
Abstract
Abstract
Background:
Nosocomial infections represent an important problem for the health of hospitalized patients. Peri-operative infections—those occurring during surgery or in the post-operative period—account for 15%–20% of cases. Most surgical site infections (SSIs) are caused by endogenous gram-positive microorganisms, in particular, Staphylococcus aureus, S. epidermidis, and other coagulase-negative staphylococci that are part of the flora of the skin.
Methods:
A retrospective study was conducted from January 2006 to December 2010 to describe the epidemiology of methicillin-resistant S. aureus (MRSA) in SSIs. The MRSA isolates were analyzed by a combination of two genotyping methods: SCCmec and pulsed-field gel electrophoresis (PFGE). Also, biofilm-forming ability was analyzed for all isolates as an indicator of their ability to persist despite antibiotic treatment.
Results:
During the study period, 1,793 swabs from SSIs were analyzed, and S. aureus was identified in 318/987 positive specimens (32%). Methicillin resistance was observed in 10% of the S. aureus isolates (n=33). Analysis by PFGE revealed that isolates with the same SCCmec type were unrelated. Instead, biofilm-forming ability tests showed that SCCmec type I MRSA had the highest ability to form a film.
Conclusions:
The strains analyzed in our study showed a homogeneous pattern of SCCmec type. The difference in ability to produce biofilm between strains of SCCmec type I and isolates with other SCCmecs was substantial. This virulence factor could have critical implications for the formation and persistence of chronic SSIs.
Many reports indicate an increase in the incidence of MRSA skin and soft tissue infections in individuals with no apparent risk factors for hospital acquisition [7,8]. Pathogenesis of MRSA is attributable not only to antibiotic resistance, which complicates the treatment of infections, but also to other factors. One of these is the presence of genes for Panton-Valentine leukocidin (PVL), a bicomponent cytotoxin virulence factor associated with skin, soft tissue, and other more serious infections. The PVL factor is encoded by LukS and LukF and has been detected in both hospital and community MRSA isolates [9–11]. A second factor is the ability of this pathogen to adhere to medical devices and to form biofilms [12]. A biofilm creates a protected mode of growth that allows bacteria to survive in hostile environments protected from host defenses and the action of antibiotics. Bacteria in biofilms show high antibiotic resistance because of the difficulty the drugs have in penetrating the biofilm (antibiotics do not reach sufficient concentrations in any part of the biofilm) and the metabolism of bacteria at the base of the biofilm (they are metabolically inactive and therefore resistant to some antibiotics) [13].
However, although methicillin resistance is considered to be a prognostic factor, it remains unclear whether the virulence associated with MRSA is actually attributable to other microbiologic characteristics such as staphylococcal toxin production or biofilm-forming ability. The aim of this study was to determine, by SCCmec or restriction fragment typing, the epidemiology of MRSA involved in skin and soft tissue infection and to correlate some clones with the presence of other virulence factors such as PVL or biofilm-forming activity.
Materials and Methods
Bacterial strains
The work was performed on 33 MRSA strains isolated from infected surgical sites of patients admitted to the University Hospital “Federico II” of Naples over a period of five years. The isolation was done after incubation of the organisms on Trypticase soy agar (TSA) plates with 5% sheep blood (bioMérieux, Marcy l'Etoile, France) for 24 h at 37°C under aerobic conditions. Bacterial identification was performed on the base of biochemical tests carried out by the automated system Vitek II (bioMérieux). Stock cultures were kept at −80°C in Mueller Hinton broth (DIFCO, Detroit, MI) containing 20% glycerol. S. aureus ATCC 25923 was used as the reference strain (American Type Culture Collection, Manassas, VA).
Study of biofilm formation
The biofilm-producing ability was assessed using a quantitative method in microplates with spectrophotometer readings according to Stepanović et al. [16]. In brief, single colonies obtained from overnight cultures on TSA plates were used to prepare suspensions of MRSA in brain–heart infusion medium (BHI); (DIFCO) and incubated at 37°C for 18–20 h. The suspensions were read spectrophotometrically at 600 nm and diluted to obtain cultures of 0.1 OD. Next, 200 mcL of each culture was dispensed into a well of a 96-well microplate, then left without shaking at 37°C for 18–20 h. Subsequently, the wells were washed with 1×phosphate-buffered saline, stained with 200 mcL of crystal violet 0.04%, washed to remove excess dye, filled with 100 mcL of absolute ethanol, and read spectrophotometrically at 600 nm. For the results, we used the classification of adherence capabilities introduced by Stepanovic et al. [16] with modifications. Specifically, the cutoff ODc for the microtiter plate was defined as the OD of the negative control. The negative control was prepared by inoculation of 200 mcL of bacterial suspension inactivated by boiling. The isolates tested were classified into four categories: Non-adherent (NA), weakly adherent (WA), moderately adherent (MA), or strongly (A) adherent based on the OD of biofilm as follows: NA (OD≤ODc), WA (ODc≤OD≤2×ODc), MA (2×ODc≤OD≤4×ODc) and A (4×ODc≤OD).
Antimicrobial susceptibility tests
Antimicrobial susceptibility testing was done using the automated Vitek II system, and the results were interpreted using the EUCAST (European Committee) breakpoint criteria [14]. The antimicrobial drugs tested were amoxicillin, ciprofloxacin, clindamycin, erythromycin, gentamicin, oxacillin, rifampicin, teicoplanin, tetracycline, trimethoprim–sulfamethoxazole, azithromycin, clarithromycin, quinupristin-dalfopristin, linezolid, and vancomycin. Resistance to methicillin was determined by the oxacillin (5 mcg) disk susceptibility test. To determine the antimicrobial susceptibilities of sessile cells, we induced biofilm formation as described in the previous section, and then added 200 mcL of BHI medium containing serially double-diluted concentrations of each antimicrobial agent, and the plates were incubated for a further 24 h at 37°C. The positive control included antimicrobial-free wells, and the negative control boiled bacterial cells. After incubation, the medium in excess was discarded, and biofilm formation was quantified adding a 100 mcL aliquot of XTT-menadione (0.1 mg mL−1 of 2,3-bis (2-methoxy-4-nitro-5-sulfo-phenyl)-2H-tetrazolium-5-carboxanilide (XTT) and 1 mcM menadione) to each well. The plates were incubated in the dark for 2 h at 37°C and then read spectrophotometrically at A490 with a microtiter plate reader (Bio-Rad, Marne-la-Vallée, France). Sessile minimum inhibitory concentrations were determined at 50% (MIC50) inhibition compared with antimicrobial-free control wells. Experiments were carried out in triplicate.
Extraction of genomic DNA and PCR
Genomic DNA was purified using the DNeasy Blood and Tissue Kit (Qiagen, Germantown, PA) according to the manufacturer's instructions and was evaluated spectrophotometrically and checked by electrophoresis on 1% agarose gel before amplification.
Two multiplex PCRs, designed to identify the type of ccr gene complex (1–5) and mecA gene class (A–C), were performed according to the procedure of Kondo et al. [15]. The primer pairs used are listed in Table 1. For identification of the LukS and LukF PVL genes, the amplification reaction was carried out using the primer pairs listed in Table 1. For all amplifications, the reaction mixture contained 10 ng of genomic DNA, 0.2 mcM each primer, dNTP 200 mcM each, 3.2 mM MgCl2, and 2.5 U of Taq polymerase. The program included the following phases: Initial denaturation at 94°C 10 min; 35 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min; and elongation at 72°C for 1 min with final elongation at 72°C for 10 min. S. aureus ATCC 25923 was used as the negative control of both reaction mixes.
Pulsed-field gel electrophoresis (PFGE)
The overnight cultures were harvested by centrifugation at 4000×
Statistical analysis
The statistical differences in biofilm formation between strains with different SCCmec types were calculated using the χ2 test with Yates correction for continuity. Distributions were considered significantly different at p<0.05.
Results
In a period of five years between January 2006 and December 2010, 1,793 swabs from surgical sites were analyzed in the Functional Area of Microbiology of the University Hospital “Federico II.” Of these, 987 swabs were positive for bacteria. Species identification was performed by the Vitek II automated system (bioMérieux).
The species isolated most commonly was S. aureus (318/987; 32%) of which 10% were resistant to methicillin. Many isolates originated from orthopedic device-associated site infections (6/33) and vascular device-associated site infections (6); the other isolates originated from abdominal (6), dermatologic (4), and other non-device-associated site infections (11).
Each MRSA strain gave a reproducible and well-separated banding profile containing 13–17 fragments ranging from 50 to 10,000 kilobases. Strains were considered to belong to the same PFGE type (possibly related) if they were characterized by as many as a six-bands difference in the macro-restriction band patterns according to the Tenover et al. criteria [17] and by a cutoff similarity value of 90%. The molecular characterization by PFGE classified the 33 strains into 29 types with high diversity. Only four PFGE types (C, D, G, and N) consisted of two possibly related strains (Fig. 1).
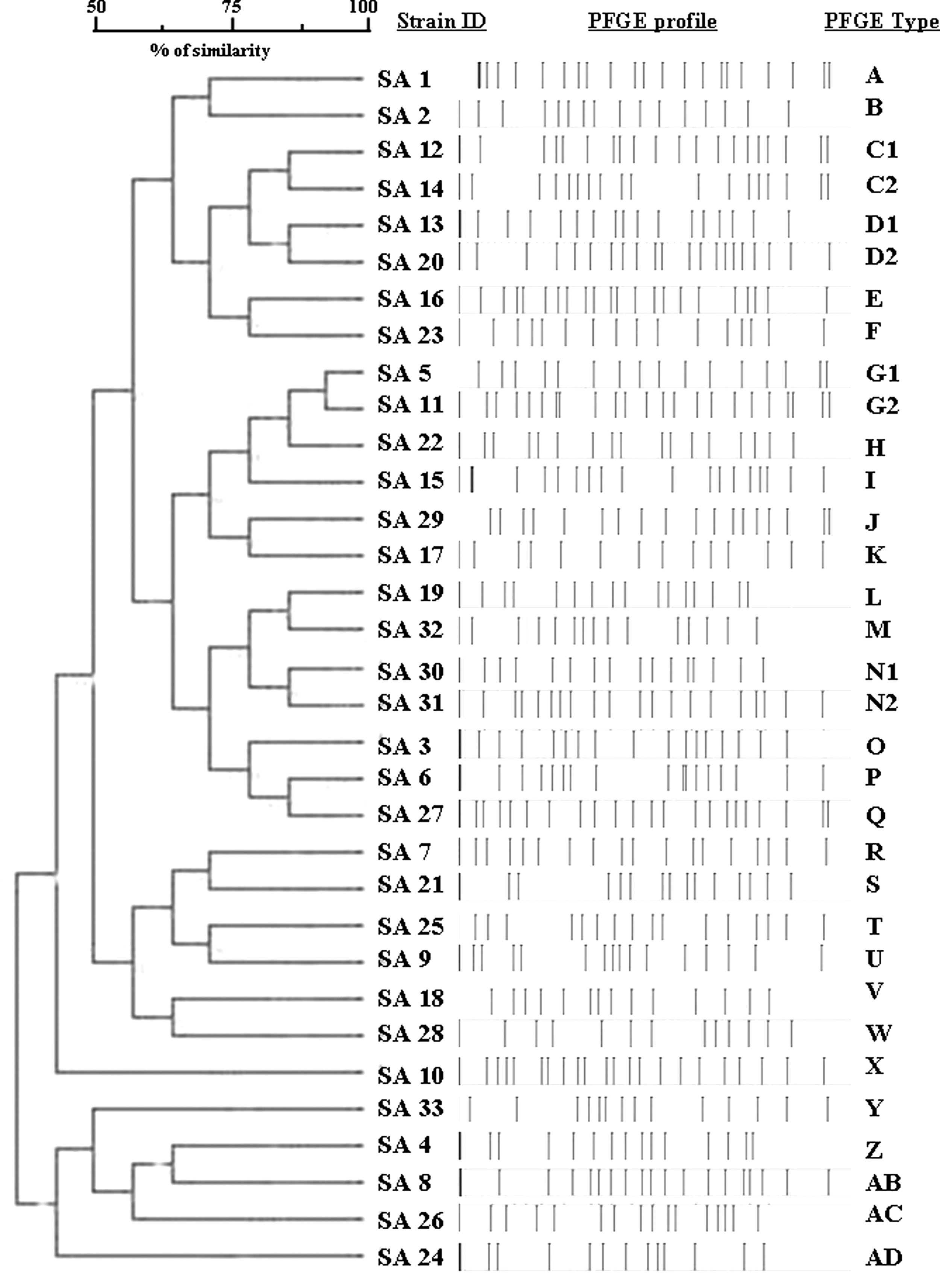
Schematic representation of SmaI digestion of MRSA strains and dendrogram showing clusters. Pulsed-field gel electrophoresis types are indicated by letters and numbers and are defined at a cutoff value of 90%.
For each strain, the SCCmec type was obtained by the combination of the mec complex class and the ccr gene complex. Table 2 summarizes the results. The SCCmec cassette type I was found in 16 strains, followed by type IV (six strains), type III (five strains), and type II (four strains), whereas type V was found in a single bacterial strain. A strain also was found that carried two boxes: Types V and III simultaneously.
A=strongly adherent; AMX=amoxicillin; AZM=azithromycin; ATCC=American Type Culture Collection (reference strain); CIP=ciprofloxacin; CLR=clarithromycin; CLI=clindamycin; ERY=erythromycin; G=gentamicin; MA=moderately adherent; NA=non-adherent; O.D.=optical density; OXA=oxacillin; PVL=Panton-Valentine leukocidin; RIF=rifampicin; TET=tetracycline; STX=trimethoprim/sulfamethoxazole; T=totally drug resistant; WA=weakly adherent.
The antibiotic sensitivity of all 33 strains was tested (Table 2). All strains were resistant to penicillins and to first- and second-generation cephalosporins; the strains were susceptible to teicoplanin and vancomycin, in addition to presenting sensitivity to linezolid and quinupristin-dalfopristin.
The analysis of biofilm formation showed that 22 strains (67%) were strong or moderate biofilm producers (Table 2). Most of them (15/16) were characterized as SCCmec type I. Statistical analysis showed a significant difference (p<0.05) between the amount of biofilm produced by strains having SCCmec I and those with other SCCmecs.
The biofilm sensitivity was tested for each isolate that showed biofilm-forming ability. There was an increase in the resistance of each isolate to all tested antimicrobial agents, although isolates that formed strong or moderate biofilm showed higher MICs than other ones (Table 2).
Finally, the strains were tested for PVL (Table 2). Of the 13 strains, five carried PVL; four of them belonged to SCCmec type IV and one to SCCmec type I.
Discussion
Prevention and control of nosocomial infections is a priority health goal to achieve a reduction of infections associated with invasive procedures, in particular operative ones. European hospital epidemiology investigation showed that S. aureus is the microorganism isolated most frequently in the course of SSIs [18–21]. Saini et al. [22] also identified S. aureus as the principal microorganism responsible for infections of orthopedic injuries, attributing them to the invasive properties of the bacteria and their presence in the normal flora of the skin, which can easily be inoculated into the surgical site.
MRSA is a major health problem: The incidence of nosocomial infections is increasing steadily, although there are differences between countries, and now MRSA has become an important pathogen that also can be acquired in the community setting.
In our study, 1,793 swabs of surgical sites were analyzed, evidencing S. aureus and coagulase-negative staphylococci, microorganisms belonging to the usual flora of the skin as among the most frequently isolated organisms. Strains of MRSA were responsible for SSIs in 3.3% of cases, in disagreement with data presented by Villari et al. from the same university hospital. However, those investigators collected isolates mainly from outbreaks in intensive care units [18]. Analysis by PFGE showed different pulsotypes, although the isolates of the same PFGE subtypes were found in the same unit during different periods during the five-year study. With the exception of these isolates, a unique PFGE type was associated with each patient, showing the low clonality of the MRSA strains responsible for SSIs.
The MRSA strains tested in this study had, for the most part, cassette type I (16 strains of the 33 considered). Cassettes I and II are found mostly in strains of nosocomial origin; they are larger than cassettes III, IV, and V, which are found mainly in community strains and carry multiple antibiotic resistance genes. Thus, these organisms are resistant, not only to β-lactams, but also to other families of antibacterial drugs, making their eradication difficult.
The strains analyzed in our study had heterogeneous patterns of antibiotic resistance, as there were strains with SCCmec type IV, which showed resistance to third-generation cephalosporins, tetracycline, and azithromycin; and strains with SCCmec type I or III, which were sensitive to many antibiotics. The identical pattern of susceptibility was reported by Crivellaro et al. [23] for 90 MRSA isolates collected in the same period in a North Italy hospital, but the majority of those isolates harbored SCCmec type II. The pattern of susceptibility was comparable also to that observed by Campanile et al., although in our study we did not observe decreasing susceptibility to linezolid [24].
Only five of the 33 strains possessed the LukSF gene, one of which was characterized by SCCmec type I, contrary to what typically is reported in the literature, but what is in accord with more recent data [10,11]
Two thirds of the strains were strong or moderate biofilm producers. As many as 15 of these 22 strains (68%) were carriers of SCCmec type I. Statistical analysis indicated that the difference between strains with SCCmec type I and isolates with other SCCmecs is substantial in terms of the ability to produce biofilm, showing that MRSA strains that carry SCCmec type I form biofilm to a greater degree than do the MRSA strains with different types of cassettes. Most biofilm producers were isolated from orthopedic (13/23) and vascular (5) sites, particularly related to implants (13). Biofilm production influences antibiotic resistance by increasing MICs for most antimicrobial agents [25]. In our study, antimicrobial sensitivity testing of planktonic and sessile cells for each isolate revealed the expected increase in MIC. Although the increase for moderately and strongly adherent isolates was higher than for weakly adherent ones, the shift of these isolates in the resistance category made eradication equally difficult.
Kaito et al. identified the PSM-mec cytolytic peptide on the mobile genetic element SCCmec as altering the virulence properties of MRSA through increases in the amount of biofilm formed [26]. Gill et al. showed that there are 140 genes associated with the severity of infection present in regions near the SCCmec cassette, evidencing a possible co-evolution of SCCmec genes and the region of the MRSA genome [27]. According to these data, and as stated by Manago et al. [28], we suggest that association between biofilm formation and HA-MRSA could encourage chronic colonization by these strains.
Our results, showing that MRSA strains had a different ability to form biofilm depending on the type of SCCmec, suggest that strains with SCCmec type I have greater pathogenicity linked to biofilm formation. Thus, this virulence characteristics without being the most important, may contribute to the pathogenicity of the bacterium, and therefore to the outcome of an infection.
Unfortunately, most of the patients enrolled in the study were highly compromised, and for this reason, it was not possible to relate the virulence of microorganism to the clinical outcome. Future studies will aim to deepen the relation between genotypic and phenotypic aspects (i.e., SCCmec and biofilm formation) to the virulence of the strains.
In conclusion, it could be useful to analyze the SCCmec type of MRSA isolates to predict, not only the probable antimicrobial susceptibility but also the ability to form biofilm and then grow protected from the immune response and antibiotics. It would be helpful to know these virulence characteristics, not essential to causing SSIs directly but capable of influencing the outcome of infections.
Author Disclosure Statement
No competing financial interests exist.