
Abstract
Abstract
Background:
Secretory immunoglobulin A (sIgA) and immunoglobulin G (IgG) are the principal immunoglobulins in the respiratory tract. Under normal circumstances, the upper respiratory tract contains predominantly sIgA, whereas IgG is of primary importance in the lower tract. Unlike other antibody isotypes, IgA antibodies participate in host defense functions without inciting inflammatory processes that might cause collateral damage to tissues. However, the ability of sIgA to modulate inflammatory reactions induced by other humoral factors is unclear. We examined the effect of the sequence of exposure to the two immunoglobulin isotypes on bacteria- or lipopolysaccharide (LPS)-mediated cytokine production by monocyte-polymorphonuclear neutrophil (PMN) cells in vitro.
Methods:
Blood monocytes were co-cultured with Escherichia coli or LPS. Either sIgA or IgG was added to subsets of cultures, which were incubated for 1 h at 37°C. Culture supernatant liquids were then co-cultured with naïve PMNs for 1 h at 37°C. Either IgG was added during this co-culture step or sIgA was added if IgG was added first. Cytokines were quantitated by enzyme-linked immunosorbent assay.
Results:
Significant increases in interleukin (IL)-6, tumor necrosis factor (TNF)-α, and IL-8 were noted after E. coli or LPS co-culture with monocytes and subsequent PMN challenge. Marked decreases in these pro-inflammatory cytokines were seen after the addition of sIgA or sIgA–IgG but not after IgG alone. This effect was more apparent with the immunoglobulin sequence IgA followed by IgG.
Conclusion:
The sequence of immunoglobulin isotype involvement in infectious processes is important in modulating the cytokine response to bacteria and LPS by inflammatory cells. Our in vitro results support the critical role of sIgA in the proximal airways in mitigating inflammatory responses by other humoral defenses in the distal airways. Loss of effective sIgA function may contribute to increased morbidity and deaths from nosocomial pneumonia in compromised patients.
Humoral defense is provided by immunoglobulin (Ig) A and IgG in mucosal secretions. The IgA in respiratory and other mucosal secretions is mainly of the secretory form (sIgA) [4–6]. It is synthesized by plasma cells in the lamina propria of mucosal cells in a dimeric form in association with a single J chain. The resultant IgA is transported to the basolateral surface of the overlying epithelial cells, where it binds to the polymeric immunoglobulin receptor (pIgR). This complex is transported to the apical surface of the epithelial cell and cleaved to yield sIgA. In human beings, 56–67% of bronchial immunocytes are committed to produce IgA and 12–16% to synthesize IgG [4]. Unlike sIgA, IgG appears in humoral secretions either by passive transudation or possibly from injury to the epithelial airway.
There are several important differences in the distribution and function of sIgA and IgG in the airways [6]. Upper-tract secretions contain predominantly sIgA with lesser amounts of IgG. In contrast, IgG plays a more prominent role in the lower respiratory tract. Approximately 19% of the total protein in bronchial lavage fluid is IgG and 10% is sIgA [4]. Another important difference is the relatively non-inflammatory nature of sIgA. It has limited capacity for complement activation and can downregulate the oxidative burst and release of inflammatory cytokines (e.g., tumor necrosis factor [TNF]-α and interleukin [IL]-6) by myeloid cells [7].
In a previous study using Calu-3 respiratory epithelial cells in an in vitro pneumonia model, the cytotoxic potential of polymorphonuclear neutrophils (PMNs) was decreased by co-culture with sIgA but not IgG [8]. These results were confirmed in a pneumonia model comparing conventional mice with pIgR-knockout mice (pIgR-/-) [9]. These mice lack the pIgR and therefore cannot transport IgA into mucosal secretions. They are otherwise immunologically intact, including having normal IgG bronchial concentrations. The pIgR-/- mice had significantly higher inflammatory cytokine concentrations in the broncheolar lavage fluid and serum, as well as greater PMN accumulation in lung tissue. These changes were associated with a nearly three-fold increase in the mortality rate. Previous animal and cell culture studies demonstrated that an absolute lack of sIgA in respiratory secretions may be detrimental after bacterial challenge.
However, patients with selective IgA deficiency have a highly variable susceptibility to infection. This may be secondary to compensatory increases in IgM in mucosal secretions and the fact that IgA deficiency may not be absolute in these patients. There are a number of other clinical conditions common in the critically ill where sIgA is present in respiratory and other secretions but in a decreased concentration [10–13]. Furthermore, the higher sIgA concentrations normally present in the proximal airways may modulate inflammatory responses in the distal airways, where IgG is more prevalent. In the current study, we examined the modulatory effect of the different immunoglobulin isotypes alone and in sequence on the cytokine responses to bacteria or lipopolysaccharide (LPS) by inflammatory cells.
Materials and Methods
Institutional approval
This study was approved by the Human Investigation Committee at Wayne State University.
Polymorphonuclear neutrophil isolation
Venous whole blood was collected from random healthy donors in vacuum tubes containing ethylene diamine tetraacetic acid (EDTA). The PMNs were isolated by incubating the whole blood with 6% dextran for 45 min at 4°C. The leukocyte-rich supernatant liquid was aspirated, layered on Histopaque 1077 (Sigma, St. Louis, Mo), and centrifuged at 1,300 rpm (400 × g) for 30 min at 4°C. A 25-sec red blood cell lysis was performed on the pellet and isotonicity restored by adding 2 mL of 3.4% NaCl. The cells were then diluted in phosphate-buffered saline (PBS) and centrifuged for 10 min at 1,300 rpm at 4°C. Supernatant liquids were discarded and the PMNs resuspended in PBS and washed. The pellet was gently resuspended in PBS at a concentration of 1 × 106 cells/mL and used immediately.
Monocyte isolation
Venous whole blood was collected from random healthy donors in vacuum tubes containing EDTA. Whole blood was layered in UNI-SEPmaxi Tubes (Novamed Ltd., Jerusalem, Israel) and centrifuged at 1,000 × g for 20 min at 18–20°C. The mononuclear layer was removed with the aid of a pipet and washed several times with PBS. Monocytes were resuspended in Roswell Park Memorial Institute (RPMI)-1640 medium + fetal bovine serum (FBS), added to 12-well tissue culture plates, and incubated for 2 h at 37°C. Plates were subsequently washed with PBS to remove any non-adherent cells. More RPMI-1640 + FBS was added, and the plates were allowed to incubate overnight at 37°C, making the monocytes ready to be used in the experiments.
Experimental design
Blood monocytes were co-cultured with LPS (10 micrograms/mL) or Escherichia coli (106 organisms/mL). Then 50 microgram/mL amounts of sIgA purified from human colostrum and IgG (Sigma Chemical, St. Louis, MO) were added, and the cultures were incubated for 1 h at 37°C. The sIgA purified from human colostrum was used because it is commercially available and contains percentages of the IgA1 and IgA2 subclasses similar to those found in respiratory secretions. After the incubation, supernatant liquids from monocytes exposed to LPS, E. coli, sIgA, IgG, or some combination were used to treat naïve PMNs. The PMNs were subsequently assayed for IL-6, TNF-α, and IL-8 production. Chemotaxis by naïve PMNs was assessed using the supernatant liquids as the chemoattractant. A detailed description of the methodology is shown in Fig. 1.

Experimental method. Abbreviations: IL = interleukin; TNF = tumor necrosis factor.
Cytokine analysis by enzyme-linked immunosorbent assay
Monocyte supernatant liquids incubated with E. coli or LPS in the presence or absence of either sIgA or IgG were co-cultured with naïve PMNs for 1 h at 37°C. The PMN supernatant liquids were collected and stored immediately at −70°C for subsequent cytokine analysis. The IL-6, IL-8, and TNF-α were quantitated using a solid-phase sandwich enzyme-linked immunosorbent assay (ELISA). These immunoassay kits are commercially available and were used according to the manufacturer's directions (Cytoscreen, Biosource International, Camerillo, CA). The minimum concentrations of IL-6, Il-8, and TNF-α detectable with these kits are 2 pg/mL for IL-6 and IL-8 and <1 pg/mL for TNF-α.
Chemotaxis assay
Chemotaxis of PMNs was assessed using the ChemoTx System purchased from NeueoProbe, Inc. (Gaithersburg, MD). Supernatant liquids (300 microliters) from the treated monocyte populations were placed in the bottom layer of the chemotaxis system. Purified PMNs were resuspended at 1 × 106/mL in RPMI-1640, and 50 microliters of this suspension was added to the top layer of the chemotaxis membrane and allowed to incubate for 2 h at 37°C. After incubation, PMN chemotaxis into the basal chamber was quantitated using CellTiter 96 Aqueous One Solution purchased from Promega Corp. (Madison, WI). A 20-microliter sample of the above reagent was pipetted into each well and incubated for 4 h at 37°C, after which absorbance was read at 490 nm using a 96-well plate reader.
Statistical analysis
All samples were compared using analysis of variance (ANOVA) with a post-hoc Tukey test. Statistical significance was inferred at p < 0.001. All data are expressed as mean ± standard deviation.
Results
The effect of the supernatant liquids from the different monocyte co-cultures on IL-6 production is shown in Figs. 2 and 3. Release of IL-6 was the greatest when PMNs were exposed to liquids from monocytes cultured with either E. coli or LPS (1387 ± 45 and 1237 ± 45 pg/mL, respectively). Similar results were noted after the addition of IgG, where IL-6 concentrations were similar to those seen with PMN exposure to E. coli or LPS alone. In contrast, sIgA alone or the sequential addition of sIgA followed by IgG led to more than a five-fold decrease in IL-6 production. However, the addition of IgG followed by sIgA had a much smaller effect on IL-6 production in both the E. coli and the LPS group.
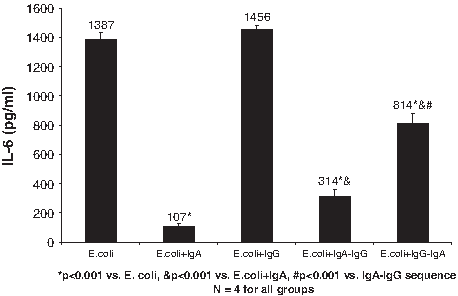
Effect of sequence of exposure to secretory immunoglobulin (Ig) A and IgG on Escherichia coli-mediated interleukin (IL)-6 production by naïve polymorphonuclear neutrophils (PMNs) treated with supernatant liquids from treated monocytes. Monocytes had been cultured with E. coli and sequential addition of sIgA–IgG or IgG–sIgA. The IL-6 concentration was measured in the PMN culture liquids by enzyme-linked immunosorbent assay.

Effect of sequence of exposure to secretory immunoglobulin (Ig) A and IgG on lipopolysaccharide (LPS)-mediated interleukin (IL)-6 production by polymorphonuclear neutrophils (PMNs) treated with supernatant liquids from treated monocytes. Monocytes had been co-cultured with LPS and sIgA and IgG separately or in sequence. The IL-6 concentration in the PMN culture liquid was measured by enzyme-linked immunosorbent assay.
The impact of the addition of the different antibody isotypes on TNF-α production in the cultures is shown in Figs. 4 and 5. The highest concentrations of TNF-α were noted in the E. coli or LPS alone groups (1721 ± 50 and 1632 ± 272 pg/mL, respectively). The addition of IgG had no effect on TNF-α, which was similar to what was seen with IL-6. On the other hand, the addition of sIgA or the sIgA–IgG antibody sequence led to profound decreases in TNF-α. With SIgA alone, TNF-α concentrations were decreased 100-fold for the E. coli and LPS groups. A significant but less profound effect was noted after addition of immunoglobulins in the sIgA–IgG sequence, where a more than three-fold reduction in TNF-α production was seen for both E. coli and LPS co-cultures. Addition of immunoglobulin in the IgG–sIgA sequence had a lesser effect on TNF-α, the reduction being only about two-fold for the E. coli and LPS groups.

Effect of sequence of exposure to secretory immunoglobulin (Ig) A and IgG on Escherichia coli-mediated tumor necrosis factor (TNF)-α production by naïve polymorphonuclear neutrophils (PMNs) treated with supernatant liquids from treated monocytes. Monocytes had been cultured with E. coli and sIgA and IgG alone or in sequence. The TNF-α concentration was measured in the PMN culture liquid by enzyme-linked immunosorbent assay.
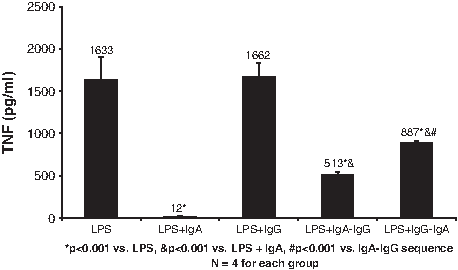
Effect of sequence of exposure to secretory immunoglobulin (Ig) A and IgG on lipopolysaccharide (LPS)-mediated tumor necrosis factor (TNF)-α production by naïve polymorphonuclear neutrophils (PMNs) treated with supernatant liquids from treated monocytes. Monocytes were exposed to LPS and sequential sIgA–IgG, IgG–sIgA, or the two antibody isotypes alone. The TNF-α concentration in the PMN culture liquid was measured by enzyme-linked immunosorbent assay.
The IL-8 data are shown in Figs. 6 and 7. The addition of E. coli or LPS produced the highest IL-8 concentrations (2750 ± 210 and 2528 ± 397 pg/mL, respectively). These results were similar to those with either E. coli or LPS and IgG. Similar to the results with IL-6 and TNF-α, IL-8 concentrations were lowest with either sIgA or the addition of immunoglobulins in the sequence sIgA–IgG. With sIgA alone, the concentrations of IL-8 were more than ten-fold lower for the E. coli and LPS preparations. Similarly, the sequence sIgA–IgG resulted in more than a two-fold reduction in IL-8 concentrations. A more modest reduction in IL-8 was noted after the addition of immunoglobulin in the IgG–sIgA sequence.

Effect of sequence of exposure to secretory immunoglobulin (Ig) A and IgG on Escherichia coli-mediated interleukin (IL)-8 production by naïve polymorphonuclear neutrophils (PMNs) treated with supernatant liquids from treated monocytes. The monocytes were co-cultured with E. coli and either sIgA or IgG alone or in combination. The IL-8 concentration was measured by enzyme-linked immunosorbent assay.

Effect of sequence of exposure to secretory immunoglobulin (Ig) A and IgG on lipopolysaccharide (LPS)-mediated interleukin (IL)-8 production by naive polymorphonuclear neutrophils (PMNs) treated with supernatant liquids from treated monocytes. The monocytes were co-cultured with LPS and sIgA and IgG alone or in combination. The IL-8 concentration was measured by enzyme-linked immunosorbent assay.
A chemotaxis chamber was used to index the potential for PMN accumulation in the lungs. Both sIgA alone and the immunoglobulin sequence sIgA–IgG caused profound decreases in PMN accumulation in the basal compartment of the chamber (Fig. 8). There was minimal effect of the IgG–sIgA immunoglobulin sequence compared with culture with E. coli alone.

Effect of supernatant liquids from monocytes co-cultured with Escherichia coli, secretory immunoglobulin (Ig) A, and IgG on chemotaxis of naive polymorphonuclear neutrophils (PMNs). Secretory IgA alone or the isotype sequence sIgA–IgG produced marked decreases in PMN chemotaxis, whereas the IgG–sIgA sequence had little effect.
Discussion
Nosocomial pneumonia remains a major cause of morbidity and death in the critically ill patient. Although prompt diagnosis and initiation of appropriate antibiotics are important in improving outcome, host defenses also are crucial in eradicating the pneumonic process.
Secretory IgA protects the respiratory tract by its direct antibody function and by interacting with other inflammatory cells [3]. The antibody functions of sIgA include prevention of micro-organism attachment to mucosal surfaces, mucus trapping, neutralization of certain bacterial enzymes, and inhibition of antigen penetration (immune exclusion). These functions are important in the prevention of tracheobronchitis and eventual pneumonia [14]. As most nosocomial pneumonias begin with upper airway colonization followed by bacterial entry into the lower respiratory tract, the higher concentrations of sIgA in the proximal airways would aid in “gatekeeping” [14,15]. However, numerous insults, including hemorrhagic shock, ethanol consumption, and less-than-ideal routes and types of nutrition, decrease respiratory sIgA concentrations and increase the pneumonia risk [12,13,16,17].
The host response to bacterial entry into the distal airways is initiated by the phagocytic function of resident alveolar macrophages. Neutrophils and blood monocytes are then recruited to help eradicate invading pathogens. Cytokine production by the effector cells is a key factor in the inflammatory response but must remain compartmentalized to prevent systemic spillover and collateral damage to the host [18,19].
Injury to the alveolar membrane by either bacterial toxins or an exaggerated inflammatory host response results in the release of pro-inflammatory mediators into the systemic circulation and possibly organ dysfunction and death [20–22]. The indirect effects of sIgA may be critical in controlling the host inflammatory response in this situation. Secretory IgA is recognized as a “non-inflammatory” antibody [3]. First, the gatekeeper function of sIgA at mucosal surfaces occurs without initiating an inflammatory response. Second, the indirect functions of sIgA are considered non-inflammatory: (1) sIgA does not activate complement by either the classic or the alternate pathway under physiological conditions; and (2) sIgA downregulates inflammatory responses by myeloid cells, including neutrophils and monocytes. In our study, sIgA decreased IL-6, TNF-α, and IL-8 production by neutrophils cultured in supernatant liquids from monocytes stimulated by either bacteria or LPS. No effect on cytokine production was noted after the addition of IgG to the cultures.
Both sIgA and IgG are present in variable amounts in respiratory secretions in vivo. Therefore, experiments were conducted with both sIgA and IgG added to the monocyte–bacteria or LPS preparations. Interleukin-6, TNF-α, and IL-8 production was lower than that achieved with either bacteria or LPS alone or with IgG. However, the concentrations of these cytokines still were higher than in the SIgA alone group. Because of the proximal-to-distal airway concentration gradient of the sIgA and IgG isotypes, we studied the impact on pro-inflammatory cytokine production according to the sequence of immunoglobulin administration. We found the SIgA–IgG sequence had a more profound effect on cytokine concentrations than did the IgG–SIgA sequence. The minimum concentration of sIgA necessary to obtain this effect was not determined and is a subject of current studies. Chemotaxis was used to index the potential for influx and accumulation of PMNs in lung tissue. Secretory IgA alone or combined with IgG decreased PMN chemotaxis, indicating lower potential for an exaggerated inflammatory response.
There is conflicting information in the literature regarding the anti-inflammatory properties of IgA [23,24]. The reasons for discrepancies include the type of IgA (serum, polymeric, or secretory), the myeloid cell lines employed, and the pathway used to stimulate monocytes/macrophages (ERK1/2 dependent vs. independent) [25]. Despite this, there is increasing recognition of the multiple mechanisms whereby sIgA protects mucosal surfaces against potential pathogens. The ability to protect respiratory and mucosal surfaces from micro-organisms in secretions is well described. Additionally, when all Ig isotypes and cell-mediated mechanisms are present, an important role of sIgA may be to mitigate the inflammatory side effects of the other immune effector cells.
There is little clinical information regarding sIgA concentrations on the respiratory and other mucosal surfaces in critically ill patients. Annane et al. demonstrated that bronchial secretion of IgA is blunted in ventilated patients with pneumonia [26]. There also is indirect evidence of the importance of “adequate” sIgA concentrations in bronchial and other mucosal secretions. The salutary effects of enteral rather than parenteral nutrition on the development of pneumonia and other septic complications in the intensive care unit has been attributed to preservation of mucosal immunity [27,28]. The results of the present study support the importance of sIgA in respiratory and other mucosal secretions in critically ill patients.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
*
Presented at the Twenty-ninth Annual Meeting of the Surgical Infection Society, Chicago, Illinois, May 6–9, 2009.