
Abstract
Abstract
Background:
Although the cause of intestinal necrosis in diseases such as necrotizing enterocolitis (NEC) is multi-factorial, oxidants and bacteria likely are key factors. Many animal models of NEC generate histologic necrosis, but it is rare to see the gross necrosis that is observed in infants. Here, we present a novel model that produces full-thickness intestinal necrosis when chloramine-T, an oxidizing agent, is introduced directly into isolated intestinal loops. The aim of this study was to determine the role of bacteria in this model.
Methods:
Three-week-old mice underwent isolated ileal loop construction by suture ligation, and either chloramine-T or saline was injected into the loop. Intestines were then returned to the abdominal cavity, and the incision was closed for 24 h, after which the intestinal loops were analyzed histologically and microbiologically. To determine if bacteria home to the site of injury, some mice had intracecal injection of luminescent Pseudomonas aeruginosa (PA). These mice were sacrificed 2, 6, 8, and 24 h later, and luminescent bacteria were localized via photon camera imaging.
Results:
No gross necrosis was observed in the saline-treated group, but 59% of the animals in the chloramine-T-treated group had necrosis (p<0.001). Relative bacterial species numbers were calculated for untreated control animals and those with saline-treated loops and chloramine-T-treated loops without and with necrosis. Lactobacillus was detected in 60% of the control animals but was absent in all animals that underwent surgery. Methicillin-sensitive Staphylococcus aureus, PA, and Enterococcus faecium were present only in animals that underwent loop construction; however, bacterial communities did not differ according to loop treatment or the presence or absence of necrosis. After intracecal injection of PA, bacteria homed to the loop and proximal bowel even though the loop was discontinuous with the remaining bowel.
Conclusions:
Intraluminal chloramine-T causes full-thickness necrosis in three-week-old mice and is characterized by a loss of probiotic bacteria such as Lactobacillus. Necrotic loops of bowel become colonized rapidly with pathogenic bacteria by unconventional mechanisms. Oxidant stress and colonization by pathogenic bacteria may play important roles in intestinal necrosis across a wide spectrum of pathologic states, including NEC.
Although NEC is most often associated with prematurity, adults can also develop full-thickness intestinal necrosis in a variety of circumstances in which the pathogenic mechanisms remain unclear. There is a major gap in the knowledge of the precise mechanisms by which intestinal epithelial dysfunction or inflammation that is initiated by a systemic disturbance or a microorganism leads to full-thickness necrosis. Clinically, this injury often occurs in a localized fashion without the development of systemic sepsis until there is perforation. New experimental models are needed to define the natural pathway from intestinal inflammation to full-thickness necrosis.
Here, we describe a novel murine model that allows investigation of the combined effects of bacteria and oxidants on the development of intestinal necrosis. We believe that this model will permit molecular interrogation to identify the specific microorganisms at the site of necrosis and determine how they cause transformation from inflammation to necrosis.
Materials and Methods
Animal model
All experiments were approved by the Animal Care and Use Committee at the University of Chicago (IACUC protocol 72201). Three-week-old male C57BL6 mice were used. Mice were housed with access to standard chow and sterile tap water ad libitum under 12 h light/dark cycles. They were allowed to acclimate for at least 72 h before surgery.
To determine whether intestinal necrosis could be generated by intraluminal injection of an oxidizing agent, mice underwent surgical construction of an isolated loop of ileum. They were anesthetized with a combination of ketamine 90 mg/kg and xylazine 5 mg/kg via intraperitoneal injection. The abdomen was prepared aseptically, and a midline laparotomy incision was created. Using sterile cotton swabs, the bowel was eviscerated to expose the insertion of the ileum into the cecum. A 4-0 silk suture was tied to occlude the small bowel immediately proximal to the ileocecal junction, and a second suture was tied to occlude the bowel 4 cm proximal to the ileocecal junction, creating an isolated loop of ileum. Using a 1-mL syringe and a 27-gauge needle, 200 mcL of 0.05% chloramine-T or sterile saline was injected into the lumen of the isolated loop immediately distal to the most proximal suture. Chloramine-T, a derivative of monochloramine, was selected to induce oxidative injury because of the well-known oxidative capacity of active chlorine compounds. The ability of chloramine-T to induce tissue damage results from its cleavage of peptide bonds and by chlorination of aromatic rings, oxidation of thiols and thioesters, decarboxylation reactions, and chlorination of N-terminals [6,7]. The increased oxidative capacity and decreased bactericidal capacity of chloramine-T compared with monochloramine made it an ideal agent for this study [8].
After injection of the intestinal lumen with either saline or chloramine-T, a third 4-0 silk suture was placed 1 cm distal to the most proximal suture, and the injection site was resected to avoid injury-induced necrosis from the puncture site. The mesenteric blood supply to the intestine remained intact. The intestines were then returned to the abdominal cavity in discontinuity, and the laparotomy was closed in two layers. After 24 h, all animals were euthanized, and the intestines were exteriorized and inspected visually for gross necrosis. The isolated loop was harvested aseptically and divided for histologic and microbial analysis.
To determine if the denser population of cecal bacteria has the capacity to home to the site of proximal bowel injury, we injected the pathogenic bacterium Pseudomonas aeruginosa (PA) into the cecum. Three groups of mice were studied. Group 1 consisted of control mice, which underwent a laparotomy as described previously followed by intracecal injection of 100 mcL of a solution of bioluminescent PA at a concentration of 107 colony-forming units (CFU)/mL. The intestine was returned to the abdominal cavity, and the laparotomy was closed. Group 2 consisted of mice that underwent laparotomy and creation of an isolated ileal loop, after which 200 mcL of sterile saline was injected into the lumen of the isolated loop. After loop construction, all mice in Group 2 received an intracecal injection of 100 mcL of bioluminescent PA 107 CFU/mL via a laparotomy followed by abdominal closure. Group 3 consisted of mice that underwent laparotomy and creation of an isolated ileal loop as described previously. Then 200 mcL of 0.05% chloramine-T was injected into the lumen of the isolated loop. After loop construction, all mice in Group 3 received an intracecal injection of 100 mcL of bioluminescent PA 107 CFU/mL, following which, the abdomen was closed. After 24 h, all animals were euthanized, and the gastrointestinal tract, from the stomach to the rectum, the liver, and the spleen, were harvested from each animal and placed in Petri dishes for photon camera (Xenogen Corp., Alameda, CA) imaging of luminescent bacteria as described below.
On the basis of the results from the above experiments, we sought to determine the rate at which bacteria migrated from the cecum to the more proximal bowel, liver, and spleen. Animals underwent laparotomy and construction of an isolated ileal loop as described. Then 200 mcL of either 0.05% chloramine-T or sterile saline was injected, followed by intracecal injection of 100 mcL of bioluminescent PA at a concentration of 107 CFU/mL. Mice were sacrificed at 2, 6, 8, or 24 h after the operation; and the intestine, liver, and spleen of each animal were harvested and prepared as described for photon imaging. After initial imaging, all samples were maintained undisturbed at room temperature overnight to allow further bacterial proliferation to enhance detection by photon imaging. Each sample was re-imaged with the Xenogen camera 24 h after the start of the experiment.
Bacterial preparation
A well-characterized, constitutively luminescent strain of PA, XEN41 (Calpier Life Sciences, Inc., Hopkinton, MA) was used for all experiments. After overnight growth on a tryptic soy broth (TSB) agar plate, colonies were suspended in sterile saline and diluted to achieve a final optical density of 0.02 at 600 nm, corresponding to 1×107 CFU/mL. This suspension was placed on ice and utilized within 2 h of preparation.
Histologic analysis
Harvested tissue was placed immediately in 10% phosphate-buffered formalin for at least 12 h. Tissue was then embedded in paraffin, stained with hematoxylin and eosin, and analyzed for histologic damage.
Bacterial community analysis
The entire section of the isolated loop was homogenized in sterile saline. Five serial dilutions of the homogenate were performed in sterile saline, and the remaining solution was plated on TSB agar plates and incubated overnight at 37°C. All culture plates were taken to the University of Chicago Clinical Microbiology Laboratory for identification of bacterial species by standard microbiologic techniques. Bacterial communities present in control animals and those with saline-treated loops and chloramine-T-treated loops with and without necrosis were compared. Statistical analysis was performed with SPSS (Chicago, IL), and p<0.05 was considered significant.
Xenogen imaging
At 2, 6, 8, and 24 h after intracecal injection of luminescent PA, mice were anesthetized with sevofluorane and sacrificed. The gastrointestinal tract from the stomach to the rectum, including the spleen and liver, were harvested immediately and placed in Petri dishes. The organs were imaged ex vivo within 30 min of harvest using the photon camera with a 30-second exposure time [9]. After initial imaging, organs from the 2, 6, and 8 h time points were maintained undisturbed at room temperature overnight and re-imaged with the photon camera approximately 24 h after the original experiment start time.
Results
Injection of 0.05% chloramine-T into isolated ileal loops results in gross necrosis in 59% of animals
The majority of mice survived to the time of sacrifice (saline-treated loop 78%; chloramine-T-treated loop 63%). No intestinal necrosis was detected in animals that died prior to the 24 h study endpoint. Of the animals that survived to 24 h, no gross necrosis was observed in the saline-treated group; however, 59% of the animals in the chloramine-T-treated group had gross transmural necrosis (p<0.001) (Fig. 1). Greater histologic damage was observed in necrotic loops than in control animals, animals with saline-injected loops, and animals with chloramine-T-injected loops that did not develop gross necrosis (Fig. 2).

Gross necrosis after oxidative injury. After intraluminal injection of 0.05% chloramine-T into isolated ileal loops, gross necrosis was detected in 59% of animals, whereas no necrosis was apparent in animals that underwent sterile saline injection into the isolated loop (p<0.001).

Histologic analysis of isolated loop architecture. Damage was observed in the sections prepared from loops with necrosis compared with control animals, animals with saline-injected loops, and animals with chloramine-T-injected loops that did not develop gross necrosis.
Lactobacillus is not detectable after surgical instrumentation, whereas overall gut bacterial community structure remains similar in mice with and without necrosis
After bacterial species identification via standard microbiologic techniques, results were sorted into one of four groups: untreated controls (n=5), saline-treated loops (n=7), chloramine-T-treated loops without necrosis (n=17), and chloramine-T-treated loops with necrosis (n=10). Relative bacterial community structure was analyzed for each group. Lactobacillus was detected in approximately 60% of control animals, whereas it was not detected in any animal that underwent a surgical procedure (Fig. 3). Methicillin-sensitive Staphylococcus aureus, PA, and Enterococcus faecium were detected only after surgical intervention, but their relative abundance did not differ by loop type (i.e., chloramine-T or saline) or by the presence or absence of necrosis (Fig. 3).

Analysis of loop bacterial community structure. After bacterial species identification via standard microbiologic techniques, results were sorted into four groups: Untreated controls (n=5), saline-injected loops (n=7), chloramine T-injected loops without necrosis (n=17), and chloramine-T-injected loops with necrosis (n=10). Bacterial community structure was analyzed for each group. Lactobacillus was detected in approximately 60% of control animals; however, it was completely depleted in all animals that underwent a surgical procedure. Methicillin-sensitive Staphylococcus aureus, PA, and Enterococcus faecium were detected only after surgical intervention. Their relative abundance did not differ by loop treatment (i.e., chloramine-T or saline) or by the presence or absence of necrosis.
Surgical instrumentation promotes cecal bacterial migration to the proximal intestinal tract
Photon imaging demonstrated that 24 h after intracecal injection of bioluminescent PA in animals that did not undergo construction of an isolated intestinal loop, all luminescent bacteria had been cleared from the gastrointestinal tract (Fig. 4). Further, there were no luminescent bacteria in the liver or spleen of control animals. However, 24 h after intracecal injection of luminescent PA in mice that underwent construction of a saline- or chloramine-T-treated loop, retention of bacteria in the cecum with proximal extension into the isolated loop and the entire proximal gastrointestinal tract was detected (Fig. 4). In addition, luminescent bacteria were apparent in the liver and spleen of each of these animals.
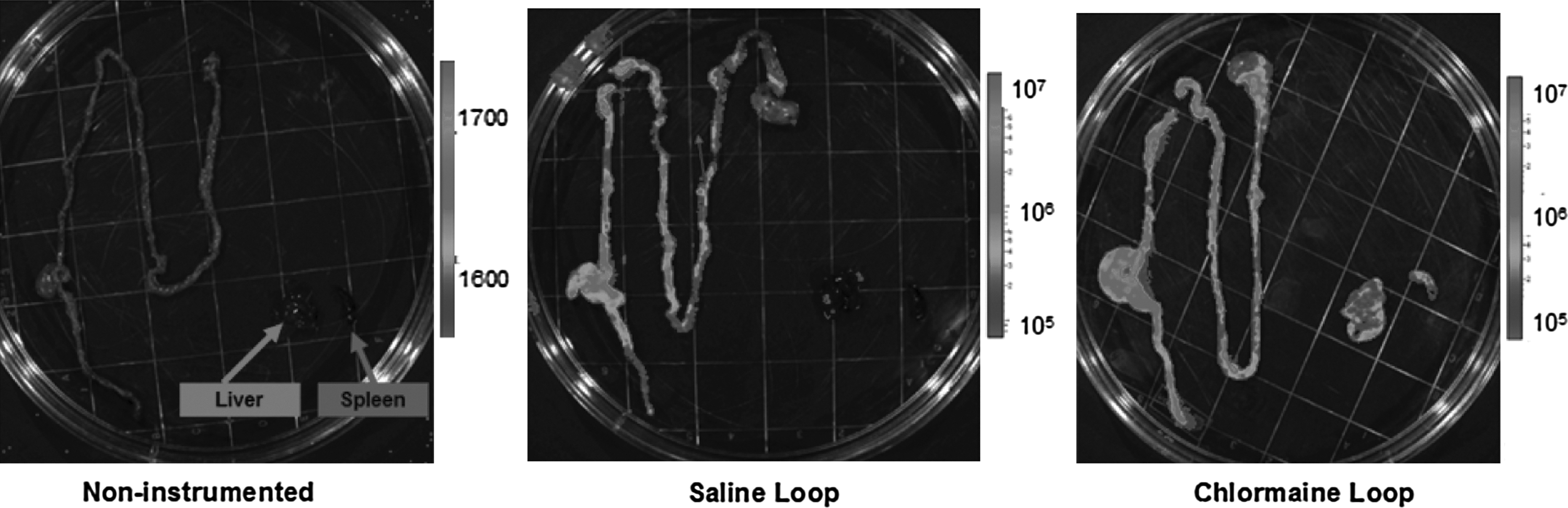
Xenogen camera imaging 24 h after intracecal injection of bioluminescent Pseudomonas aeruginosa (PA) in animals that did not undergo construction of an isolated intestinal loop, demonstrating that all luminescent bacteria had been cleared from the gastrointestinal tract. There also were no luminescent bacteria in the liver or spleen of control animals. However, 24 h after intracecal injection of luminescent PA in mice that underwent construction of a saline- or chloramine-T-injected loop, bacteria were retained in the cecum, with proximal extension into the isolated loop and the entire proximal gastrointestinal tract. Further, luminescent bacteria were detected in the liver and spleen of all of these animals.
Delayed photon imaging of luminescent bacteria enhances detection
Xenogen camera imaging of luminescent bacteria 2, 6, and 8 h after intracecal injection demonstrated low to no signal intensity (Fig. 5). However, after overnight incubation of the tissues at room temperature, the signal was enhanced (Fig. 5), suggesting that bacterial proliferation occurred during the interval between the two imaging events. Delayed imaging results did not differ between animals with a chloramine-T-treated loop and those with a saline-treated loop.

Xenogen camera imaging of luminescent bacteria 2, 6, and 8 h after intracecal injection, demonstrating little or no signal. However, on repeat imaging 24 h later, intense signal was observed.
Discussion
Full-thickness intestinal necrosis leading to perforation and sepsis likely involves complex pathophysiologic mechanisms that include a systemic insult (hypotension, hypoxia), release of inflammatory and tissue-destroying mediators (i.e., cytokines, oxidants), and pathogenic bacteria (i.e., PA). Intestinal necrosis with perforation is rare, yet is seen in a variety of clinical scenarios, including after cardiac bypass with shock and in prematurity. Despite decades of investigation, our understanding of the exact role of microorganisms in the pathogenesis of intestinal necrosis is poor. Difficulty in designing an experimental model that recapitulates accurately the gross necrosis observed clinically contributes to the limitations in our understanding of the disease. Surgeons operate where there is necrosis, and necrosis defines the surgical disease, but its pathogenesis is unclear. Models are needed that produce full-thickness necrosis consistently.
One of the primary advantages of our model is that, unlike most models in which the entire bowel is utilized to study a patchy disease process, it offers consistent generation of isolated sections of necrosis, allowing in-depth investigation. Although the model does not necessarily represent NEC entirely accurately, targeted analysis of intestinal necrosis does permit site-specific interrogation, which could enhance our understanding of the complex interaction between microorganisms and the host inflammatory response. Uncovering the complex bidirectional crosstalk between intestinal bacteria and host tissues is now possible with high-resolution molecular analysis applied to appropriate animal models.
Our initial analysis of bacterial community structure was notable for complete depletion of Lactobacillus after surgical instrumentation. A similar decrease in Lactobacillus has been demonstrated recently in critically ill patients [10]. Given its potentially protective properties [11], post-operative depletion of Lactobacillus may cause loss of colonization resistance to other, more harmful, pathogens and therefore likely is important. Also of interest was the finding that gut bacterial community structure did not differ in the presence or absence of necrosis. This finding suggests that it is not merely the presence of a certain microorganism or community of bacteria that promotes necrosis but rather qualitative differences in the properties of a given pathogen or community of pathogens that cause disease. Such characteristics can now be understood better using metagenomic/metatranscriptomic analysis at specific sites of disease such as necrotic intestine [12]. Properties such as acquisition of new pathogen islands, enhanced virulence capacity, and antibiotic resistance could enhance the virulence of the microbiota [13–15].
This model also has allowed the use of bioluminescent bacteria after loop construction to determine patterns of migration. Surprisingly, after intracecal injection of luminescent PA, bacteria were found to migrate within 24 h to the isolated loop, the proximal small bowel, the liver, and the spleen. This migration occurred despite bowel discontinuity, suggesting hematogenous or lymphatic rather than direct spread. Further work is under way to assess bacterial dissemination/migration patterns in this model more comprehensively.
Although our efforts to determine the rate at which microbes migrated throughout the gastrointestinal tract offered little insight into this issue, these studies did identify an important methodological concern. Specifically, most studies that use Xenogen camera imaging of bioluminescent bacteria image tissue immediately after harvest. Interestingly, when we imaged organs immediately from animals sacrificed 2, 6, and 8 h after injection of luminescent PA, little signal was detected. However, after continued incubation at room temperature, repeat imaging 24 h after the start of the experiment demonstrated a relatively robust signal in most of the animals. This suggests that immediately after harvest, luminescent bacteria may be present in various locations at a density too low to generate a detectable signal. Delayed imaging thus may provide more accurate detection of luminescent bacteria that are alive and proliferating
Although use of this novel model of intestinal necrosis may enable more targeted analysis of the role of bacteria and oxidizing agents in the development of necrosis, this study is not without limitations. First, we utilized three-week-old rather than newborn or pre-term mice, which may be more representative of the immature gut. This limits the direct applicability of our results to NEC pathogenesis. We elected to utilize three-week-old mice because they are weaned and able to live independently. Further, the older animals are better able to tolerate surgical intervention than are younger ones. However, to analyze the impact of this model on the immature gut realistically, future studies will require newborn mice. Our work also is limited by reliance on culture-based investigation. Recent work has demonstrated that only approximately 20% of bacteria associated with the human body are detectable by standard laboratory culture [16]. Application of culture-independent techniques (i.e., 16S ribosomal RNA pyrosequencing) to this model will permit more robust analysis of the bacteria that contribute to necrosis.
Our model also may have been impacted by systemic factors that were not accounted for. For example, 22% of mice that underwent construction of a saline-treated loop and 37% of mice that underwent construction of a chloramine-T-treated loop died without developing intestinal necrosis. Previous work from our laboratory has demonstrated that surgical stress alone can trigger alterations in bacterial virulence that lead to gut-derived sepsis and a high mortality rate [17]. Further, the creation of a complete mechanical bowel obstruction in these animals may have caused systemic stress that increased the mortality rate. Future studies will include characterization of microbial virulence gene expression and systemic inflammatory changes after surgery to gain a better understanding of why animals in the model experience a 22%–37% mortality rate that is not associated with the development of intestinal necrosis. Direct analysis, not only of systemic inflammatory changes but also of changes in mucosal cytokines in necrotic and non-necrotic tissue would offer a unique perspective regarding the inflammatory changes associated with necrosis. Future studies will include this analysis.
In summary, we demonstrate that intraluminal chloramine-T causes full-thickness, gross intestinal necrosis in three-week-old mice. Interrogation of this novel model demonstrated that surgical injury resulted in the depletion of protective Lactobacillus from the gut. Further, although particular bacterial species were not determinants of the development of necrosis, reliance on this model will permit in-depth metagenomic interrogation of host–pathogen interaction within a confined space that consistently undergoes transmural necrosis similar to that observed in human NEC. It is possible that virulence factor production by bacteria, rather than bacterial composition, plays a more important role in the pathogenesis of intestinal necrosis. Further work will be necessary to clarify this potential mechanism.
Footnotes
Author Disclosure Statement
The authors have no disclosures.