/mL. The same effect is observed at 48 h, although there is only a significant difference with 5 and 10 ng/mL; whereas at 72 h, it is only statistically different from 10 ng/mL (Fig. 1A, B).

IFN-γ levels differentially affect PD-L1 expression in BM-MSCs. BM-MSC cultures were treated with 5, 10, 20, or 90 ng/mL IFN-γ for 24, 48, and 72 h. The basal expression of PD-L1 in BM-MSCs not stimulated and cultured for the same time was used as the control.
TNF-α does not stimulate PD-L1 expression in BM-MSCs
We observe that TNF-α does not affect PD-L1 expression in BM-MSCs (Fig. 2). Furthermore, the results show no difference in the percentage of PD-L1+ cells or the MFI between cells stimulated only with IFN-γ and those stimulated with TNF-α + IFN-γ for 72 h. In contrast, all concentrations of IFN-γ (5, 10, and 90 ng/mL), in the absence or presence of TNF-α, significantly increase the percentage of PD-L1+ cells compared with the control. When analyzing the MFI, the increase in PD-L1 levels is more evident with high concentrations of IFN-γ (Fig. 2).
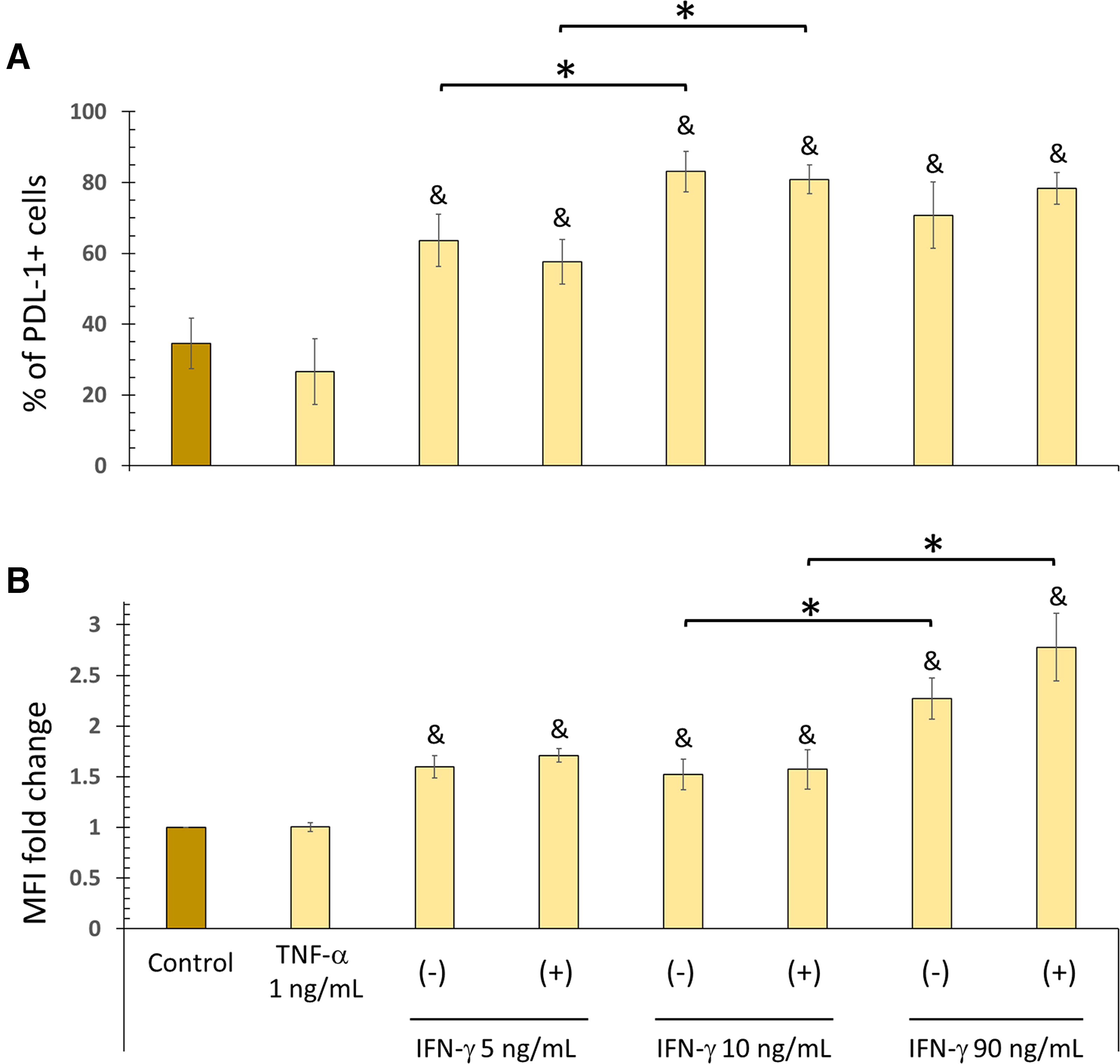
TNF-α does not affect PD-L1 expression in BM-MSCs. BM-MSCs were stimulated with TNF-α (1.0 ng/mL), IFN-γ (5, 10, or 90 ng/mL), or both for 72 h. The basal expression of PD-L1 in nonstimulated BM-MSCs cultured for 72 h served as the control.
To determine whether the absence of significant changes in PD-L1 expression by BM-MSCs could be due to the release of this molecule in L-EVs, the effect of IFN-γ or TNF-α + IFN-γ on the production of PD-L1+ L-EVs was analyzed.
Characterization of L-EVs by TEM and nanoparticle tracking analysis
Characterization of L-EVs by TEM reveals their rounded morphology, presence of a membrane, and a diameter exceeding 100 nm (Fig. 3A). Nanoparticle tracking analysis (NTA) revealed a size distribution with a mean diameter of 209 ± 83 nm. It was observed that 90% of the particles had a hydrodynamic diameter between 136 and 327.3 nm, whereas only 10% had a diameter smaller than 136.4 nm. In addition, a concentration of 3.40 × 108 ± 3.24 × 107 particles/mL was determined (Fig. 3B). Besides flow cytometry analysis (Fig. 3C) reveals that L-EVs exhibit characteristic markers of BM-MSCs on their surface, including the transmembrane proteins HLA-I and CD105, as well as the glycosylphosphatidylinositol (GPI)-anchored proteins CD90 and CD73 (Fig. 3D).
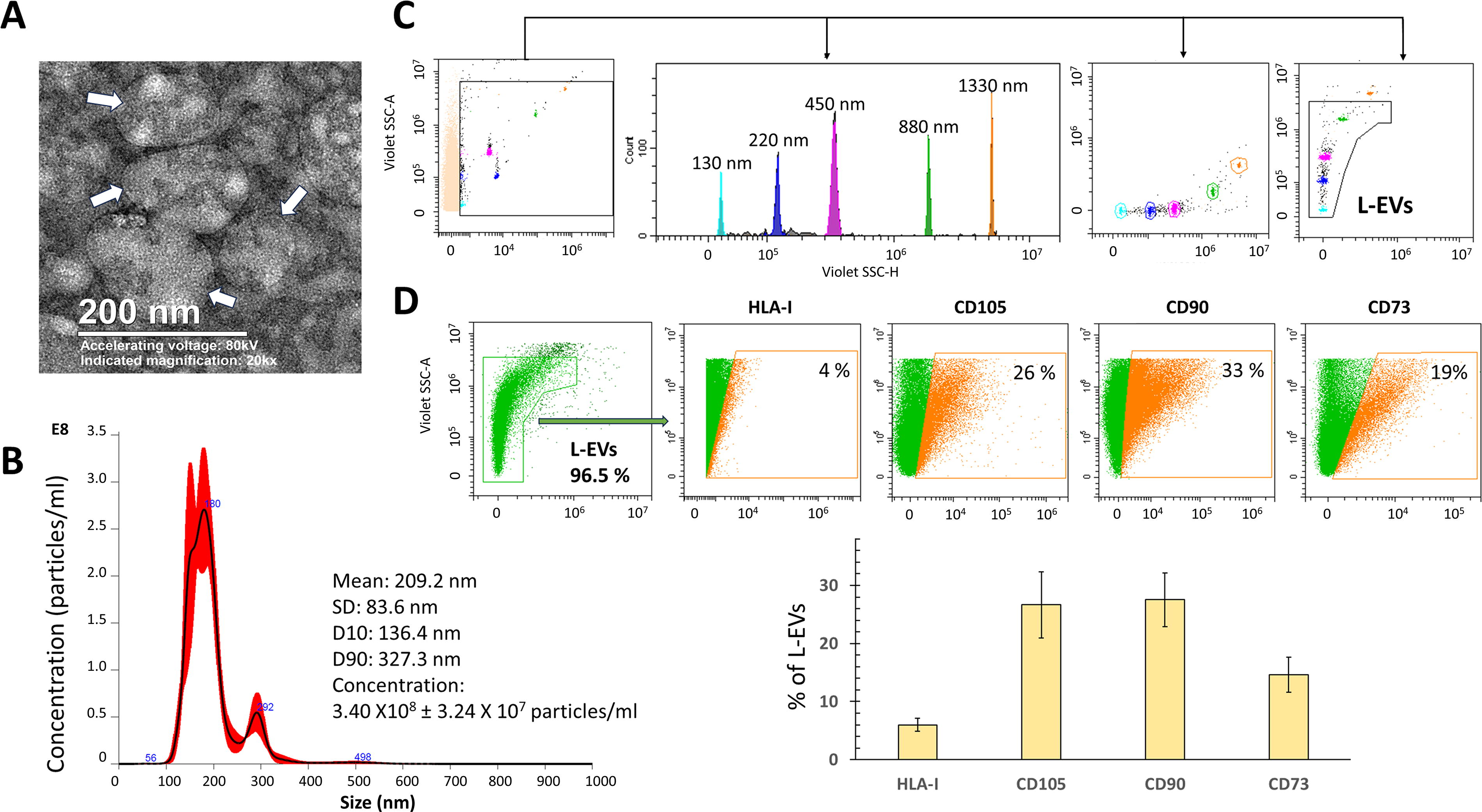
Characterization of L-EVs released by BM-MSCs. L-EVs were obtained from BM-MSC supernatants in the basal state and analyzed as described in the Materials and Methods section.
Inflammatory stimuli differentially affect the presence of PD-L1 and CD73 in L-EVs
BM-MSCs stimulated for 24 h with TNF-α + IFN-γ release a higher percentage of PD-L1+ L-EVs, and enrichment of this molecule is observed (trend) (Fig. 4A). Stimulation with IFN-γ increases the percentage of PD-L1+ L-EVs only at 72 h, without increasing PD-L1 levels. In contrast, BM-MSCs exposed to 90 ng/mL IFN-γ do not release a higher percentage of PD-L1+ L-EVs (Fig. 4B).
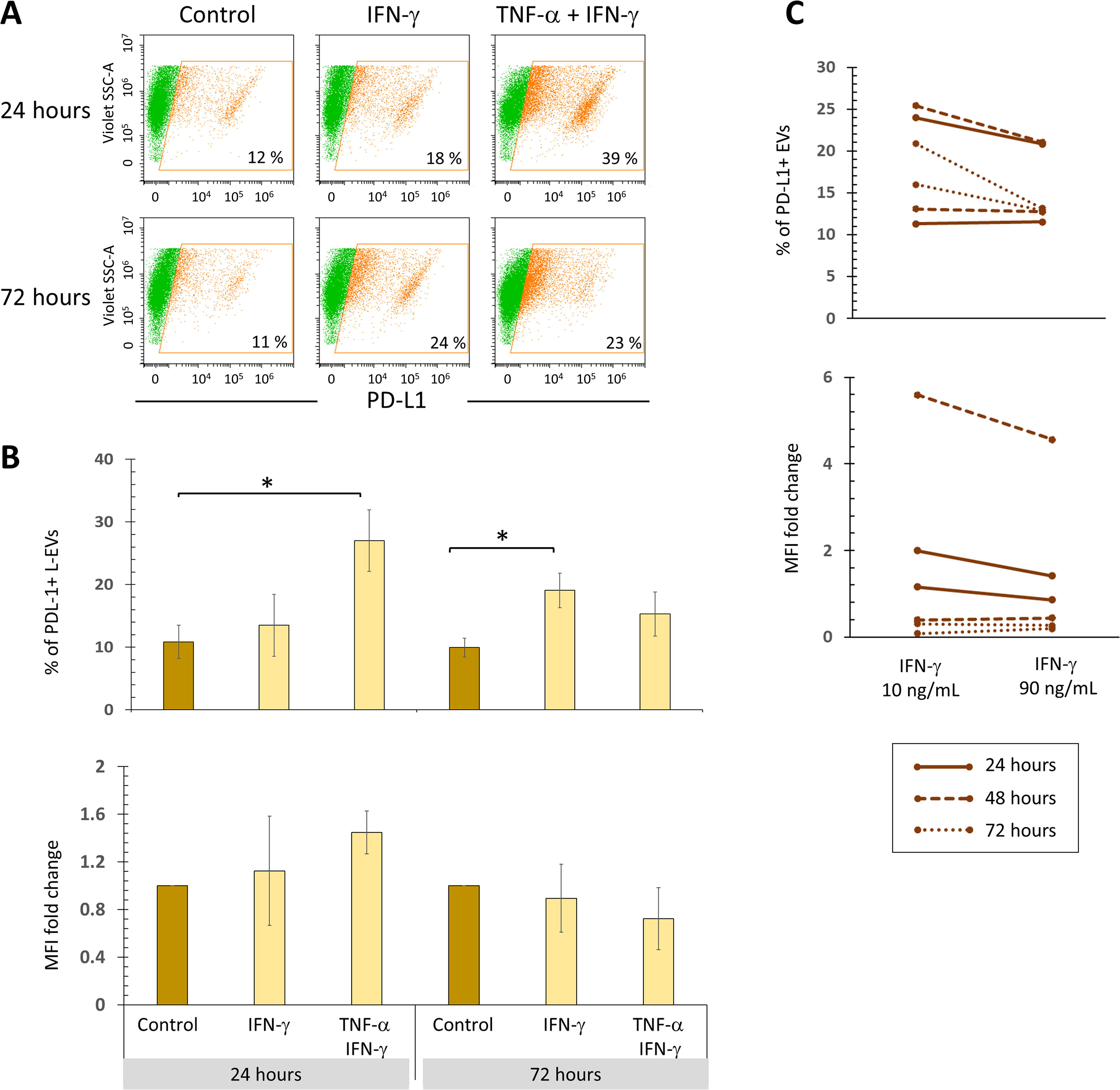
Simultaneous stimulation with TNF-α and IFN-γ increases the release of PD-L1+ L-EVs from 24 h of treatment. BM-MSC cultures were treated with 10 ng/mL IFN-γ for 24 and 72 h, in the absence or presence of TNF-α (1 ng/mL). The basal expression of PD-L1 in BM-MSCs not stimulated and cultured for the same time was used as the control.
Evaluating the presence of transmembrane and GPI-anchored proteins is a well-established guideline for analyzing EVs. 11 Therefore, in addition to detecting PD-L1 (a transmembrane protein), CD73 (a GPI-anchored protein) was also examined. Treatment with TNFα + IFN-γ for 24 h stimulates the release of CD73+ L-EVs, but without increasing the levels of this ectonucleotidase; this effect is lost after 72 h. In contrast, IFN-γ stimulation increases CD73 levels in L-EVs only after 72 h, but does not increase the percentage of CD73+ L-EVs (Fig. 5A, B).
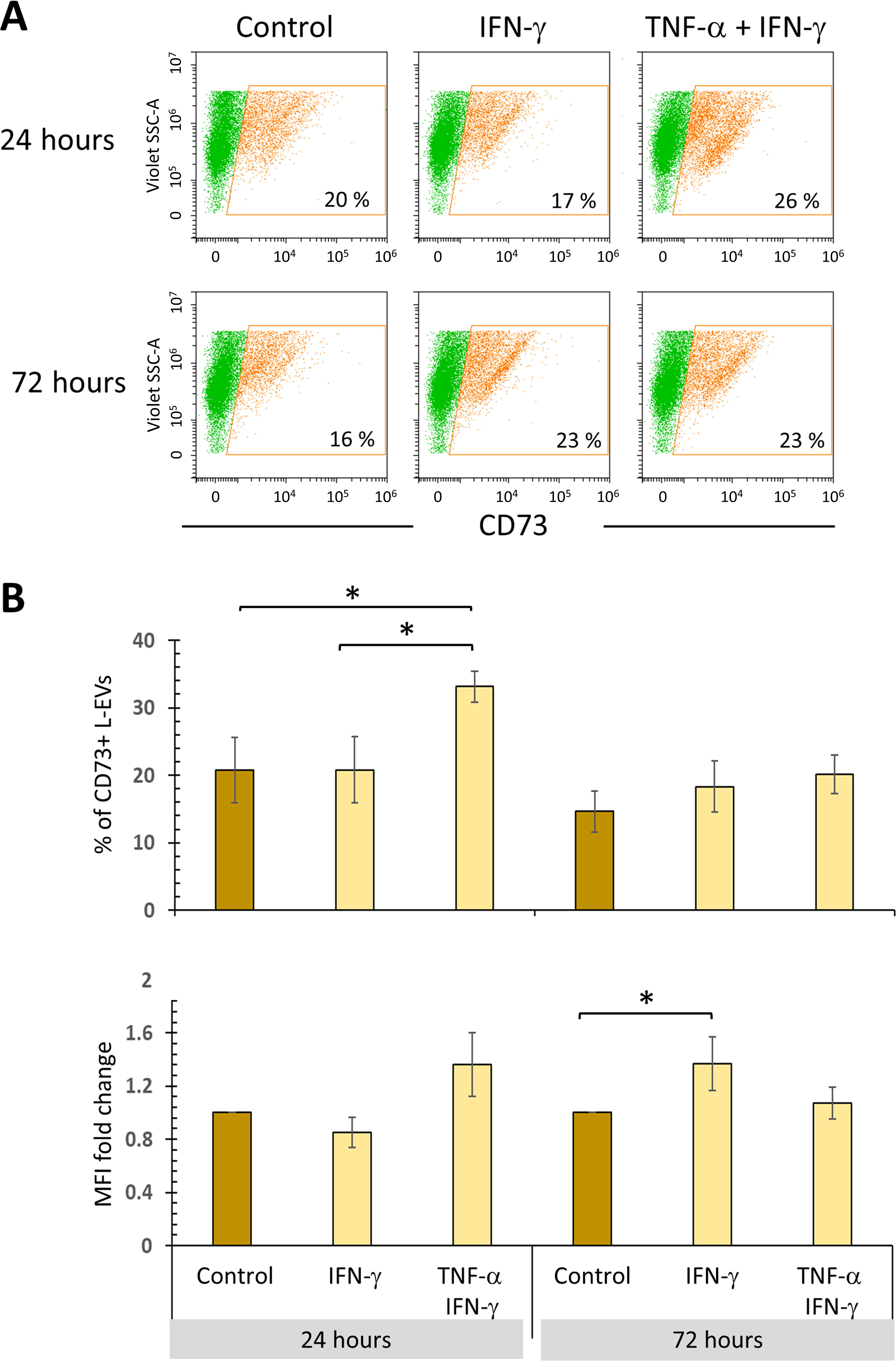
TNF-α and IFN-γ stimulate the release of CD73+ L-EVs. BM-MSC cultures were treated with 10 ng/mL IFN-γ for 24 and 72 h in the absence or presence of TNF-α (1 ng/mL). The basal expression of CD73 in L-EVs from BM-MSCs not stimulated and cultured for the same time was used as the control.
Treatment with TNF-α + IFN-γ, for a short time, stimulates the release of L-EVs with an immunoregulatory phenotype
It was observed that BM-MSCs exposed to both cytokines for 24 h released a higher percentage of PD-L1+CD73+ L-EVs. The same effect is observed with IFN-γ alone, but only up to 72 h of treatment. Interestingly, stimulation for 72 h with TNF-α + IFN-γ significantly decreases the percentage of PD-L1+CD73+ L-EVs to levels similar to those of the control. These results confirm the adverse effect of a persistent TNF-α stimulus on the production of L-EVs with immunoregulatory phenotype (Fig. 6).
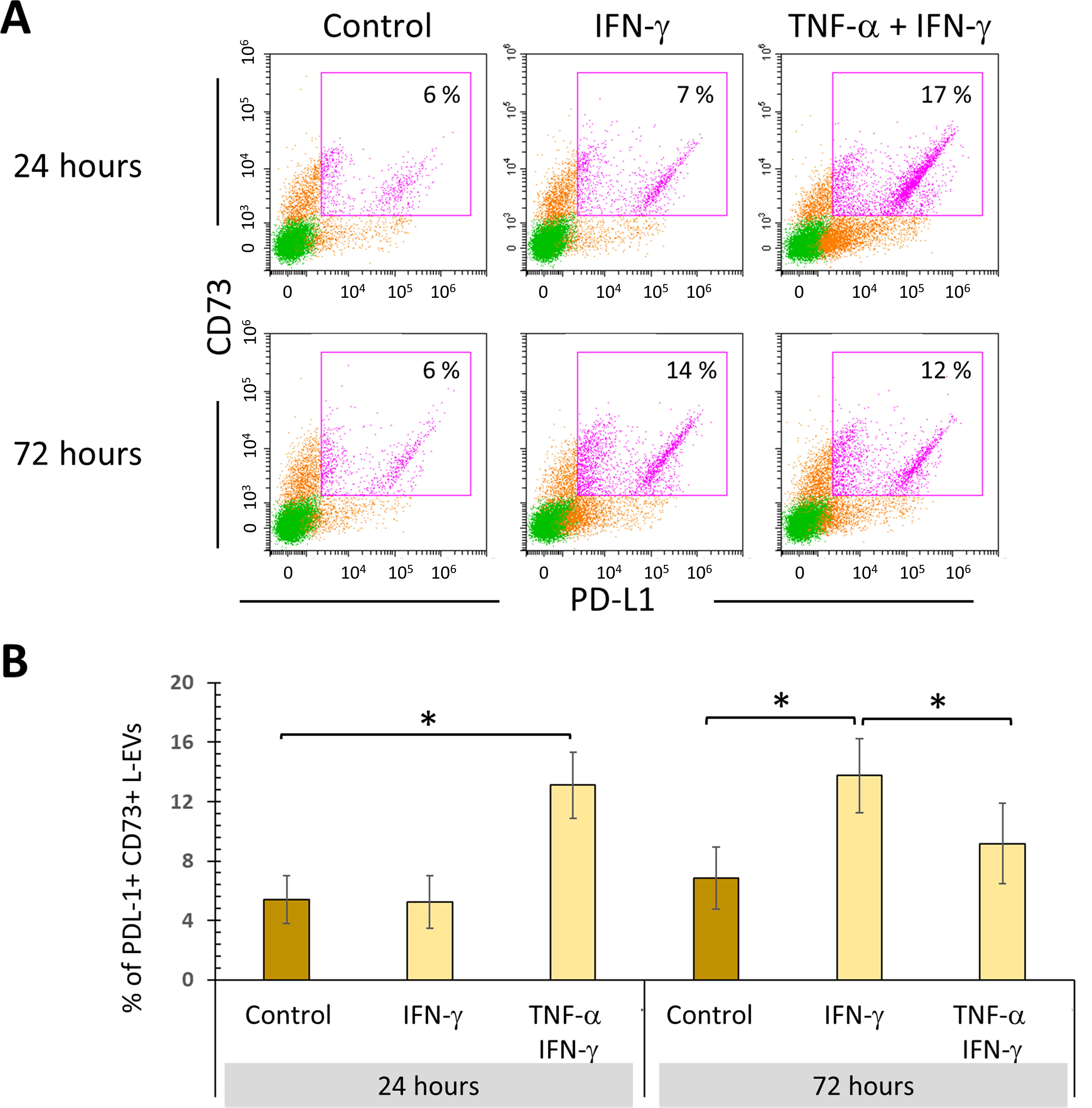
Persistent stimulation of BM-MSCs with TNF-α + IFN-γ affects the release of L-EVs with an immunoregulatory phenotype. BM-MSC cultures were treated with 10 ng/mL IFN-γ for 24 and 72 h in the absence or presence of TNF-α (1 ng/mL). The basal expression of PD-L1 and CD73 in L-EVs from BM-MSCs not stimulated and cultured for the same time was used as the control.
The impact of IFN-γ or TNF-α + IFN-γ on ICAM-1 cargo in L-EVs is delayed
We first determined that stimulation with IFN-γ alone or in combination with TNF-α significantly increases the percentage of ICAM-1 + BM-MSCs and the MFI at all analyzed times (Fig. 7A, B, respectively). It was also observed that during the first 48 h of treatment with one or both cytokines, there were no significant changes in the percentage of ICAM-1+ L-EVs. However, after 72 h of stimulation with IFN-γ, the percentage of ICAM-1+ L-EVs increases, and this effect is more pronounced when cells are exposed to TNF-α + IFN-γ. In addition, the enrichment of ICAM-1 in L-EVs was observed only up to 72 h of treatment and was statistically significant only in BM-MSCs treated with IFN-γ (Fig. 8A, B).
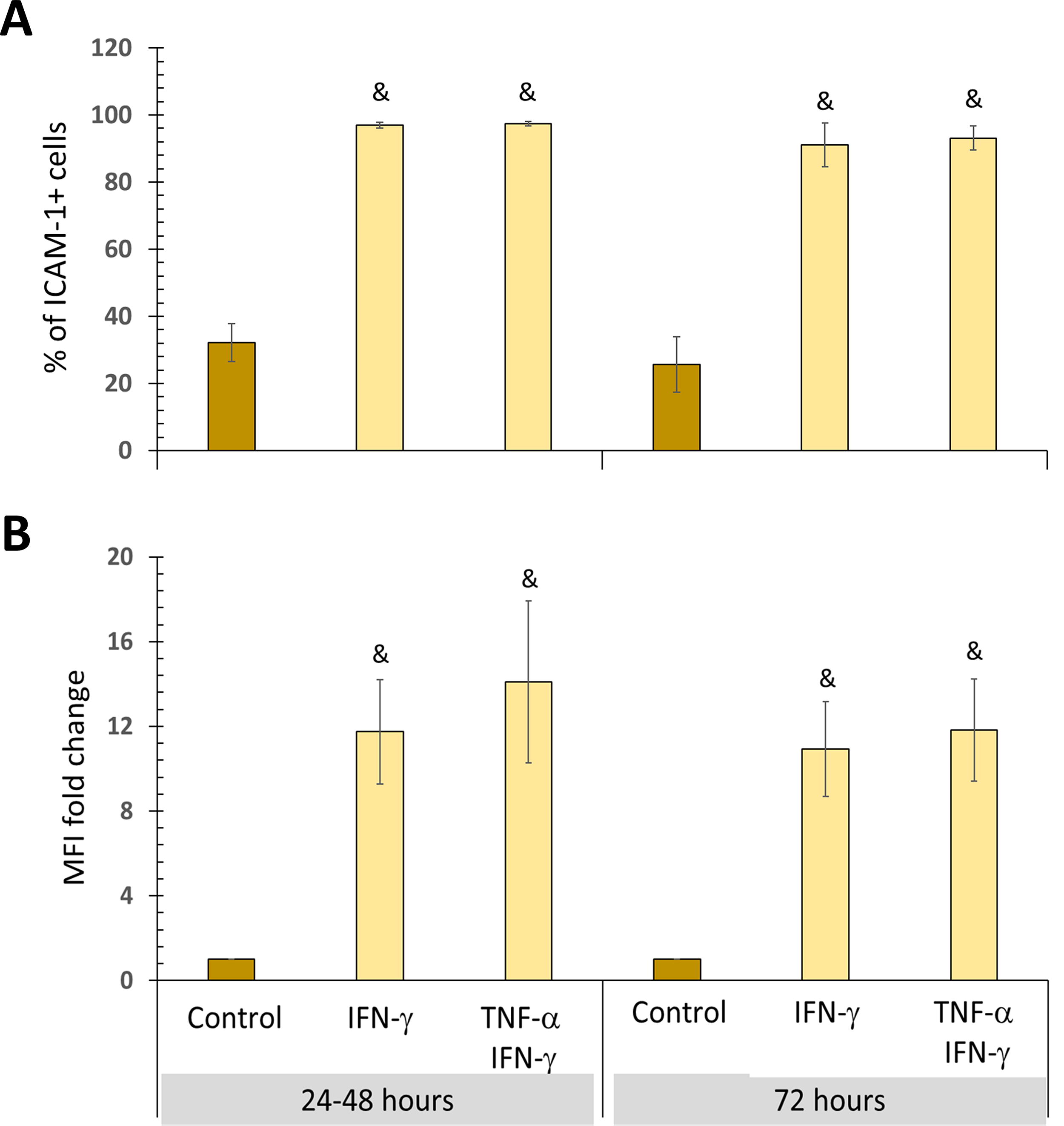
All inflammatory stimuli increase ICAM-1 expression in BM-MSCs. BM-MSC cultures were treated with 10 ng/mL IFN-γ, with or without 1.0 ng/mL TNF-α, for 24, 48, and 72 h. The basal expression of ICAM-1 in BM-MSCs not stimulated and cultured for the same time was used as the control.

The impact of IFN-γ or TNF-α + IFN-γ on ICAM-1 cargo in L-EVs is delayed. L-EVs were obtained from unstimulated BM-MSC supernatants (control), stimulated with IFN-γ, or treated with TNF-α + IFN-γ for 24 and 48 h (pooled data) and 72 h.
Discussion
The results of this study demonstrate the importance of the inflammatory microenvironment in regulating the expression of immunoregulatory molecules in MSCs and their loading into L-EVs. The time-dependent effect of cytokines on the loading of L-EVs with PD-L1, CD73, or ICAM-1 is possibly associated with the decrease and resolution of the immune response, essential in preventing chronic inflammation and maintaining tissue integrity (Fig. 9).

Hypothesis: Participation of L-EVs with immunoregulatory phenotype on T lymphocytes. During the first 24 h of stimulation with TNF-α and IFN-γ, the release of L-EV with immunoregulatory phenotype (PD-L1+ CD73+) increases, whose interaction with T lymphocytes could have the following effects: PD-L1 on the surface of L-EVs could interact with PD-1 on the surface of T cells, triggering an inhibitory signal that affects the activation of these cells, decreases their proliferation and cytokine secretion, and increases the differentiation of regulatory T lymphocytes.
4
Simultaneously, CD73 could promote the production of adenosine (ADO) in the extracellular space, which decreases T lymphocyte proliferation.
15,20
PD-L1 and CD73 are considered inhibitory checkpoints that promote the generation of an anti-inflammatory environment
Our results show that stimulation with low concentrations of IFN-γ for 24 h increases PD-L1 expression in human BM-MSCs, consistent with previous reports on murine BM-MSCs. This effect is more pronounced when using high concentrations of IFN-γ (90 ng/mL). Furthermore, our data show that TNF-α does not stimulate PD-L1 expression. Something similar has been observed in endothelial cells, 24 UC-MSCs, 25 and murine BM-MSCs. 26 In contrast, some studies report a synergistic effect between TNF-α and IFN-γ. 25,27 However, our results do not show synergy; these contradictory results are possibly due to the high cytokine concentrations used in some of these studies. 25,28
This study is the first to describe the influence of concentration, exposure time, and type of cytokine (TNF-α or IFN-γ) on the cargo of L-EVs released by human BM-MSCs. We observed that in the basal state, these cells produce a low percentage of PD-L1+ L-EVs; similar results have been reported in S-EVs released by UC-MSCs (exosomes). 12 Even in BM-MSCs in basal state, the absence of PD-L1 has been reported in S-VEs. 29 However, it has also been seen that S-EVs released by WJ-MSCs are enriched in PD-L1. 14 These contradictory results are due to the methods used to separate EVs of different sizes and the techniques used to analyze the molecules they transport. Western blot assays are generally performed to determine the increase in immunoregulatory molecules in EVs. However, this methodology does not allow observing differences between the increase in the percentage of EVs carrying the molecule of interest and the enrichment of the same in the EVs.
The method used in this work favors the enrichment of L-EVs; TEM and NTA verified this. In addition, the characterization of the phenotype in the CytoFLEX LX cytometer allows the selection of the population of L-EVs with a diameter between 130 and 1000 nm, whereas the S-EVs are discarded since the detection limit of the equipment is 130 nm. Therefore, the changes in the immunophenotype detected in this work are mainly due to the L-EVs present in the samples.
We did not observe significant differences in the levels of PD-L1 on BM-MSCs subjected to low concentrations of IFN-γ (5 and 10 ng/mL) in the absence or presence of TNF-α. This result appears to be related to a greater release of PD-L1+ L-EVs. This idea is supported by the results obtained when BM-MSCs are stimulated with high concentrations of IFN-γ (90 ng/mL), in which we observed a significant increase in the levels of PD-L1 on the cell surface. Still, the release of PD-L1+ L-EVs and the loading of this molecule on these structures did not increase. This behavior is possibly due to the high levels of IFN-γ, which can alter the physiology of the cells. 3,30 –32 The present study indicates, for the first time, that it can also modify the content and release of L-EVs.
Our results show that exposing BM-MSCs to TNF-α and IFN-γ for 24 h is sufficient to increase the release of PD-L1+ L-EVs, but not to increase the levels of this molecule. Similar results have been reported previously, in which the same levels of PD-L1 were found on EVs (60–400 nm) released by BM-MSCs at rest or activated. 6 Our study demonstrates, for the first time, that stimulation with both cytokines is essential to promote the release of PD-L1+ L-EVs. However, if this microenvironment persists, the production of L-EVs with these characteristics decreases. On the contrary, 72 h of stimulation with IFN-γ alone are required to increase the release of PD-L1 + L-EVs, but without PD-L1 enrichment. However, a recent study reports that UC-MSCs stimulated with TNF-α and IFN-γ for 48 h increase PD-L1 levels in S-EVs (120 nm). 13 In that report, the evaluation was performed using western blot, which does not allow for the identification of whether the observed increase is due to PD-L1 enrichment in EVs or to the increase in the percentage of PD-L1+ S-EVs.
We observed that none of the inflammatory stimuli increases the levels of PD-L1 on the surface of L-EVs; however, under certain conditions, the percentage of PD-L1+ L-EVs increases. This behavior is likely due to the crucial role of this molecule in regulating the immune response, particularly the activation of T lymphocytes (Fig. 9). Therefore, strategies have been developed to increase PD-L1 levels in EVs and achieve an immunosuppressive effect in inflammatory diseases 33 or to promote the acceptance of transplanted organs. 12,29 These valuable strategies should be carefully evaluated to identify and rule out any potential adverse effects since high levels of this molecule have been associated with the development and progression of cancer. 34 –37
TNF-α and IFN-γ have been shown to regulate PD-L1 expression in tumor cells at different levels. IFN-γ stimulates its transcription, whereas TNF-α has a post-transcriptional effect, as it favors deubiquitination and stability. 38 These mechanisms may be present in our cultures, where IFN-γ would stimulate the expression of PD-L1 mRNA and TNF-α would favor its stability in the membrane, resulting in an increased release of PD-L1+ L-EV from 24 h.
The analysis of other immunoregulatory molecules, including one GPI-coupled (CD73) and another transmembrane (ICAM-1), suggests that the effect of different inflammatory stimuli on the loading of L-EVs with PD-L1 is specific. We observed that BM-MSCs release a low amount of CD73+ L-EVs in the basal state. Similar results have been obtained with S-EVs from UC-MSCs. 39 In contrast, it has been reported that UCB-MSCs release a large percentage of CD73+ EVs. 15 At the same time, our results show that 72 h of IFN-γ stimulation are required for BM-MSCs to release CD73high L-EVs.
This study is the first to demonstrate that treating human BM-MSCs with TNF-α + IFN-γ for 24 h is sufficient to stimulate the release of PD-L1+CD73+ L-EVs; however, this effect decreases if the stimulus persists (Fig. 9). In contrast, studies carried out with TG-MSCs exposed to TNF-α, IFN-γ, and TNF-α/IFN-γ increased CD73 levels in S-EVs up to 48 h. 17 It has been reported that TNF-α is capable of stimulating the release of CD73+ exosomes by TG-MSCs at 24 and 48 h, which favors the polarization of macrophages toward an M2 phenotype. 16 It is important to highlight that, in these studies, cytokines were used at high concentrations.
During the first 48 h, both cytokines are essential for BM-MSCs to release L-EVs with an immunoregulatory phenotype (PD-L1+ CD73+). After this time, IFN-γ would be the main cytokine responsible. L-EVs with immunoregulatory phenotypes could participate in the regulation of the immune response from the beginning, and several pieces of evidence support such a hypothesis. It has been shown that T lymphocytes increase ATP production a few minutes after their activation, which induces a pro-inflammatory signal. However, in activated T lymphocytes, the expression of CD39 also increases and, therefore, the production of AMP, from which the action of CD73 produces adenosine (ADO). Thus, the activity of CD39 and CD73 is necessary to increase the production of ADO and decrease T lymphocyte proliferation, which favors the generation of an anti-inflammatory environment (Fig. 9). 15,20
Contrary to what has been observed with PD-L1 and CD73, our data show that the impact of IFN-γ or TNF-α + IFN-γ on ICAM-1 cargo in L-EVs is delayed, as 72 h of stimulation are required. Furthermore, IFN-γ alone can promote the enrichment of this adhesion molecule in L-EVs; however, this effect is diminished in the presence of TNF-α. Previously, we demonstrated that BM-MSCs stimulated with IFN-γ release L-EVs (microvesicles) enriched in ICAM-1 (MV-ICAM-1high). 18 Furthermore, a proteomic study of S-EVs released by UC-MSCs treated with TNF-α and IFN-γ also reports increased ICAM-1. 13
The delay in the production of ICAM-1high L-EVs may be relevant to the resolution of inflammation. The release of ICAM-1-enriched EVs at the onset of an inflammatory process could decrease the interaction of immune cells with the endothelium and their extravasation into areas of infection or damage. 40 In this regard, it has been proposed that soluble ICAM-1 can inhibit the interaction between lymphocyte function-associated antigen-1 (LFA-1) and membrane ICAM-1. 41 In addition, during the first 24 h of T lymphocyte activation, the interaction between ICAM-1 and LFA-1 is essential for increasing cytokine secretion and promoting proliferation. 42 This suggests that, after 72 h, the participation of ICAM-1+ L-EVs in regulating the immune response would be relevant. These structures could compete for their receptor, LFA-1, in various immune and endothelial cells, thereby contributing to the resolution of the inflammatory process (Fig. 9). This hypothesis is supported by several evidences that highlight the role of ICAM-1 in the resolution of inflammation and wound healing. 23
Our study demonstrates for the first time that the prolonged presence of TNF-α reduces the cargo of immunoregulatory molecules in L-EVs. The unfavorable effect of persistent stimulation with TNF-α on CD73 mRNA expression 17 and its increase in the apoptosis of TG-MSCs 43 have been noted. In addition, the persistence of high TNF-α levels is detrimental to regulating inflammation, which has been linked to the development of inflammatory and autoimmune diseases. Even this cytokine is considered a therapeutic target. 21,22,44
Conclusion
This work demonstrates that stimulating human BM-MSCs with TNF-α + IFN-γ for a short time promotes the early release of L-EVs carrying two critical molecules in regulating the immune response (PD-L1 and CD73); however, if this stimulus persists, the production of L-EVs with this immunoregulatory phenotype decreases. In contrast, a longer exposure time to IFN-γ is necessary to increase the release of CD73high and ICAM-1high L-EVs, and this effect is also decreased if TNF-α is present. These results contribute to understanding the influence of the microenvironment on the load of immunoregulatory molecules in L-EVs released by BM-MSCs. Further in vitro and in vivo studies are required to determine the biological impact of these L-EVs on immune cells and their contribution to the control and resolution of inflammation. Furthermore, our study generates knowledge that could improve in vitro conditioning protocols to obtain L-EVs with desired characteristics.
Footnotes
Acknowledgments
The authors gratefully acknowledge the MSc. Ignacio Martínez for his support in reviewing and translating the article. The authors also thank Carlos Villanueva García, Kouthumi Mondragon Figueroa, Sandra Salazar Aguilar, Lourdes Mora García, Jorge Hernández Montes, and Carlos Paredes Monsalvo for their valuable technical assistance. As well as to Rodolfo Paredes Díaz and Ruth Rincón Heredia for their support in the TEM, and to Alejandra Espinoza Bonilla for NTA analyses.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This work was supported by the National Council of Humanities, Sciences, and Technologies (CONAHCYT) grant to Marta E Castro-Manrreza (No CF-2023-G-568), currently called Secretary of Science, Humanities, Technology and Innovation (Secihti).