
Abstract
Abstract
Background:
Staphylococcus aureus continues to be an important pathogen for surgeons. Surgical infections of interest caused by this pathogen include community-acquired infections, especially of skin and soft tissue; surgical site infections; and nosocomial infections during the post-operative period.
Methods:
A literature review was conducted to detail the history of S. aureus in surgical care; to identify the structural elements of the pathogen, the secreted exotoxins and enzymes, and the changing resistance patterns that make S. aureus an ever-formidable clinical foe; to consider whether screening for the pathogen and directed antimicrobial prophylaxis are useful; and to identify antimicrobial agents employed currently in treatment of established infections.
Results:
Staphylococcus aureus has shown a pattern of increasing prevalence of infections among hospitalized patients and a pattern of increasing resistance. A broad array of virulence factors account for its continued pathogenicity, and new virulence characteristics continue to be acquired. Staphylococcus aureus has an uncanny ability to adapt and develop resistance to each new antibiotic as it is introduced into clinical care. Although screening and directed surgical prophylaxis against methicillin- (now sometimes referred to as meticillin-) resistant S. aureus (MRSA) is controversial, effective efforts to decrease the incidence of nosocomial infections may be decreasing the numbers of MRSA infections encountered, and current agents remain effective for therapy.
Conclusions:
Staphylococcus aureus continues to be a major pathogen for surgical patients. The scope of infections has expanded recently to include toxic shock syndrome, severe community-acquired soft tissue infections with unique virulence characteristics, and highly resistant pathogens. The adaptability of the organism requires that new prevention and treatment strategies be developed continuously to meet the challenge of future infections.
The high prevalence of S. aureus infections in surgical patients has led to vigorous efforts to develop specific preventive and therapeutic measures directed against this pathogen. The threat of these infections has led to intensive efforts in the prevention of transmission by enhanced infection control practices, the liberal use of topical antiseptics, and specific systemic antibiotics. Despite these efforts, prevention has remained elusive, and therapeutic management of established infections continues to be difficult. An important key that has led to suboptimal prevention and treatment of S. aureus infections has been the rapid evolution of resistance to antimicrobial agents, and the development of new virulence factors.
In this paper, the changing epidemiology, risk profile, and antimicrobial susceptibility of S. aureus are reviewed, as are prophylaxis and therapy. This ever-changing phenotype of resistance and virulence is leading to new challenges and changes in the prevention and treatment of infection in the surgical patient.
Historical Evolution of Staphylococcus aureus
Studies of the microbiology of S. aureus led directly to the discovery of penicillin and the inauguration of the era of antibiotic treatment. An uncovered culture plate in a laboratory, an airborne fungal contaminant of the plate, and a recognized zone of inhibition about the perimeter of the unwelcome fungus resulted in the discovery of penicillin by Alexander Fleming [1,2]. Although available on a limited basis to the military during the war, penicillin and sulfonamide preparations were introduced into general clinical practice for the treatment of infection after World War II. Many believed at the time that the millennium of infection-free surgery was at hand, in that antibiotics for surgical patients held promise for both prevention and treatment of post-operative infections. This optimism was short-lived, as the first penicillin-resistant S. aureus was discovered in 1944 [3]. As the use of the first generation of antibiotics was expanded, so was the emergence of resistant strains of S. aureus, such that a pandemic was identified among hospitalized patients in the 1950s.
In 1959, methicillin (now sometimes referred to as meticillin) was introduced as a semi-synthetic penicillin with activity against penicillin-resistant S. aureus (PRSA), and it was deployed rapidly for hospital-acquired infections [4]. By 1961, the first methicillin-resistant S. aureus (MRSA) pathogens were identified. During the late 1960s and 1970s, additional semi-synthetic penicillins (e.g., oxacillin) and the first-generation cephalosporins were introduced into clinical practice to combat PRSA. By 1970, PRSA predominated in the hospital, but penicillin-sensitive pathogens persisted among community-acquired staphylococcal infections, and penicillin G still was recommended for community-acquired infections [5]. The liberal use in clinical practice of oral β-lactam antibiotics led to the extinction of penicillin-sensitive S. aureus by the mid-1970s [6]. The former PRSA pathogens then were re-named methicillin-sensitive S. aureus (MSSA), as expanded ambulatory and in-patient antibiotic use attended an increasing frequency of MRSA isolates in hospital-acquired infections. During the 1980s, this growth in the incidence of MRSA infections led to the re-emergence and widespread usage of vancomycin as the treatment of choice. Owing to cumbersome administration and the risk of ototoxicity and, to a lesser extent, nephrotoxicity, there arose considerable interest in the identification of alternative antibiotic therapies for MRSA.
As was observed with PRSA in the 1970s, in the 1990s, the paradigm was that community-acquired S. aureus infections were methicillin-sensitive whereas hospital-acquired infections became nearly uniformly MRSA. In 1993, the first community-associated MRSA (CA-MRSA) was identified [7], and in 1998, the first isolates were identified in the U.S. [8]. This MRSA variant expressed a unique genotype and phenotype, demonstrating new and unique virulence factors (e.g., Panton-Valentine leukocidin) not ordinarily seen in hospital-acquired MRSA (HA-MRSA), as well as a different pattern of antibiotic susceptibility, indicating that the organism arose de novo in the community and was not a feral, escaped hospital clone. Community-associated MRSA demonstrates susceptibility to selected non-β-lactam antibiotics (e.g., co-trimoxazole) and is now recognized worldwide as a virulent pathogen [9]. The rate of infections with both HA-MRSA and CA-MRSA increased rapidly in the U.S., the former until 2008, at least.
A recent report identified a reduction in 2008 rates of HA-MRSA infection [10] (Fig. 1). Whether this reduction signals a true decline or plateau in the frequency of HA-MRSA infections remains to be determined. Such a reduction is plausible owing to recent reductions in the rates of certain nosocomial infections often caused by S. aureus, including central line-associated blood stream infection (CLABSI) and VAP [11–13], and possibly to recent controversial efforts to screen for and eradicate the staphylococcal carrier state as a tactic to reduce infections after surgery and in the intensive care unit (ICU) [14–17]. How these efforts will mitigate MSSA infections, both in the community and in healthcare-associated settings, is an open question.
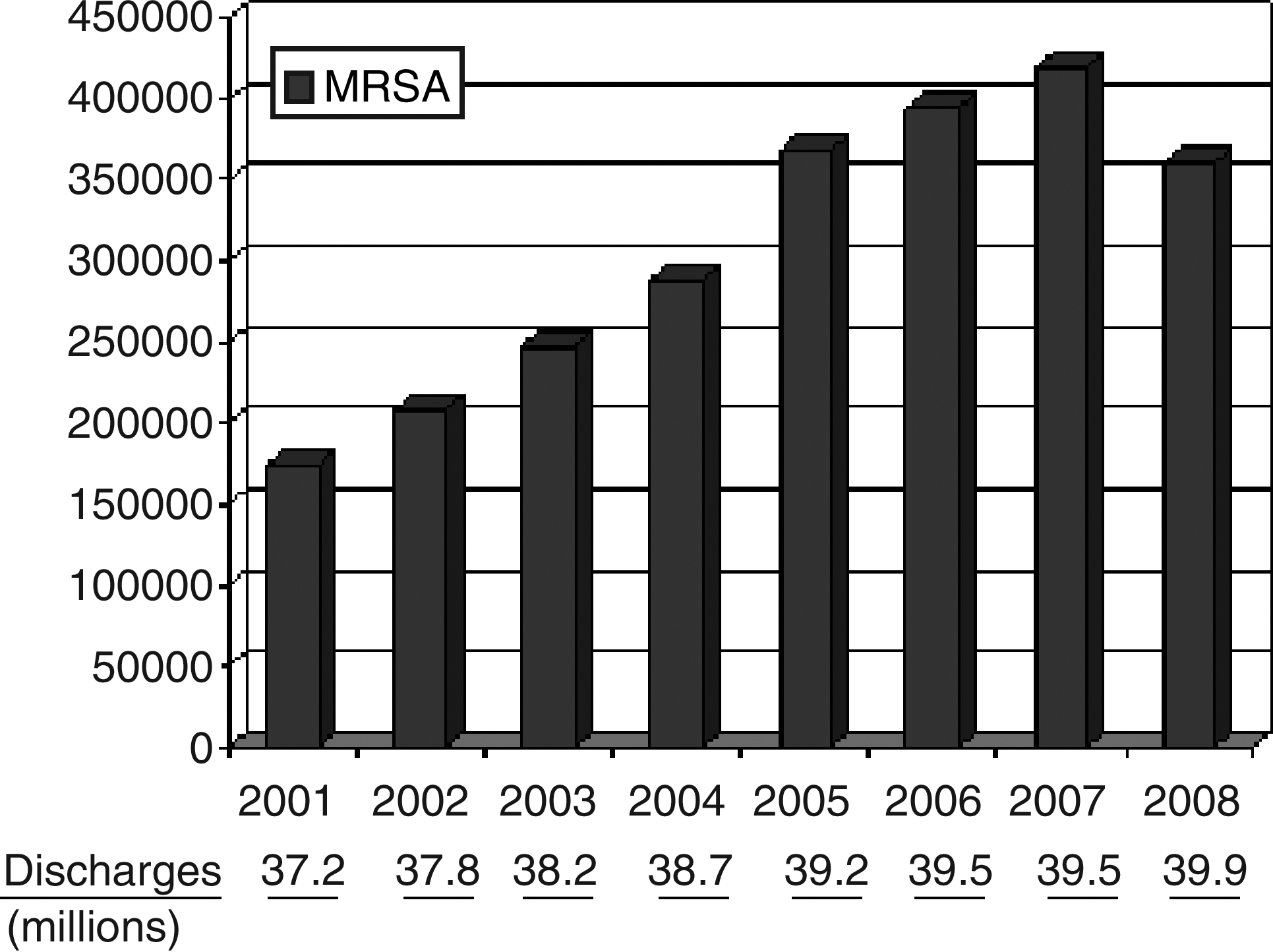
Increase in Infections with Methicillin-Resistant Staphylococcus aureus in Hospitalized Patients in the U.S. from 2001 to 2008 (Agency for Healthcare Research and Quality. HCUPnet. http://hcupnet.ahrq.gov/ Accessed December 10, 2010.)
The historical evolution of resistance of S. aureus continued with evidence of increasing minimum inhibitory concentrations (MICs) among vancomycin-susceptible isolates, vancomycin-intermediate resistance (VISA), and even a small number of vancomycin-resistant isolates (VRSA) (see below). It is likely that the continued use of antibiotics against MRSA will create environmental pressure for the continued decrease in the prevalence of infections caused by MSSA and evolution of the next generation of S. aureus that is highly resistant to current and future antibiotic agents [18].
Mechanisms of Virulence and Resistance
Understanding the complex issues of virulence and resistance in S. aureus requires at least a limited knowledge of bacterial genetics [19,20]. The genome of S. aureus is a single circular chromosome. This haploid genetic configuration means that changes in genotype have a direct influence on phenotypic expression (e.g., antibiotic resistance). The genome consists of 75 percent core genes that appear in all species and 25 percent accessory genes that are highly variable among strains. Within the accessory component of the genome are mobile genetic elements that are transferable among S. aureus strains. Accessory components may consist of pathogenicity islands, prophages (from bacteriophages), staphylococcal cassette chromosomes (SCC), genetic islands, and plasmids. Whereas specific definitions are used to distinguish mobile genetic elements one from another, a common feature is that specific clusters of transferrable genes move together, leading to expression of common phenotypes. Virulence factors of S. aureus may be common to all species (e.g., coagulase), or they may be highly specific for each strain. This ability to exchange mobile genetic units among strains of S. aureus, but also to acquire genetic material from other bacterial species (e.g., Enterococcus), results in patterns of virulence and resistance that are changing continually.
Microbial virulence factors of S. aureus generally have been grouped into bacterial structural factors, secreted bacterial products and enzymes, and resistance mechanisms to antimicrobial agents. Structural elements of the bacterial cell include a polysaccharide capsule that resists phagocytosis, and the peptidoglycan cell wall that anchors a number of surface adhesion proteins, permitting microbial binding to the extracellular matrix of targeted host tissues [21]. The surface adhesion proteins of the cell wall are referred to collectively as microbial surface components recognizing adhesive matrix molecules (MSCRAMMs) [19–21]. The MSCRAMMs are identified in many species of gram-positive organisms, of which several are unique to S. aureus. Table 1 identifies many (but not all) of the MSCRAMMs that have been identified [22–29].
A virulence factor that is a secreted product, but also part of the staphylococcal bacterial cell structure, is a polysaccharide and protein matrix, or biofilm [30–32]. Biofilm is produced by several bacterial species, including selected strains of S. aureus, after adherence of the bacterial cell to a foreign surface. Biofilm encases a community of cells and changes the phenotype from planktonic (rapidly dividing) to dormancy, affording barrier protection from host defenses and imperviousness to antibiotics [33]. Not all strains of S. aureus are biofilm producers, but those that are have been identified in particular as pathogens associated with device-related infections.
Products secreted by S. aureus include those that enhance microbial adherence to host tissues, exotoxins that are directly toxic to host cells, and enzymes that degrade and digest the extracellular matrix. Secretable expanded repertoire adhesive molecules (SERAMs) are a group of secreted proteins that have duplicative functions of the MSCRAMMs identified above, except that they are secreted proteins, not part of the staphylococcal cell wall [34]. A SERAM of considerable interest for S. aureus is coagulase, which targets prothrombin and fibrinogen. The result of the action of coagulase is generation of a staphylococcal–fibrin matrix and creation of a local environment that facilitates microbial adherence to tissue and a protected environment against host defense mechanisms and antimicrobial therapy [34]. Coagulase has effects that are analogous to clumping factor of the MSCRAMMs, but it is a freely secreted and unique protein [35].
Exotoxins are those secreted products that are cytotoxic to host cells, whereas the superantigens are secreted products that provoke an over-exuberant inflammatory response by the host. Individual cytotoxins have different cell targets but generally function similarly by damaging the cell membrane, creating pores, and leading to extravasation of cellular contents. The cytotoxin of greatest recent interest has been the Panton-Valentine leukocidin (PVL) [36], which is but one of several leukocidins that are produced. The PVL consists of two proteins that insert into the plasma membrane of leukocytes and create pore-mediated damage to the target cell. The PVL has been particularly associated with CA-MRSA and has been implicated in the increased virulence of the community-acquired vs. the healthcare-acquired (HA-)MRSA [37].
A representative selection of exotoxins is identified in Table 2 [38–50]. The superantigen toxins identified in Table 2 are powerful immuno-stimulatory proteins. They provoke an exaggerated response from T cells, with massive pro-inflammatory cytokine release being the result. The virulence expression from these superantigens is not necessarily from direct cytotoxicity, but rather from the consequences of severe local inflammation. For S. aureus, these superantigens play an important role in severe enterocolitis (food poisoning), scalded skin syndrome, and toxic shock syndrome.
Staphylococcal Resistance Mechanisms and their Clinical Consequences
Since the introduction of antibiotics into clinical practice, the rapid evolution of resistance by S. aureus has been of great interest to, and problematic for, clinicians. Changes in the penicillin-binding protein on the staphylococcal cell membrane was the mechanism for the emergence of resistance to penicillin, but more recently also has been important in the emergence of methicillin resistance. Using pulsed-field gel electrophoresis (PGFE), eight strains of MRSA have been characterized and labeled as PGFE USA100 through USA800 [51]. The resistance determinant lies in the staphylococcal cassette chromosome within the accessory portion of the genome (SCCmec) [52]. However, the genetics of resistance differs among these strains. Seven SCCmec gene cassettes have been identified in MRSA. Types I, V, VI, and VII of SCCmec are lower molecular weight cassettes of genes and typically carry only resistance for β-lactam antibiotics. By contrast, SCCmec type IV has been most commonly associated with the prevalent CA-MRSA in the U.S. (USA300) [53]. Accordingly, the CA-MRSA has retained susceptibility to non-β-lactam antibiotics such as clindamycin, trimethoprim/sulfamethoxazole, and others. By contrast, HA-MRSA has the higher molecular weight SCCmec types II and III gene cassettes, which results in resistance to most non-β-lactam antibiotics as well. The continuing genetic changes within the accessory genes, the acquisition of plasmids with additional resistance genes, and the increasing presence of CA-MRSA within the hospital environment suggest that the phenotypic difference in susceptibility among all strains of MRSA is likely to become less distinct [54–56].
A looming threat that may enhance dramatically the adverse consequences of S. aureus infection is vancomycin-intermediate (susceptibility) resistant S. aureus (VISA) and vancomycin-resistant S. aureus (VRSA). First reported in 1997 [57], VISA (historically, with minimum inhibitory concentrations [MICs] 4–16 mcg/mL) appears to be increasing steadily in frequency. Moreover, although controversial, “MIC creep” has been observed among S. aureus isolates within the susceptible range [58], meaning that even among isolates that are “susceptible” (i.e., vancomycin-susceptible S. aureus; VSSA) by definition [MIC <2 mcg/mL], susceptibility is reduced. This correlates clinically with an increased risk of failure of therapy of isolates at the higher end of the range [59,60] (Table 3) and has led to revision of the susceptibility breakpoints for S. aureus to vancomycin by the Clinical and Laboratory Standards Institute (Table 4) [61] to decrease the possibility of treatment failure by labeling the formerly “borderline” VSSA isolates as being either VISA or VRSA, thereby influencing antibiotic choice. Higher reported prevalences of VISA and VRSA will be the inevitable result.
CI=confidence interval; MIC=Minimum inhibitory concentration.
From reference 60.
From reference 61.
The definition of VISA (i.e., MIC of 4–8 mcg/mL by the 2006 revised breakpoints) and the laboratory methods for testing isolates (disc diffusion can be inaccurate) also have become issues [62]. The VISA phenotype is not the consequence of vancomycin-resistance genes transmitted from enterococci. The natural history of VISA expression is believed to begin with adaptive thickening of the cell wall of the occasional heteroresistant organism, which leads to its emergence as the dominant phenotype when exposed to environmental pressures, including vancomycin use. The thickened cell wall impedes penetration of the vancomycin molecule, and up-regulates expression of peptidoglycan binding sites to create the relative resistance problem [63]. Unremitting vancomycin utilization pressure is likely to make the evolution of VISA a continuing problem.
Heteroresistant VISA (hVISA) strains are those for which MICs conventionally indicate susceptibility (≤2 mcg/mL) by routine testing, except when high-density inocula are used. Within such inocula, minority sub-populations emerge for which MICs are in the intermediate range (4–8 mcg/mL) [64]. The detection of hVISA requires inocula containing >106 bacteria, because resistant clones occur infrequently spontaneously (∼1/1,000,000) [65]. The MICs for most heteroresistant clones are in the 4–8 mcg/mL range [66], but unstable heteroresistance can be detected [66,67]. Clones of S. aureus demonstrating unstable heteroresistance grow in the presence of high concentrations of vancomycin (>4 mcg/mL), although conventional MICs for such clones are not elevated; i.e., non-susceptible clones revert rapidly to normal phenotypes and therefore cannot be detected by the usual laboratory tests. Clinical failure with vancomycin or MICs to vancomycin of ≥2 mcg/mL should alert the laboratory to proceed with the more specific methods of population analysis profiling along with hVISA screening performed by the macro-Etest (MET) to identify hVISA or VISA isolates [68]. Both VISA and hVISA appear to emerge in SCCmec II isolates among vancomycin-exposed patients and are detected better by MET [69].
A total of 11 VRSA infections have now been identified in the U.S. (Table 5) [70–79]. It appears that the vanA gene was acquired by plasmid-mediated gene transfer from vancomycin-resistant enterococci encodes for highly resistant VRSA. All VRSA are methicillin resistant. Most are classified as USA100. These clinical infections have been associated with preceding MRSA or enterococcal infections, and the patients have had prior vancomycin therapy. Chronic co-morbid conditions are present, notably chronic kidney disease/end-stage renal disease. All infections followed surgical care or have occurred in patients with soft tissue infections (e.g., chronic ulcers), where surgeons are involved in patient management.
Modified from U.S. Centers for Disease Control and Prevention: CDC reminds clinical laboratories and healthcare infection preventionists of their role in the search and containment of vancomycin-resistant Staphylococcus aureus, May 2010. www.bt.cdc.gov/coca/reminders/2010/2010may06.asp) [70].
Clinical Consequences of Staphylococcal Infection
Staphylococcus aureus can cause a host of infections, of skin and soft tissue, the blood stream, the lungs, and prosthetic devices, among others (Table 6). The 15 heat-stable (and thus not destroyed by cooking) enterotoxins elaborated by S. aureus may cause food-borne disease characterized by malaise, nausea, vomiting, abdominal pain, and diarrhea. The epidemiology of infection depends on the organism (MSSA vs. HA-MRSA vs. CA-MRSA) and the clinical setting (outpatient vs. inpatient vs. ICU). In a retrospective analysis of the Agency for Healthcare Research and Quality Nationwide Inpatient Sample database (2000 and 2001 editions), S. aureus infection was reported as a discharge diagnosis for 0.8% of hospitalized patients, and was estimated to include 292,045 inpatients/year in the U.S. [80]. Mean hospital length of stay was increased three-fold, from 4.5 days to 14.3 days, and hospital charges also were increased three-fold, from $14,141 to $48,824. The risk of in-hospital death increased nearly five-fold, from 2.3% to 11.2%. In 2005, more than 85% of the deaths of hospitalized patients with staphylococcal infections were attributed to HA-MRSA [81]. Although attribution of death is complex and error-prone for patients with multi-system disease, and estimates do not always account properly for differences in severity of illness and delays or errors in treatment (or serious morbidity therefrom), the risk of death from invasive MRSA infection (i.e., bacteremia) is approximately double that of MSSA infection [82,83]. Excess mortality and hospital cost has been associated as well for SSIs caused by MRSA [84], but other data are conflicting [85].
Prevention of MRSA Infection
Population-based studies have consistently identified male gender and both extremes of age as important risk factors for the acquisition of staphylococci. Dialysis dependence, diabetes mellitus, cancer, human immunodeficiency virus infection, intravenous drug use, alcohol abuse, recent surgery, critical illness, and staphylococcal nasal carriage have been identified as independent risk factors [86,87]. Therefore, identification of the patient at risk is a crucial first step. Much recent work in prevention of invasive staphylococcal infections focused on prevention of colonization of the patient at risk and decolonization of staphylococcal carriers. Although work is being pursued with novel antimicrobial molecules, other tactics being investigated include phage therapy, molecules aimed at blocking regulation of virulence determinants and surface adhesins, and vaccination. Although anti-staphylococcal vaccines have been raised against a variety of targets, multiple efforts have failed to demonstrate efficacy, including in Phase III trials.
Individuals who are nasal carriers (the primary locus, although other sites are possible) of S. aureus may represent as much as 50% of the population, although the persistent (rather than intermittent) carriers make up 20%–30% of healthy people. Most persistent carriers (as many as 90%) harbor a single strain of MSSA, although MRSA nasal carriage is recognized to be increasing (now up to 10% of carriers). Patients who are persistent carriers are at higher risk of acquiring S. aureus infection. Eradication of the carrier state decreases the risk of nosocomial infection, but routine screening for the carrier state is controversial in that the evidence extant is mixed regarding effectiveness [15,88]. Eradication of the carrier state with topical nasal 2% mupirocin ointment twice or thrice daily for five to seven days may be effective about 60% of the time if undertaken according to established protocol [89], but treatment failures are common [90]. Alternatively, topical application of 2% chlorhexidine gluconate-impregnated cloths to critically ill trauma patients as a “sponge bath” decreased the rates of colonization with MRSA, and MRSA VAP [91].
It has long been speculated that alteration of antimicrobial prophylaxis regimens can be used as an anti-MRSA infection control tactic. Fukatsu et al. demonstrated that using first- rather than third-generation cephalosporins for surgical prophylaxis and reducing the duration of prophylaxis to emphasize pre- and intra-operative rather than postoperative dosing was associated with control of an outbreak of post-operative MRSA SSIs [92]. However, providing an anti-MRSA agent (e.g., intravenous vancomycin) as surgical prophylaxis generally is discouraged [93–95] unless the patient has an allergy to beta-lactams, and even then, clindamycin is preferred [95]. However, countervailing arguments in favor of vancomycin prophylaxis for cardiac [95] and orthopedic surgery have been put forth. Vancomycin is the recommended antibiotic for prophylaxis of major surgical procedures when MRSA is a source of concern [95–97]; although the threshold level of prevalence of MRSA SSI is vague in the literature, it may be as low as 10% [96]. In a randomized clinical trial in patients undergoing cardiac surgery, vancomycin was compared with cefazolin for prevention of SSI in a MRSA SSI high-prevalence medical center with high MRSA infection rates [98]. Overall, SSI rates in the two groups were the same, but cefazolin patients had a higher rate of MRSA infection, whereas vancomycin patients had a higher MSSA infection rate. Moreover, vancomycin prophylaxis of SSI has not been subject to rigorous evaluation in clean elective surgery, despite the recommendation to use it when MRSA is the target pathogen. In the case of orthopedic surgery, a bewildering array of delivery vehicles have been described (e.g., topical vancomycin powder [99], antibiotic-loaded cement or allograft bone [100]) without much scientific rigor, leaving this question largely unanswered.
Ultimately, the tenets of infection control—hand hygiene, barrier precautions, and responsible antibiotic prophylaxis—-are the most effective prevention. Concerted effort has reduced the incidence of both CLABSI and VAP recently [101,102]. In 2001, an estimated 43,000 CLABSIs occurred among patients hospitalized in ICUs in the U.S. [103]. In 2009, the estimated number of ICU CLABSIs had decreased to 18,000. Reductions in CLABSIs caused by S. aureus were more marked than reductions in infections caused by gram-negative bacilli, Candida spp., and Enterococcus spp. In 2009, an estimated 23,000 CLABSIs occurred among patients in in-patient wards and, in 2008, an estimated 37,000 CLABSIs occurred among patients receiving out-patient hemodialysis. In 2009 alone, an estimated 25,000 fewer CLABSIs occurred in U.S. ICUs than in 2001, a 58% reduction. As many as 6,000 lives and $414 million in excess health-care costs were saved in 2009, and approximately $1.8 billion in cumulative excess health-care costs have been saved since 2001.
These efforts may be decreasing the number of invasive MRSA infections overall. According to data submitted to the National Healthcare Safety Network (HNSN) of the U.S. Centers for Disease Control and Prevention (CDC), the incidence of MRSA CLABSI decreased significantly in all U.S. ICU types from 2001 through 2007 except in pediatric units, for which incidence rates remained static [13,104]. Decreases in MRSA CLABSI incidence ranged from −51.5% (95% confidence interval [CI] −33.7%, −64.6%; p<0.001) in non-teaching-affiliated medical-surgical ICUs (0.31 vs. 0.15/1,000 central line days) to −69.2% (95% CI −57.9%, −77.7%; p<0.001) in surgical ICUs (0.58 vs. 0.18/1,000 central line days). In all ICU types, the MSSA CLABSI incidence decreased from 1997 through 2007, with changes in incidence ranging from −60.1% (95% CI −41.2%, −73.1%; p<0.001) in surgical ICUs (0.24 vs. 0.10/1,000 central line days) to −77.7% (95% CI −68.2%, −84.4%; p<0.001) in medical ICUs (0.40 vs. 0.09/1,000 central line days). Although the overall proportion of S. aureus CLABSIs attributable to MRSA increased 25.8% (p=0.02) in the 1997–2007 period, the overall MRSA CLABSI incidence decreased 49.6% (p<0.001) over the period.
Corroborative data were obtained from a population-based study of nine diverse U.S. metropolitan areas [10]. Rates of invasive health care-associated MRSA infections over the 2005–2008 period decreased among patients for infections that began in the community and those with hospital-onset invasive disease. From 2005 through 2008, there were 21,503 episodes of invasive MRSA infection; 17,508 were health care-associated. Of these, 15,458 were MRSA BSIs. The incidence rate of hospital-onset invasive MRSA infections was 1.02 per 10,000 population in 2005 and decreased 9.4% per year (95% CI 14.7%, 3.8%; p=0.005), whereas the incidence of health care-associated community-onset infections was 2.20 per 10,000 population in 2005 and decreased 5.7% per year (95% CI 9.7%, 1.6%; p=0.01). The decrease was most prominent for the subset of infections with BSIs (hospital-onset: −11.2%; 95% CI −15.9%, −6.3%; community-onset: −6.6%; 95% CI −9.5%, −3.7%).
Treatment of MRSA Infection
The challenge of treating S. aureus infections lies particularly with MRSA infection, although MSSA infections remain common among community-onset SSTIs, and MSSA is still the most common staphylococcal pathogen of SSI. Treatment of MSSA SSTI is local drainage and debridement with appropriate β-lactam antibiotic therapy [105,106]. Pneumonia and bacteremic infections attributable to MSSA similarly require appropriate systemic β-lactam treatment; little controversy surrounds these antibiotic choices.
For surgeons, treatment of a staphylococcal infection begins with the determination of a need for debridement (in cases of SSTI and SSI), and then of the specific antibiotic to be used. By contrast, staphylococcal pneumonia and CLABSI are treated primarily with an antimicrobial agent. Staphylococcal soft tissue abscesses, even when caused by CA-MRSA, may respond to incision and drainage alone [107]; if the characteristic central, superficial area of necrosis has not suppurated, they may be managed with antibiotics alone. Surgical site infections with either CA-MRSA or HA-MRSA require drainage of the site and systemic antibiotics for erythema that extends more than 2 cm beyond the margin of the incision or for sepsis. Infections of prosthetic devices and indwelling catheters usually require removal of the infected foreign body, although mesh implanted for hernioplasty sometimes may be salvaged with antibiotic therapy and local wound care [108]. Except for soft tissue abscesses, MRSA infections require antibiotic therapy. When culture and susceptibility testing have documented MRSA infection, the selection of antibiotics is governed by those data and the severity of the infection.
Vancomycin
This glycopeptide antibiotic has been available clinically for more than 50 years, with resurgent use coincident with the emergence of MRSA infection as a major clinical problem in the 1980s. Vancomycin has a unique spectrum of coverage for gram-positive cocci and bacilli, including both CA-MRSA and HA-MRSA, but lacks any activity against gram-negative bacilli or enteric anaerobes (e.g., Bacteroides spp.). The antibacterial effect is mediated by inhibition of cell wall synthesis by mechanisms that are independent of penicillin-binding proteins. It is the MRSA antibiotic with the longest track record of use, and it is used commonly for infections of all anatomic sites. It also is prescribed commonly as an empiric choice when MRSA is suspected in patients with severe clinical infection before microbiology data become available.
Vancomycin has historically been dosed at 1 g intravenously (IV) q 12 h for patients with normal renal function, but higher doses are now recommended owing to higher MICs for MRSA for vancomycin [109,110] (Table 7) and concern about vancomycin failures [61] (see Table 3). Parenteral vancomycin is a generic preparation and remains relatively inexpensive compared with the alternatives, but the cost of mandatory monitoring of serum trough concentrations must also be accounted for.
Vancomycin has several shortcomings in clinical use. Infusion time requires at least 60 min (for a 1-g dose; 90–120 min for larger doses) to avoid complications from histamine release (e.g., “red man” syndrome, hypotension). The incidence of renal dysfunction is increasing with the use of higher doses despite the use of pharmacokinetic dosing. A major shortcoming is the apparent suboptimal effectiveness it has against MSSA. Higher mortality rates have been observed with vancomycin in bacteremic MSSA pneumonia compared with conventional β-lactam antibiotics [111] Poor lung penetration has been a suspected reason for poorer outcomes in the MSSA pneumonia patient [112]. Studies of bacteremic patients in other settings have suggested similarly that MSSA infections are best treated with β-lactam agents (preferably, nafcillin or oxacillin) rather than vancomycin [113,114].
Linezolid
Linezolid is a member of the oxazolidinone class of antibiotics, which bind to bacterial ribosomes and inhibit protein synthesis. It is bacteriostatic against all bacteria except Streptococcus pneumoniae. As does vancomycin, linezolid has antimicrobial activity only against gram-positive organisms. Unlike vancomycin, linezolid is efficiently absorbed after oral administration (tablets and an oral suspension are available), permitting effective out-patient treatment. It is generally dosed at 600 mg twice daily regardless of the route of administration or renal function. Whereas data have been presented in a subset analysis of a randomized trial suggesting superior outcomes with linezolid compared with vancomycin in complicated SSTI with MRSA [115], a prospective trial was non-confirmatory [116]. Better outcomes were identified with linezolid in a subgroup analysis of MRSA pneumonia patients in a randomized trial [117], but that conclusion has been contested [118]. Linezolid is non-inferior to vancomycin in the treatment of complicated SSTIs and in HAP/VAP. A positive feature of linezolid is activity against VISA, VRSA, and vancomycin-resistant enterococci (VRE). Linezolid does not have an indication for surgical prophylaxis and is not recommended.
The major shortcoming of linezolid has been its high cost. Both the oral and the intravenous preparation have a wholesale cost of approximately $200 per day [119]. This procurement cost exceeds by ten-fold the daily cost of intravenous vancomycin [78]. High cost has undoubtedly inhibited the use of this drug. Linezolid also is associated with myelosuppression, principally thrombocytopenia, when treatment is extended for more than two weeks [120]. Even in limited usage, S. aureus has demonstrated resistance to linezolid [121].
Daptomycin
Daptomycin is a cyclic lipopeptide antibiotic that exerts its antimicrobial action by insertion of the drug into, and depolarization of, the bacterial plasma membrane [122]. It is bactericidal rapidly. It is available only for IV use, but its long biological elimination half-life permits once-daily dosing (4 mg/kg for SSTI, and 6 mg/kg for bacteremia, although much higher doses have been reported for highly resistant pathogens or especially deep-seated infections) [123]. Daptomycin has excellent in vitro activity against MRSA and has been effective in the treatment of complicated SSTIs and in the management of MRSA bacteremia. However, daptomycin penetrates into lung tissue poorly and is inactivated by lung surfactant, and has failed in clinical trials for the treatment of gram-positive community-acquired pneumonia. It should not be used to treat HAP/VAP or empirically when HAP/VAP is in the differential diagnosis. Daptomycin does have activity against VISA, VRSA, and VRE. There is no role for daptomycin in surgical prophylaxis, for which it does not have an indication.
Daptomycin therapy has several issues. As is linezolid, daptomycin is expensive and costs more than $200/day to procure [119]. Daptomycin is associated with increases in the serum creatine phosphokinase concentration and skeletal muscle myopathy [124]. Recently, there has been a rare association with an eosinophilic pneumonia syndrome [125]. Finally, as with all antimicrobials, resistance of S. aureus to daptomycin has emerged [126].
Tigecycline
Tigecycline is a derivative of minocycline that binds to microbial ribosomes and inhibits protein synthesis [127]. It is a bacteriostatic agent with a large volume of distribution, indicating that tissue penetration is excellent but that serum concentrations decrease rapidly after administration. Tigecycline is only an IV preparation; the loading dose is 100 mg followed by 50 mg q 12 h, without regard for renal function. Tigecycline's broad spectrum of activity includes enteric gram-negative bacilli, enteric anaerobic bacteria, and gram-positive cocci, including MRSA and VRE, which may be unsuitable when targeted monotherapy is indicated. Approved indications for tigecycline include complicated SSTIs [128] and intra-abdominal infections [129].
Concern has been raised recently by a post-hoc analysis that the mortality rate of tigecycline-treated patients is higher in pooled Phase III–IV clinical trials, including unpublished registration trials [130]. The adjusted risk difference for all-cause mortality based on a random effects model stratified by trial weight was 0.6% (95% CI 0.1–1.2) between tigecycline and comparator agents. However, an independent meta-analysis found no such survival disadvantage in an analysis of eight published randomized controlled trials (4,651 patients) [131]. Overall, no difference was identified for the pooled clinically (odds ratio [OR] 0.92; 95% CI 0.76, 1.12) or microbiologically evaluable populations (OR 0.86; 95% CI 0.69–1.07) from the trials.
Synthetic lipoglycopeptides
This new group of semi-synthetic antibiotics is targeted specifically for gram-positive pathogens, particularly MRSA. They are rapidly bactericidal drugs, with the bacterial membrane as the site of action to depolarize and lyse the cell [132]. All have demonstrated in vitro activity against vancomycin-resistant bacteria. Telavancin has been approved for treatment of complicated SSTIs, having been shown to be non-inferior to vancomycin in the treatment of SSTIs at a single daily dose of 10 mg/kg [133]. As with all lipoglycopeptides, drug-associated adverse events are similar to those observed with vancomycin.
Dalbavancin has not been approved for any clinical indications at this time. The drug has a long elimination half-life that permits once-weekly dosing. One clinical trial of therapy of SSTIs with 1 g given on day one and 500 mg on day eight demonstrated non-inferiority compared with 14 days of linezolid treatment [134]. Oritavancin likewise is not approved currently for any clinical indication. As with dalbavancin, oritavancin has a long elimination half-life that may permit a single dose to be administered every 3–7 days. Unlike the other lipoglycopeptides, oritavancin is cleared primarily by the liver, which may obviate any reduction in dosing with renal insufficiency. Non-inferiority trials have been presented at clinical meetings, but the results have not been published.
Community-acquired MRSA antibiotics
As identified above, CA-MRSA commonly expresses the SCCmec type IV gene cassette in the USA300 strain, which does not have the genotype for resistance to selected non-β-lactam antibiotics. Thus, commonly used “old” and relatively inexpensive antibiotics still have a place in the treatment of these infections and should not be forgotten. Clindamycin, trimethoprim/sulfamethoxazole, doxycycline, and minocycline have in vitro and clinical experience, which indicates that these oral agents still have a role in the treatment of CA-MRSA soft tissue infections. Caution must be exercised with the apparent clindamycin-sensitive isolate that is erythromycin-resistant because of the risk of inducible resistance developing during therapy. The D-test is one method to identify and avoid this risk [135], and the other would be simply to choose another antibiotic (e.g., trimethoprim/sulfamethoxazole).
Iclaprim
Iclaprim is an advanced-generation dihydrofolate reductase inhibitor that has activity against a broad array of gram-positive organisms including MSSA, MRSA, VISA, VRSA, and VRE. It also has activity against Enterobacteriaceae and Haemophilus influenzae. Oral and intravenous preparations have been formulated. Iclaprim has been studied for the treatment of resistant gram-positive SSTIs [136], but has not been approved for use.
Cephalosporins
Despite the identity of MRSA as resistant to cephalosporins, ceftobiprole [137] and ceftaroline [138] are cephalosporin agents with binding affinity for the mutant penicillin-binding protein of MRSA. Ceftobiprole has activity against MRSA and most gram-negative and anaerobic species but no reliable activity against Pseudomonas aeruginosa. Published data demonstrate non-inferiority to vancomycin in complicated skin and skin-structure infections [139]. Ceftobiprole is approved in Canada for the treatment of complex SSTIs at a dose of 500 mg IV q 12 h, but it has not been approved for any indication in the U.S.
Ceftaroline has microbiological activity similar to that of ceftobiprole. Two randomized trials comparing ceftaroline with vancomycin in “complicated acute bacterial skin and skin structure infections” demonstrated a non-inferiority outcome [140,141]. Ceftaroline has been approved in the U.S. to treat “acute bacterial skin and skin structure infections” at a dose of 600 mg q 12 h.
Summary
Staphylococcus aureus is currently, and will remain in the future, a major pathogen among surgical patients. It offers a large catalogue of virulence factors that make infections a likely outcome following bacterial contamination of soft tissues, implanted and indwelling devices, and the respiratory tract. It demonstrates new patterns of infections (e.g., toxic shock syndrome and CA-MRSA SSTI). It was identified briefly 50 years ago as a cause of nosocomial enterocolitis [142], which may be re-emerging [143]. However, the most dramatic “changing face” of Staphylococcus aureus is the continued evolution of resistance to antibiotics. Resistant strains are identified soon after a drug is introduced into clinical practice, and evolution of resistance among the entire population of bacteria results in obsolescence of the drug over a period of years. Whereas the capacity of S. aureus to develop resistance is uncanny, the patterns of antibiotic utilization in clinical practice only fan the flames of evolution to resistant forms. Innovation in new drug development and other forms of treatment is needed urgently.
Author Disclosure Statement
Dr. Fry has received honoraria for being on the speakers bureau for Merck, and is a consultant for both Johnson & Johnson/Ethicon and Forrest Laboratories.