
Abstract
Abstract
Background:
Infections and inflammation of the lower limb skin, soft tissues, and vessels are more common than in other body regions. The aim was to determine whether cryptic bacteria dwelling in deep tissues are the cause.
Methods:
We performed bacteriologic studies of specimens harvested from arteries of amputated ischemic legs, leg varices, and tissue fluid/lymph and lymphatics in lymphedema.
Results:
Calf arteries contained isolates in 61% and femoral arteries in 36%, whereas normal cadaveric organ donors' arteries in 11%. Bacterial deoxyribonucleic acid (DNA) was detected in 70%. The majority of isolates belonged to the coagulase-negative staphylococci and Staphylococcus aureus; however, highly pathogenic bacteria were also detected. All were sensitive to all antibiotics except penicillin. Saphenous vein varices contained bacterial cells in 40% and controls 4%; bacterial DNA was found in 69%. The majority of bacteria were S. epidermidis and S. aureus susceptible to all antibiotics except penicillin, Lymph and epifascial lymphatics limb contained bacteria in 60% and 33% samples, respectively and controls in 7%. Most were S. epidermidis susceptible to all antibiotics except penicillin.
Conclusion:
Cryptic bacteria are present in lower limb tissues and may play a pathologic role in surgical site infections. Proper antibacterial prophylaxis should be considered when planning surgical interventions.
I
The presence of cryptic microbes in healthy animal tissues has been documented widely [8–11]. Dormant clostridial spores were found in healthy skeletal muscles and were responsible for myositis or myonecrosis [12]. A number of cryptic bacteria activated after injury were isolated from normal human soft tissues [13–16]. Dormant microbes were identified in calluses of closed tibia and femur fractures during operations for non-alignment [17]. Bacteria were detected in the peri-arterial tissue of tibial, popliteal, and femoral arteries in critical limb ischemia without peripheral necrosis [18]. Specimens of thrombotic fragments of saphenous vein obtained from limbs without ulcer revealed presence of bacteria [19]. The harvesting of normal great saphenous vein for aorto-coronary bypass is frequently complicated by infection and delayed incision healing even though taken under strictly aseptic conditions [20–22]. Dermato-lymphangio-adenitis (DLA) caused by staphylococci develops frequently in limbs with lymphedema with no evident ports of entry of microbes [23].
Bacteria dwelling in tissues may have entered through the breached epidermis but they also could have been translocated via blood circulation from the gut, lungs, or infective sources. The question remains as to whether these dormant bacteria play a role in the development of necrosis in ischemic legs, venous ulcers, and recurrent attacks of DLA in lymphedema.
The response of the host depends on the mass of penetrating, accumulated microbes. Single bacterial cells or biofilms may persist without host reaction. The threshold value is 105 bacteria per gram of tissue. However, under pre-disposing conditions such as venous stasis, arterial compromise, lymphedema, and neuropathy as well as systemic conditions as diabetes mellitus, central nervous system compromise, trauma, inflammatory illnesses, metabolic abnormalities, coagulopathies, immunosuppression, malnutrition, and obesity, activation of the dormant bacteria may occur [23]. The consequence is inflammation, ulceration, and necrosis of tissues.
In the search for cryptic bacteria, we performed bacteriologic studies of tissue specimens harvested from (1) arteries of amputated critically ischemic legs without skin ulcers or necrosis; (2) non-thrombosed varices of the great saphenous vein; and (3) tissue fluid/lymph and lymphatic vessels in obstructive lymphedema without clinical signs of tissue inflammation. There were no detectable ports of entry of microbes from external environment as surgical site infections, ulcers, or necrosis.
Patients and Methods
Specimens for bacteriologic studies were collected from three groups of patients: Lower limb ischemia, lower limb venous insufficiency with varices, and lower limb obstructive lymphedema. All patients were admitted to the vascular surgery ward for treatment in the order in which they presented to the outpatient clinic.
Limb arterial ischemia
Group 1 comprised 60 patients, ranging in age from 50 to 80 y (mean, 68 y). There were 18 females and 42 males, with atherosclerosis of lower limb arteries causing chronic exacerbated foot ischemia, Rutherford scale 4 [18]. Inclusion criteria were intractable ischemic foot pain (42 patients) and dry necrosis of a toe at an initial stage (18 patients). Obstruction of superficial femoral, popliteal, and calf arteries was diagnosed on arteriography. In all patients the arteriographic image precluded reconstructive operation and amputation was performed below the knee or at the mid-thigh. Fragments of vascular bundles containing arteries were harvested during surgery. All patients received broad-spectrum antibiotics for at least 7 d before amputation. Excluded were patients with overt diabetes mellitus and foot necrosis. In group 2, normal lower limb vascular bundle fragments were collected from 27 cadaveric organ donors under strictly sterile conditions in the operating room.
Limb superficial varices
Group 1 comprised 100 patients, age range 25 to 78 y (mean 49 y), 68 females and 32 males, with varicose veins (C4:C4a pigmentation or eczema, C4b lipodermatosclerosis according to the CEAP classification) of 5 y duration. Excluded were subjects with ulcer healed less than 3 y ago, limb ischemia, and those with diabetes mellitus. In group 2, 25 control specimens included great saphenous and femoral veins harvested from cadaver organ donors in the operating room.
Limb lymphedema
Group 1 was composed of 54 patients with obstructive lymphedema stage II and III. The etiology in 38 patients was a soft tissue bacterial inflammatory episode at least 3 y ago and hysterectomy and radiotherapy because of cervical cancer in 16. Duration of edema was more than 3 y. Excluded were patients with recent DLA, ulcers, venous insufficiency, ischemia, and abrasions of limb skin. Stasis was documented by lymphoscintigraphy. Control group 2 consisted of 30 healthy subjects participating in studies on reverse cholesterol transport from limb tissues via afferent lymphatics to blood circulation.
Harvesting material
Limb skin was disinfected in all groups with 70% isopropyl alcohol, and fragments were taken for bacteriologic investigation, to exclude contamination during harvesting.
Arteries
Fragments of the posterior tibial, popliteal or femoral artery and vein bundles, and calf epifascial lymphatic vessels were harvested in the operating room from the disinfected amputated calf and thigh stumps. In the control groups, fragments of the femoral artery were harvested from the cadaveric organ donors. Specimens were divided into two fragments: One for the bacteriologic culture, another for the bacterial DNA. Material for bacteriologic studies was placed in transport media, for microbial DNA it was frozen at −70°C in dry ice and stored until further use.
Superficial veins
Specimens were obtained during routine varicose vein surgery. Skin was disinfected three times, allowed to dry, and covered with Tegaderm® adhesive sheeting (3M, St. Paul, MN). An incision was made and a swab was placed in the incision for 1 min. A 1 cm long fragment of varix was excised and placed in culture medium.
Epifascial lymphatics
Specimens were obtained during routine biopsies for histologic staging of lymphedema. Skin was treated in the same manner as for harvesting of veins. Patent blue solution was injected into the foot toe-web to colorize epifascial lymphatics. Foot and calf massage allowed the dye to move to the thigh. One centimeter long fragments of superficial lymphatic collectors running along the great saphenous vein were excised.
Tissue fluid and lymph
Tissue fluid/lymph were collected during routine skin biopsies. Skin was treated in the same manner as for harvesting of veins. Tissue edema fluid spontaneously filling the incision was collected in sterile syringes and 3–5 mL were injected into bottles containing Hemoline medium (bioMérieux S.A., Marcy l'Etoile, France). In some cases lymphatic cannulation was performed as described elsewhere [24]. In the control group, lymph was harvested in normal subjects in the reverse cholesterol transport program. Obligatory bacteriologic culture had to be performed. The obtained volume was injected into bottles containing Hemoline medium.
All specimens were collected by the same surgeon, under strict sterile conditions, in the operating room. Batches of all instruments and material used for collection of the specimens were tested for sterility prior to their use in the study. Control swabs were taken from the disinfected skin and surface of scalpel after collection of arteries, veins, lymphatics and tissue fluid and lymph. Studies received consent of the Warsaw University medical ethics committee. Informed consent was signed by each patient before limb amputation and tissue biopsies.
Bacteriologic culture media
The following media were used: Hemoline liquid medium, Columbia blood agar base enriched with 5% sterile defibrinated sheep blood, MacConkey's agar, Chapman's agar, Sabouraud's agar (malt agar), and brain heart infusion (BHI; all from Difco, Detroit, MI).
Identification of bacterial strains
The cultures were incubated at 37°C and examined at 24 h and 48 h for aerobic bacterial growth. After 48 h of incubation, BHI cultures were transferred into blood agar, and examined after another 24 h culture. Biopsy material was transferred from transport media to BHI medium. These cultures were examined every day for up to 10 d and, if positive, inoculated onto blood agar slants. After 10 d, all negative cultures were transferred to blood agar. In cases in which there was no aerobic growth, additional cultures for anaerobic growth were established. Isolates were identified by standard procedures using the Analytical Profile Identification (API) System (Biomerieux). Aerobic cocci of the Micrococcaceae family were identified using the API-Staph system. Bacteria of Streptococcaceae family were identified with API 20 Strep. Members of the genus Corynebacterium (gram-positive coryneform and pleomorphic bacilli) were identified with API Coryne. Gram-negative bacilli were identified using API 20E. Gram-positive spore-forming bacilli (Bacillus spp.) were identified by evaluating the fermentation of sugars or polyalcohols. The susceptibility of isolated bacterial strains to antibiotics was examined using the ATB system (bioMerieux). Analysis of the antibiotic sensitivity was performed using the ATB-Plus reader (bioMerieux).
In another test, fragments of arteries and veins and adjacent tissues (fat, muscles) were placed on Hemoline plates and cultured for 7 d at 37°C. Bacteria migrating from the tissue and forming colonies were characterized under the microscope (magn. x 100), then transferred to differential media. This method allowed the optical evaluation of migration activity of different strains, density of colonies and competition between strains. Comparison of bacterial strains observed on the Hemoline plate with those from the fragments placed in routine transport media was made.
Identification of bacterial DNA-16sRNA
The following materials were used: Genomic DNA isolation kit (Macherey-Nagel GmbH Co., KG. Düren, Germany), polymerase chain reaction (PCR) kit (Sigma, St. Louis, MO), RedTaq Genomic polymerase (Sigma), Primers Oligo (IBB, Warsaw, Poland), polyacrylaminde gel electrophoresis (PAGE; Amersham Pharmacia Biotech, San Francisco, CA), and DNA Silver Staining Kit (Amersham). The method of extraction of DNA was performed using the DNA isolation kit (Macherey-Nagel) according to the manufacturer description. The PCR amplification was performed with primers for gene fragment coding bacterial 16s RNA: 5′AGTTTGATCCTGGCTCAG 3′ forward and 5′GGACTACCAGGGTATCTAAT 3′ reverse to detect any bacterial strain. Reactions were performed in MJ Research thermocycler in the following conditions: 95°C- 5′; 95°C- 45′′, 60°C- 45′′, 72°C- 1′- 35 cycles; 72°C- 10′. Products were separated by PAGE electrophoresis and silver stained. Products of interest were identified by comparison with the marker-100 bp ladder. The expected length of amplification product (base pairs) of 16 s RNA gene was 789 bp.
Data statistical evaluation
The frequency of bacterial isolates was presented in percentage of positive cultures. For evaluation of statistical differences between the numerical prevalence of bacterial isolates and PCR products in groups the Student t test for unpaired data was applied. A p value <0.05 was considered statistically significant.
Results
Bacteriology of disinfected limb skin
The disinfected skin specimens did not reveal presence of bacterial isolates in 96% of cultures. S. epidermidis and other coagulase-negative staphylococci were detected in 4%. This low number precluded the possibility of contamination of the harvested vessel specimens.
Bacterial isolates in arterial walls
In group 1 (60 ischemic limbs) specimens of tibial and popliteal arteries contained bacterial cells in 60.6% and femoral arteries in 30.8% (Figs. 1 and 2). In the healthy femoral arteries, microbial cells were isolated in 11% (ischemic versus controls, p<0.05). The isolates in limb arteries belonged in 71% to the coagulase-negative staphylococci and S. aureus. There were also highly pathogenic Enterococcus, Proteus, Pseudomonas, Micrococcus, Klebsiella, Enterobacter, Serratia, Acinetobacter, and Citrobacter (Table 1). The gram-positive bacteria were susceptible to all antibiotics but penicillin. Enterococcus was sensitive to vancomycin (Table 2) Topographic distribution of bacterial strains in various limb tissues are presented on Figures 1 and 2.
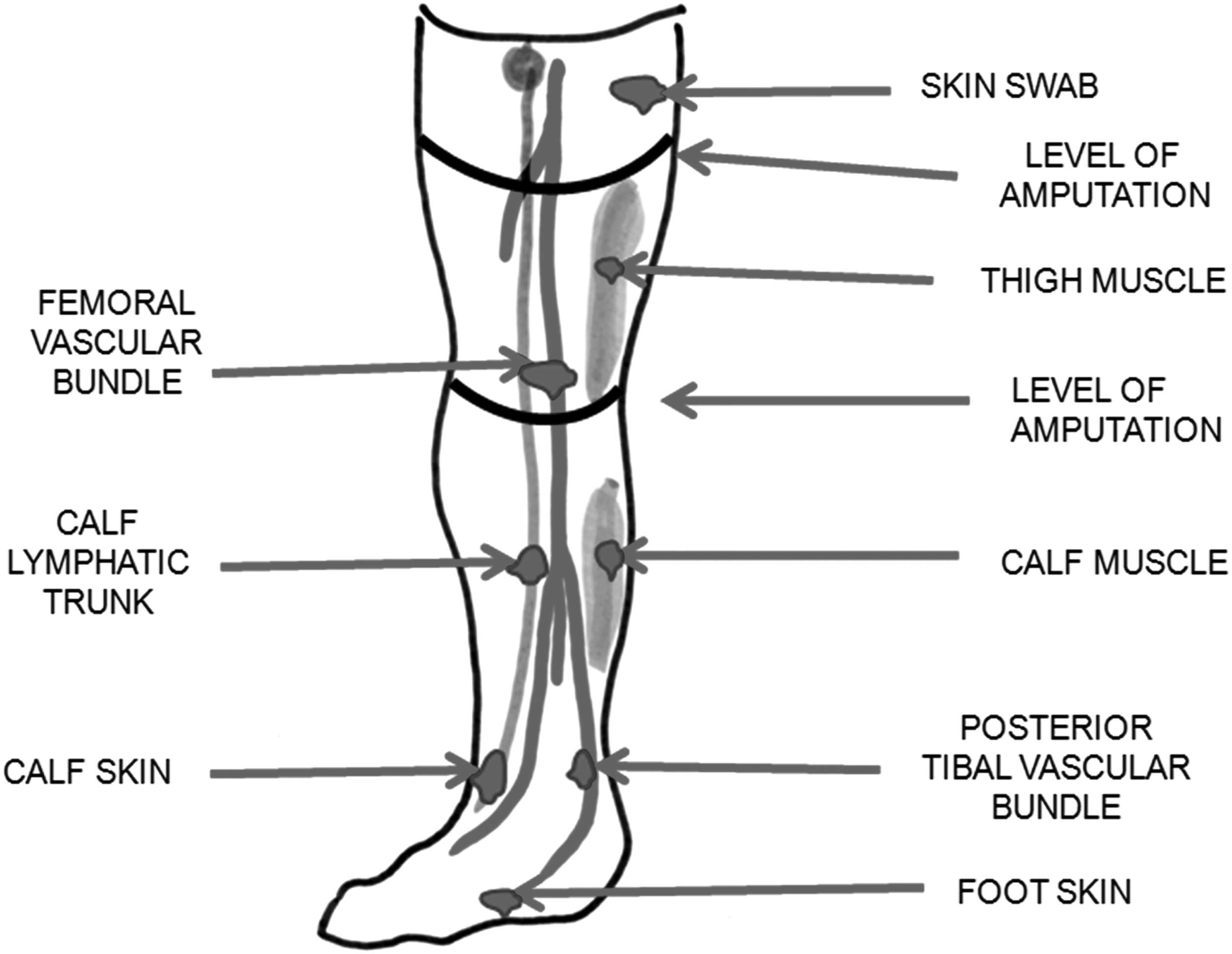
Topography of tissues harvested from an ischemic lower limb for bacteriologic studies.
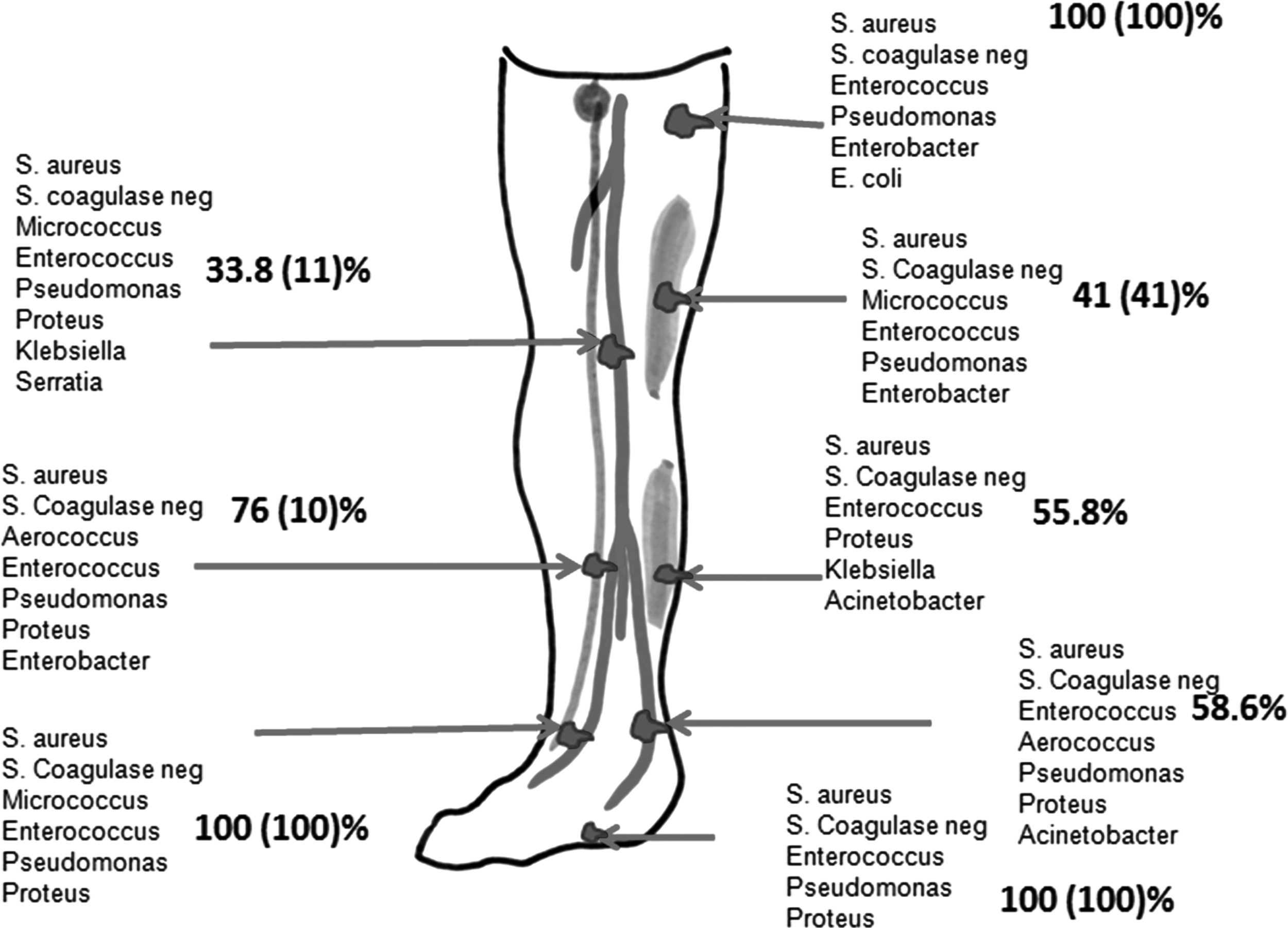
Topography of distribution of bacterial isolates in tissues of ischemic lower limb. Numbers represent bacterial prevalence of strains listed below. Note that bacteria were found in all deep tissues. Skin bacterial fragments were taken before disinfection.
Values of healthy cadaver organ donors are given in parentheses (n=27) (in percent).
Calf vs. foot or thigh p<0.05.
Ischemic vs normal tissues p<0.05.
Eleven patients, 26 specimens, 27 bacterial isolates.
Optical evaluation of colonies formed by migrating bacteria revealed dominance of staphylococci (Fig. 3). They were present in arteries, muscles, and subcutis. Single colonies of highly pathogenic bacteria, as listed above, could also be observed in some specimens. Interestingly, microbes were also present in the bone marrow.
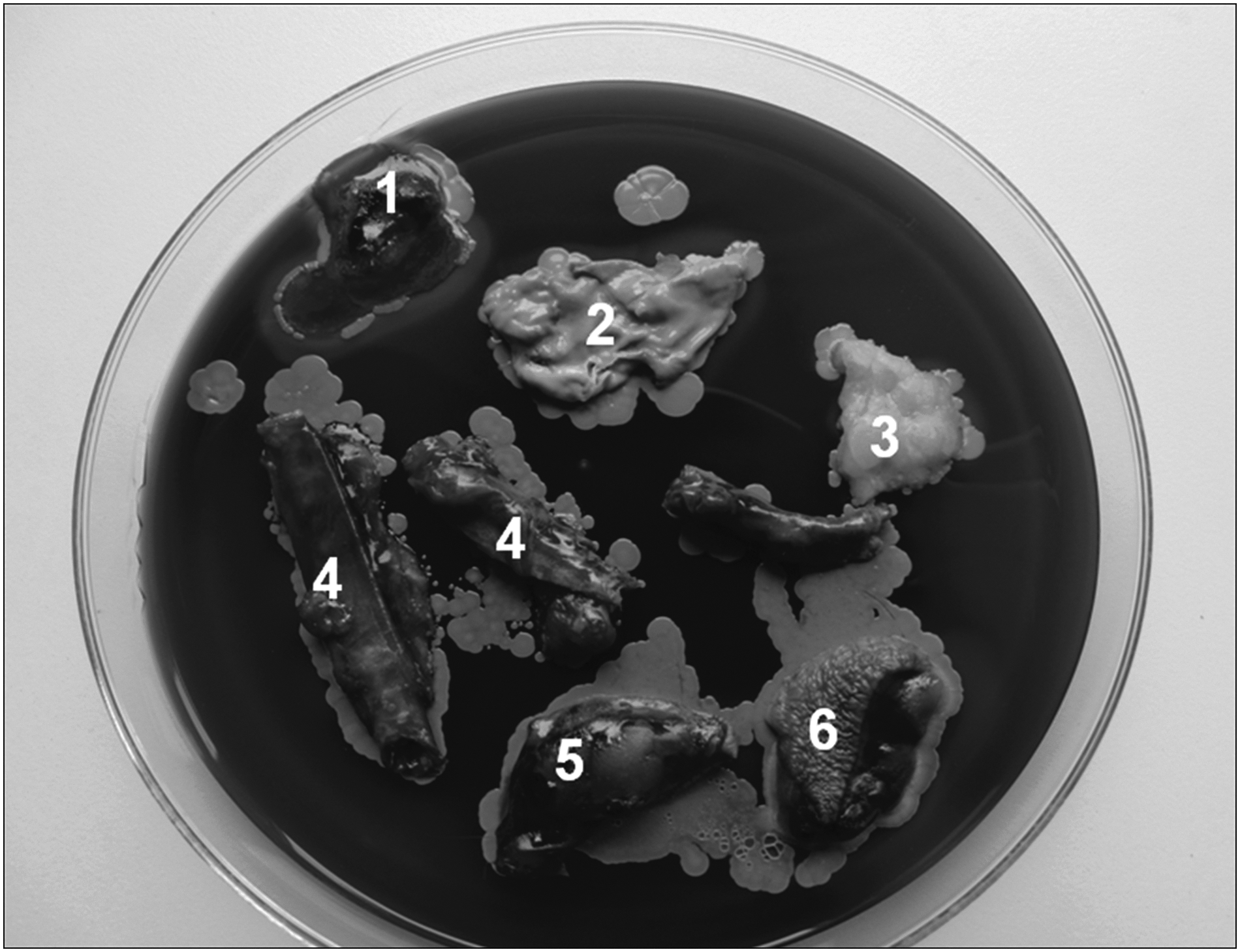
Fragments of tissue harvested from ischemic upper calf. (
Microbial DNA in arterial wall
In group 1, of 60 samples of tibial, popliteal, and femoral arteries, the 16s RNA gene was detected in 70%. Twenty-five normal femoral arteries revealed the presence of 16s RNA in 21%.
Bacterial isolates in varicose veins
Varicose vein specimens revealed the presence of bacterial isolates in 40%, whereas controls taken from healthy cadaveric organ donors contained live bacteria in only 4% (Tables 3 and 4). Disinfected skin specimens from the sites of varicectomy showed the presence of microbes in 4%. The dominant isolates from vein specimens were staphylococci, preponderantly coagulase negative, however, in a few cases Enterococcus faecium was also detected. Staphylococci were highly sensitive to antibiotics with the exception of penicillin (Table 5). Thirty-three percent of isolates were methicillin-resistant. Bacterial culture on the Hemoline plates revealed microbes migrating from the outer aspect of varices and adjacent fat but not muscles (Fig. 4). The 16s RNA was detected in 69% of specimens, evidently greater than the percentage of live bacterial cells.

Fragments of varicose great saphenous vein and adjacent tissues. (
GSV=great saphenous vein.
n=40 specimens (in percent).
Bacterial isolates from leg lymph and lymphatics in lymphedema
Cocci were isolated in 60% of lymph samples with S. epidermidis dominating and occasionally S. aureus (Tables 6 and 7). Lymphatics and nodes contained bacteria in 33%. In the normal leg lymph, S. epidermidis was detected in 12% of samples. The lymph and lymph nodes isolates, both from lymphedema and normal subjects, were sensitive to most antibiotics (Tables 8 and 9). Bacteria present in lymph were mostly coagulase-negative staphylococci and S. aureus, with few Enterococci (Fig. 5). Surprisingly, the least sensitivity showed microbes to penicillin, although this antibiotic turned to be effective in prevention of DLA attacks in a long-term administration protocol.
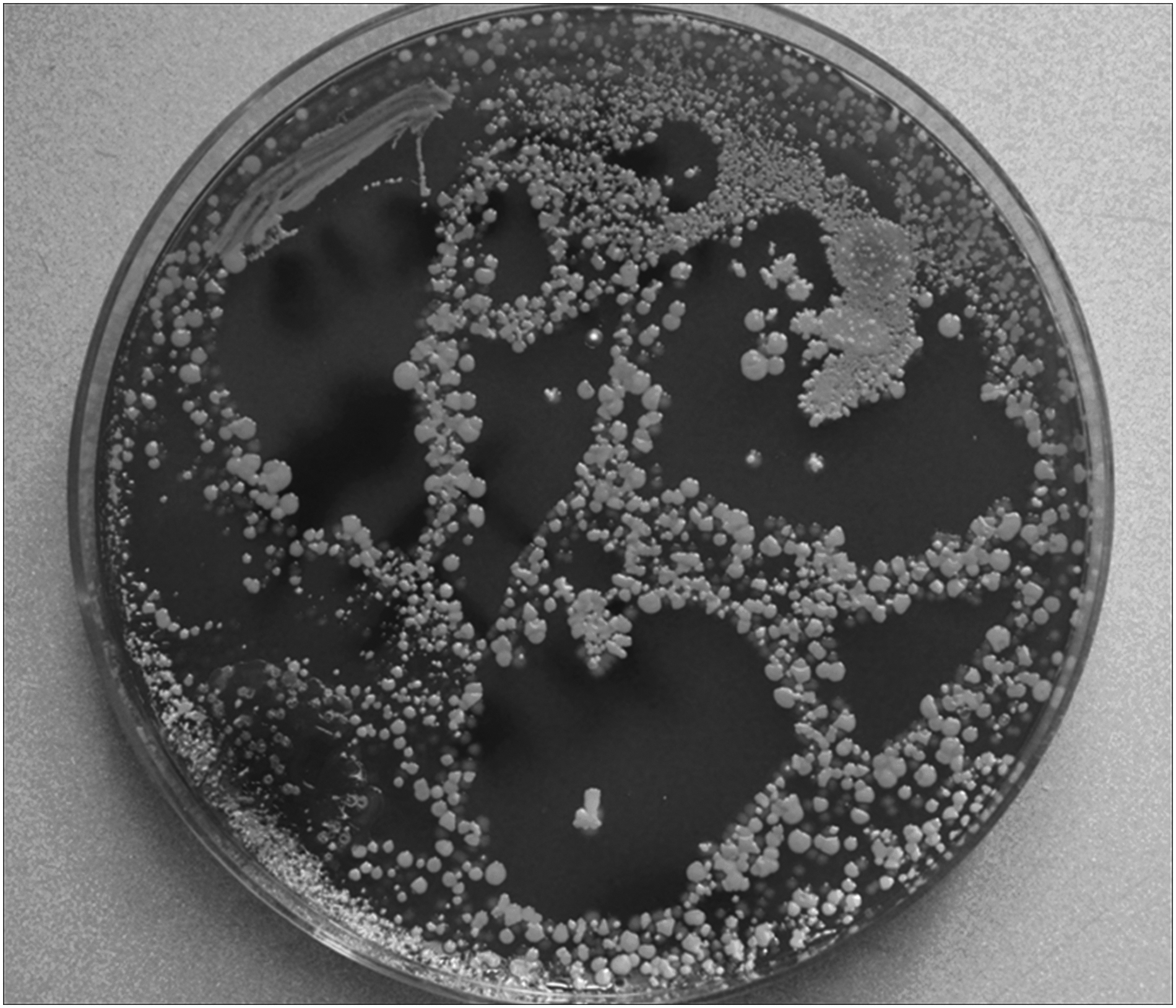
Lymph sample from lower limb during acute dermato-lymphangio-adenitis was spread on the plate. Multiple colonies of S. aureus and coagulase-negative staphylococci, few enterococci.
Values from 30 healthy volunteers are in parentheses.
p<0.05
Percent of isolates.
Percent of isolates.
Discussion
Calf arteries of ischemic limbs contained bacterial isolates in 61% and femoral arteries in 36%, whereas, normal cadaveric organ donors' arteries were positive in 11%. Bacterial DNA was detected in 70% of ischemic limb arteries. The isolates belonged in the majority to the coagulase-negative staphylococci and S. aureus, however, highly pathogenic Enterococcus, Proteus, Pseudomonas, Micrococcus, Klebsiella, Enterobacter, Serratia, Acinetobacter, and Citrobacter were also detected. They were sensitive to all antibiotics except penicillin. Saphenous vein varices contained bacterial cells in 40% of specimens and bacterial DNA in 69%. The majority of bacteria were S. epidermidis and S. aureus susceptible to all antibiotics except penicillin. Lymph and epifascial lymphatics from lymphedematous limb contained bacteria in 60% and 33%, respectively. Control samples contained bacteria in 7%. Most bacteria were S. epidermidis susceptible to all antibiotics but not penicillin.
We documented the presence of bacteria in the vascular tissue of lower limbs in the three most common vascular conditions: Atherosclerosis of arteries with foot ischemia, varicose veins, and post-inflammatory obliteration of lymphatics. In all cases limb skin remained intact. There were no ulcers or necrosis in the past history. The differences in frequency of isolates between diseased tissues and healthy controls were highly statistically significant. This suggests prior colonization of the pathologically changed arteries, veins and lymphatics by bacteria, most likely from the foot. Interestingly, the frequency rate of live bacterial cells was similar in all these vessels. Moreover, there was full susceptibility to all antibiotics with the exception of penicillin. This may point to indigenous commensal colonization of lower limb deep tissues and lack of host reaction. However, under conditions of ischemia or blood or lymph stasis, these microbes may become activated and evoke host inflammatory response. External contamination during harvesting of specimens has been excluded because samples were taken in the operating room under strict aseptic conditions. Moreover, cultures of instruments used for harvesting were proved sterile.
Lower limbs are exposed to colonization of their deep tissues in the process of physiologic penetration of micro-organisms through the normal sole skin, as well as during foot epidermal abrasions or micro-injuries. The penetrating microbes are transported away from the ports of entry via lymphatics to the lymph nodes. The low bacterial load transported in this fashion is clinically hardly recognizable. However, bacteriologic culture of lymph remains helpful in this respect allowing to isolate microorganisms from lower limb lymph in approximately 5%–10% of healthy individuals [23]. Because limb deep lymphatics run in close proximity to vein and arteries and the lymphatic endothelium has the ability to transport particles adluminally and abluminally, there exists the possibility of infecting arterial walls at the predisposed sites. The foot bacterial flora consists of indigenous microbes physiologically residing on the epidermal surface, in hair follicles, and glands. In addition, there are bacteria originating from the perineal and anal regions populating skin on desquamated epidermal scales. These microbes may be highly pathogenic in limb ischemia, venous thrombosis or lymphedema [25,26] and add to the already existing inflammatory changes.
Although there is a plethora of articles on the role of infective factor in the pathogenesis of atherosclerosis in the coronary, carotid and subclavian arteries and thoracic and abdominal aorta only few papers are dealing with identification of bacterial cells or DNA in arteries of lower limbs [27]. The presence of Chlamydia pneumoniae, Helicobacter pylori, herpes viruses such as cytomegalovirus (CMV) and herpes simplex virus (HSV), as well as odontopathogens Porphyromonas gingivalis and and S. sanguis in atherosclerotic plaques of coronary and carotid arteries and aortic wall has been documented widely both using PCR and serologic methods [28–30].
We reported recently about the presence of foot skin-derived bacteria in arterial walls [18]. Our present results confirm previous data on the type and frequency of those nosocomial microorganisms. Co-influence of multiple strains should be taken into consideration. These infectious agents may influence vascular cell functions by inducing thrombus formation, vascular cell proliferation, apoptosis, and cell death [31].
The potential mechanism of infection-induced atherosclerosis remains speculative. Infective factors can either directly or indirectly influence the process of atherosclerosis without infiltrating the artery wall. However, what may be likely is that the indigenous bacteria play a role in the mechanism of graft infections after the surgical bypass procedures and are aggravating necrotic processes in ischemic tissues.
Moreover, microbes transported in lymphatics running in close proximity to veins may also colonize the venous walls and subsequently initiate the intra-luminal coagulation process [25]. The high prevalence of isolates in the varicose walls as well as pictures of bacteria migrating from the specimens on Hemoline plates are evidence for pre-existence of bacteria in these tissues. Whether they play a role in the pathomechanism of formation of varices remains unknown [19]. They may become activated after venectomy for coronary artery bypass surgery [20–22]. There is no pertinent literature on lower limb vein bacteriology.
Each case of lymphedema is predisposed to infections and chronic DLA [32]. This is because of impairment of bacterial elimination via lymphatics. Lymphedema is complicated by infection of skin and deep tissues in approximately 40% of cases, irrespective of what is the primary etiologic factor for development of this condition [33]. We detected bacterial isolates in 33%–60% in lymphatics and lymph. The recurrence rate of acute attacks of DLA is greater in cases with long duration of edema. It is followed by rapid increase in limb volume. In lower extremities, infection with inflammation affects approximately 50% of patients [23]. It is most common in the post-inflammatory type of lymphedema, followed by the post-traumatic and post-surgical types. In advanced stages of lower limb lymphedema systemic infectious episodes requiring hospitalization and intensive antibiotic therapy are common, especially in tropical countries [23]. Patients with acute episodes of DLA reveal bacteremia in a high percentage of cases [34]. Diversity of blood and tissue bacterial isolates in these patients points to a breakdown of the skin immune barrier in lymphedema and subsequently indiscriminate bacterial colonization of deep tissues and spread to blood circulation. Fatal cases were observed. Recurrency of DLA is caused by the permanent presence of bacteria in lymphedematous deep tissues. In addition to positive cultures, the evidence for it is the effectiveness of long-acting penicillin (benzathine penicillin) for decreasing the episodes of DLA from 100% to 9% during 1-y of prophylaxis [35].
The cryptic dormant bacteria identified by us most likely belonged to the persister cells [36]. Bacterial persister cells may represent a stage of dormancy that protects them from killing by antibiotics, even in concentrations exceeding the minimal inhibitory concentration. The low level metabolism of dormant persister cells prevents the penetration and blocking action of antibiotics. These cells can switch from the dormant to a planktonic stage. For most bacterial strains persister cells are found in biofilms, which contribute to recurrent infections after antibiotic therapy.
Taken together, our studies show that lower limb deep tissues contain indigenous dormant bacteria at low frequency that do not evoke, under physiologic conditions, a host reaction. Chronic and acute ischemia, venous stasis in the superficial and deep venous systems, and lymph flow stasis change the environment for the colonizing microbes resulting in their proliferation and evoking host inflammatory response. Our findings are the first in the bacteriologic literature showing cryptic dormant cells in tissues of human lower limbs. Most articles addressing problems of dormant persister cells contain observations from in vitro studies. We showed that they are present in live tissues; however, because of a low mass, long-term cultures are needed for their detection. Taking into account the presence of persister cells in live tissues and their resistance to antibiotics, peri-operative antibiotic prophylaxis becomes mandatory and the dosages should be increased from those presently applied. The type of antibiotic is not decisive as the identified bacterial strains were susceptible to most antimicrobials.
Footnotes
Acknowledgments
This study was supported by a grant from the National Center of Science (Poland) Nr N N404 1644 34.
Author Disclosure Statement
No competing financial interests exist.