
Abstract
Abstract
Background:
The etiology of sepsis is broad. The peritoneal cavity displays compartmentalization with respect to inflammatory responses, so peripheral blood responses to sepsis of abdominal vs. non-abdominal origin are expected to be divergent. Lymphocytes and invariant natural killer T (iNKT) cells play important roles in survival from sepsis, as they dampen the neutrophil and macrophage responses. We assessed whether circulating iNKT cells display distinct phenotypic profiles depending on the presence of abdominal vs. non-abdominal infection with sepsis.
Methods:
Patients with sepsis, defined as infection confirmed microbiologically with a systemic inflammatory response syndrome (SIRS), were enrolled prospectively. They were categorized as having either exclusively sepsis of abdominal or exclusively non-abdominal origin. The white blood cell (WBC) count was recorded. Whole-blood staining with monoclonal antibodies to CD3, V-alpha-24 (to identify iNKT cells), and CD69 (marker of early activation) was applied.
Results:
Of the 53 enrolled patients, 18 had abdominal infection. Pneumonia was the most common non-abdominal type. There was no difference in gender, age, Acute Physiology and Chronic Health Evaluation (APACHE) II score, WBC count, or CD3+ T cells (7.1%±1.6% vs. 6.5%±0.9%; p=0.75) in the two groups. Patients with abdominal infection had a higher proportion of iNKT cells (2.7%±1.1% vs. 0.89%±0.14%; p=0.032). Correcting for WBC count, this translated into a higher absolute number of iNKT cells (3.4±1.8×107/L vs. 0.74±0.15×107/L; p=0.03). Patients with sepsis of abdominal origin had a lower percentage of CD69+ iNKT cells (9.1%±3.1% vs. 27.2%±5.8%; p=0.028). In patients in shock vs. those who were not, patients with non-abdominal infection exhibited a greater number of iNKT cells (1.47±0.3 v. 0.62±0.1×107/L; p=0.022) and percentage of activated iNKT cells (53±14.5% vs. 17.9±4.8%; p=0.04). Patients with non-abdominal infection who died had a lower absolute number of activated iNKT cells (0.8±1.2×107/L vs. 0.34±0.1×107/L; p=0.023); however, no such shock or death correlation was noted in patients with sepsis of abdominal origin.
Conclusions:
Divergent sepsis etiologies display distinct blood iNKT cell population changes. In non-abdominal infection, this difference was associated with septic shock and death. Elucidating the importance and basis for these changes relative to the response to sources of infection will help clarify appropriate diagnosis and management of the patient with sepsis.
N
Much of the longer-term effects of sepsis is driven by immune and inflammatory responses. Antimicrobial agents may target the organism, and resuscitation chases the hemodynamic consequences of an infection, but there remains no specific treatment aimed at the inflammatory response to the infection. Our understanding of the inflammation associated with sepsis has evolved from consideration of single cytokines to recognition of the larger role that certain cell populations play in the coordination, or conversely, the dis-coordination, of this response.
Lymphocytes play a vital role in survival from critical illness in both mice and human beings [3–5]. Recently, attention has been focused on the role of innate regulatory T cells in the coordination of the immune and inflammatory response to sepsis, specifically invariant natural killer T (iNKT) cells. Lymphocyte loss and dysfunction follow the onset of critical illnesses, including sepsis [6]. We have shown previously that despite a loss of both CD3+ lymphocytes and γδ-T cells (another innate regulatory T cell), iNKT cells were preserved after the onset of sepsis [7]. Furthermore, patients who died had significantly fewer circulating iNKT cells than did survivors [7].
Invariant natural killer T cells are a pluripotent subset of lymphocytes with a broad spectrum of activity [8,9]. Such cells play key roles in combatting many clinically relevant infections such as those caused by Pseudomonas [10] and Streptococcus [11]. Investigators, including ourselves, have shown that iNKT cells are capable of modulating neutrophil [12] and macrophage activity [13]. The liver is the largest source of iNKT cells in both human beings and mice.
The abdominal cavity behaves in a compartmentalized fashion with respect to its response to infectious and inflammatory conditions [14]. A rapid and diverse response to inflammation is noted in the modulation of the peritoneal-cell populations in response to injury and inflammation [15]. The abdominal cavity tends to sequester cytokines, chemokines, and cell populations to a greater extent than is observed with other sources of inflammation and infection, leading to a greater discordance between the hematologic and abdominal findings. In a murine model, an exodus of iNKT cells from the liver and circulation into the peritoneal cavity occurs in response to abdominal sepsis [13].
It is crucial that our understanding expands regarding how distinct regulators of the immune and inflammatory system respond to distinct anatomical sources of sepsis. Understanding the differential regulators of the immune and inflammatory responses to distinct anatomical sources of infection will be key to better individualizing and directing of future therapies. We hypothesized that patients with infection in the abdominal cavity and sepsis will display a different pattern of circulating iNKT cells than patients with non-abdominal sources of sepsis.
Patients and Methods
Patients
Approval was obtained for this study from the Institutional Review Board of Rhode Island Hospital. Critically ill surgical and trauma patients with sepsis who were in the intensive care unit (ICU) were enrolled prospectively. To be included, patients had to have a microbiologically or clinically proved source of infection and two or more systemic inflammatory response syndrome (SIRS) signs. Exclusion criteria were death within 48 h of presentation, a history of lymphoma or leukemia, and multiple sources of sepsis at the time of enrollment. At the time of the morning blood draws in the ICU for determination of the white blood cell (WBC) count, blood was collected for this study also. All blood sampling occurred within 36 h of presentation to the ICU with a diagnosis of sepsis.
Charts were reviewed to determine the source of sepsis, and patients were divided into abdominal or non-abdominal types. Patients with abdominal infection were those with a clinically or microbiologically confirmed intra-abdominal source. Pneumonia was diagnosed using fiberoptic bronchoscopically guided bronchoalveolar lavage (BAL), with quantitative cultures showing ≥105 colony-forming units/mL. Clinical data extracted from the chart included age, gender, site of infection, need for vasopressor agents, and death. Patients were defined as being in shock if they were receiving vasoactive medications (e.g., norepinephrine, phenylephrine, or vasopressin) at the time of the study blood draw. Clinical and laboratory data were extracted to calculate the Acute Physiology and Chronic Health Evaluation (APACHE) II score at the time of the blood draw.
The techniques used in this work have been described in previous publications from our laboratory [7]. In brief, whole blood analysis typically was undertaken with 100 mcL of fresh blood. Monoclonal antibodies, used according to their manufacturer's recommendations, were anti-CD3 (T cells), anti-CD69 (a marker of early cell activation), and anti-TCR Vα24-PE (for the detection of iNKT cells). Red blood cell (RBC) lysis was performed using a human RBC lysis kit (ebiosciences, San Diego, CA). The sample was then centrifuged, and the pellet was re-suspended and analyzed using a Becton-Dickinson fluorescence-activated cell sorting array (Franklin Lakes, NJ). Both the WBC count and the absolute lymphocyte number were available from the daily laboratory tests ordered by the treatment team. These data allowed the calculation of the absolute number of CD3+ cells, iNKT cells, and activated cells.
Results are expressed as mean±standard error of the mean. Chi-square values were used for categorical data and the Mann-Whitney U test was used for non-parametric data were used. Statistical analysis was done using SigmaPlot version 11 (Systat Software, Inc., Chicago, IL). Significance was set at p<0.05.
Results
Over the period of the study, 53 patients were enrolled, 18 of whom had an abdominal source of sepsis. Of those with non-abdominal sources of sepsis, pneumonia was the most common cause. Comparing patients with abdominal and non-abdominal sources of sepsis, there were no significant differences in age, gender, or APACHE II score (Table 1). There also was no difference in the mortality rate or the presence of shock in patients with abdominal vs. non-abdominal infection. Furthermore, there was no significant difference in leukocytosis or CD3+ lymphocytes as a percent of total leukocytes in patients with abdominal vs. non-abdominal infection.
APACHE=Acute Physiology and Chronic Health Evaluation; SD=standard deviation.
Next, we assessed the circulating iNKT-cells in both groups. Patients with abdominal infection had significantly higher numbers of iNKT cells as a percentage of CD3+ lymphocytes (2.66%±1.1% vs. 0.89%±0.14%; p=0.032) (Fig. 1A). Adjusting for absolute WBC count, this translated into abdominal infection patients having significantly higher absolute numbers of circulating iNKT cells (3.4±1.8×107/L vs. 0.74±0.15×107/L; p=0.03)(Fig. 1B). Patients with abdominal infection had significantly lower numbers of CD69+(activated) iNKT cells (9.1%±3.1% vs. 27.2%±5.85; p=0.028) (Fig. 2).

Circulating iNKT cells in sepsis of abdominal vs. non-abdominal origin. Patients with the former had higher numbers of circulating iNKT cells as a percentage of CD3+ lymphocytes. Accounting for individual white blood cell counts led to elevated absolute numbers of circulating iNKT cells in patients with abdominal infection.

Extent of CD69* expression (activation) on circulating iNKT cells. Patients with abdominal sources of sepsis had significantly higher degrees of activation than patients with non-abdominal sources.
To the extent that there was an association between iNKT-cell numbers and the presence or absence of septic shock, patients were stratified by whether they manifested shock at the time of blood sampling. First, we assessed the association in patients with non-abdominal sources of sepsis. Herein, it was noted that patients with septic shock had higher numbers of iNKT cells (1.47±0.3×107/L vs. 0.62±0.1×107/L; p=0.022). Further, a greater percentage of these iNKT cells were activated (C69+)(53%±14.5% vs. 17.9%±4.8%; p=0.03) (Fig. 3). However, in patients with abdominal sources of sepsis, no association was noted between shock and either number of iNKT cells (4.1±3.7×107/L vs. 2.6±1.9×107/L; p=0.7) or number of activated iNKT cells (8%±3.7% vs. 12.7±5.3%; p=0.6) (Fig. 4).
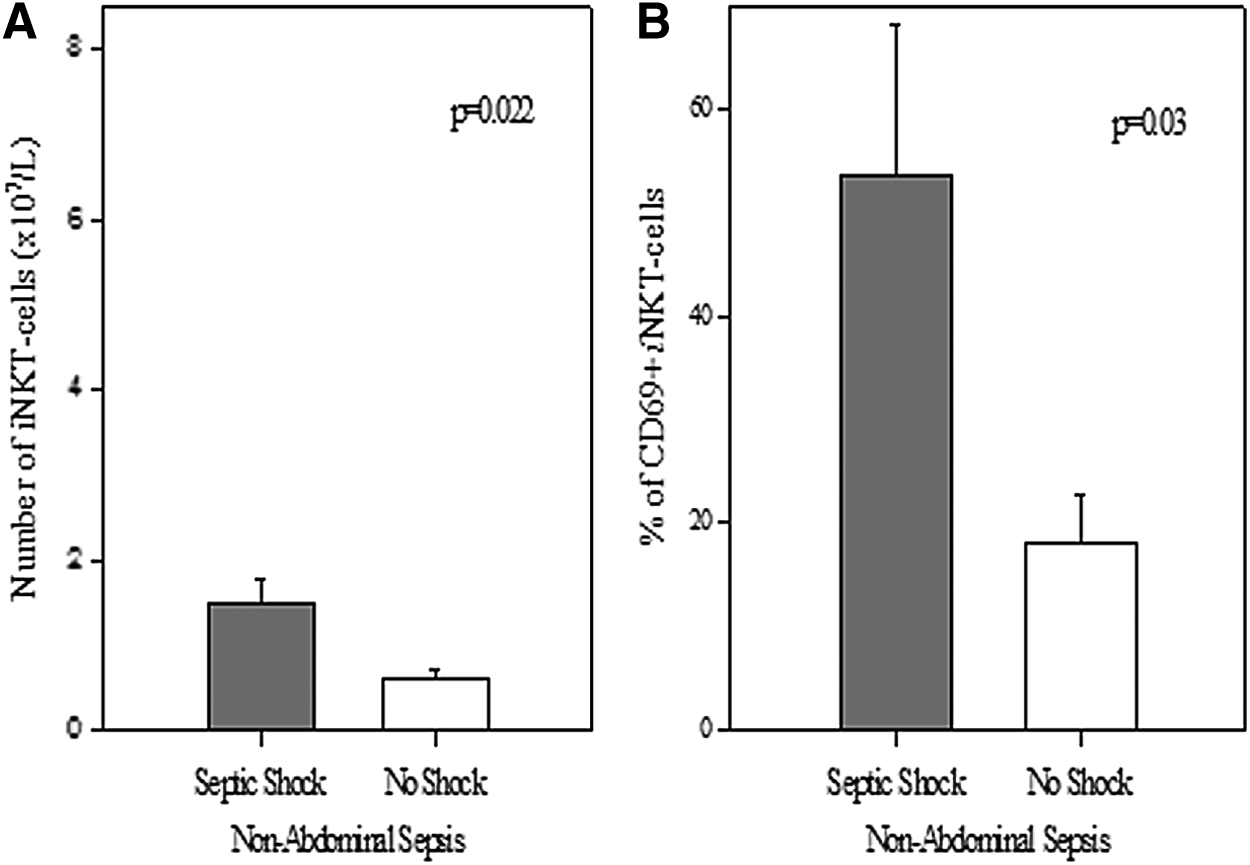
In patients with non-abdominal sources of sepsis, those with shock (
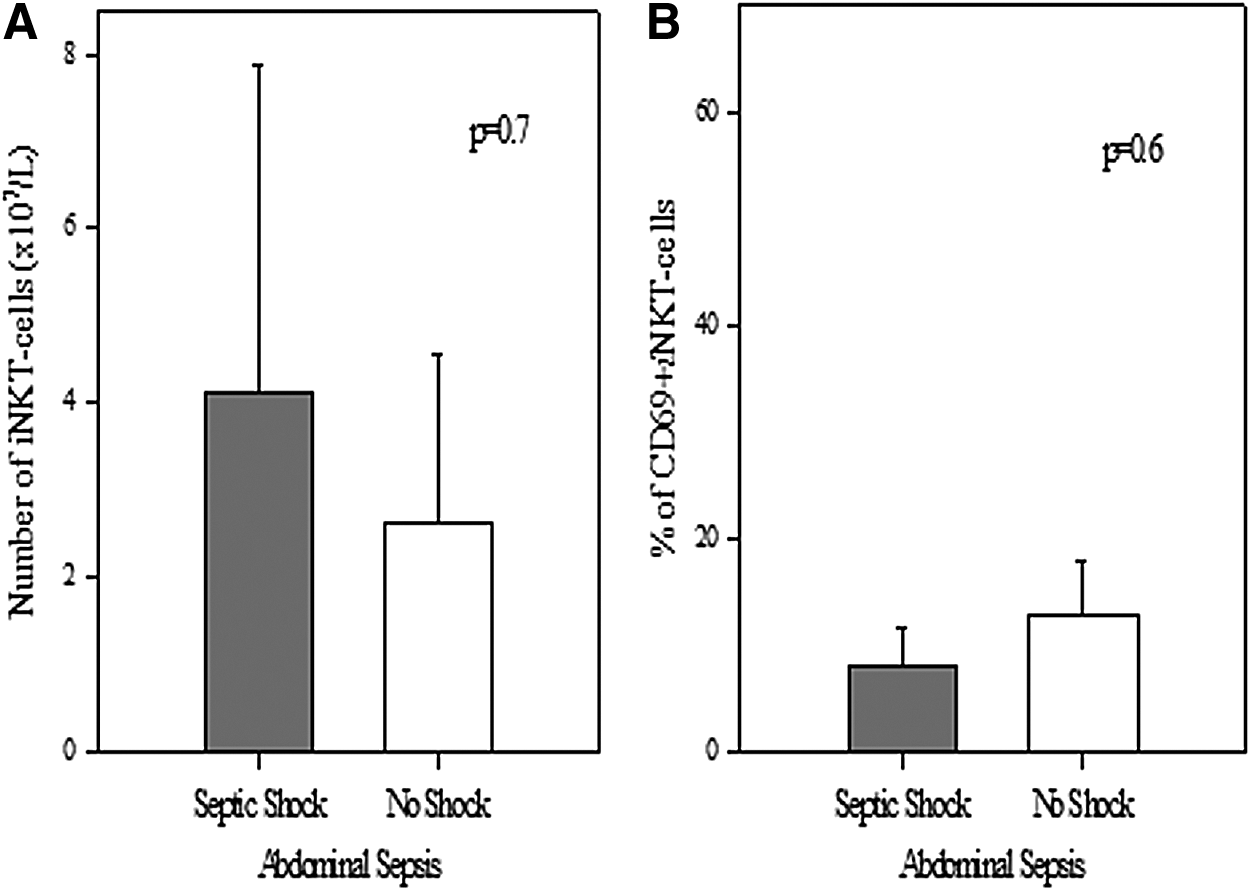
In patients with abdominal infection, no association was noted between numbers of circulating iNKT cells (
Next, we assessed for an association between death and alterations in iNKT cells. In patients with non-abdominal sources of sepsis, those who lived had significantly greater numbers of iNKT cells than those who died (0.8±1.2×107/L vs. 0.34±0.1×107/L; p=0.023). Further, patients with non-abdominal infection who lived had significantly greater percentages of iNKT cells that were activated (32.9%±6.9% vs. 7.9%±3.6%; p=0.032). Again, no association was noted in patients with abdominal infection who lived vs. those who died with respect to absolute numbers (2.8±2×107/L vs. 3.5±3×107/L; p=0.85) or percentage of activated (12.1%±4.9% vs. 14.9%±11%; p=0.79) iNKT cells.
Discussion
A full understanding of the pathophysiology of sepsis remains elusive. Sepsis is the systemic response to an infectious source. This definition remains broad and vague and is rarely individualized to the type or origin of the septic event. Clinical trials recognize that factors such as age, gender, infecting organism, source of sepsis, and need for or response to fluid resuscitation affect the outcomes of patients with sepsis [16]. This assumes that the inflammatory responses remain uniform across these demographic or microbiologic strata. There is limited recognition of how the specific patterns of the cellular mediators of sepsis affect outcomes. Indeed, the individualized cellular inflammatory response has received little attention.
Current clinical practice often tailors and directs anti-microbial agents to specific organisms or fluids to hemodynamic responses. However, there is no agent that targets differences in the specific/localized immune and inflammatory systems. Hence, we undertook an analysis of the specific response of a key innate regulatory T cell, the iNKT cell, as it related to differing sources of sepsis. We stratified our patient population into abdominal versus non-abdominal sources of sepsis. We show herein a dramatic difference in the iNKT-cell response to these sources of sepsis. Specifically, patients with abdominal sources of sepsis had significantly higher numbers of iNKT cells, both as a percentage of CD3+ lymphocytes as well as in absolute numbers of circulating cells, than did patients with non-abdominal sources of sepsis.
The liver is the largest source of iNKT cells [8]. Organisms within the abdominal cavity reach the liver rapidly via lymphatic drainage and the portal venous drainage of the intestines, exposing and thereby activating liver iNKT cells quickly. This anatomic pathway may in part explain the differences observed in our data. In a murine model, direct liver injection of bacteria led to activation and migration of iNKT cells within the liver. In our cecal ligation and puncture (CLP) model, we demonstrated that, after the onset of abdominal sepsis, liver iNKT cells undergo an exodus from the liver, migrating into the blood stream and the abdominal cavity [13]. Furthermore, the ability of iNKT cells to migrate appears to be controlled via programmed death receptor-1 (PD-1); we have shown that iNKT cells from PD-1 knockout mice were incapable of this migration to the abdominal cavity in response to abdominal infection.

Patients with non-abdominal sources of sepsis who died had significantly fewer circulating iNKT cells (
In contrast, both the lung and the blood stream carry considerably fewer numbers of iNKT cells at baseline compared with the liver [8]. However, the lung is a site wherein iNKT cells exert a profound inflammatory response in both infectious and non-infectious/allergic conditions [17]. It is believed that iNKT cells involved in lung infections are either from the lung itself or are derived from already-circulating iNKT cells rather than coming from the liver population. These observations would be in keeping with our clinical finding here of a smaller number of circulating iNKT cells in patients with non-abdominal sepsis.
Interestingly, we noted significantly different extents of activation of iNKT cells depending on the source of the sepsis. Thus, in patients with abdominal sources of sepsis, a significantly lower percentage of iNKT cells are activated than in those with non-abdominal sources of sepsis. This discrepancy may explain some of the apparently conflicting data pertaining to the role and function of iNKT cells. With respect to lung pathology, iNKT cells play a key role coordinating the leukocyte to lung interaction as it pertains specifically to preventing the influx of innate inflammatory cells, most notably neutrophils [12]. In models of experimental asthma or pneumonia, activated iNKT cells again appear to play an important gate-keeper role in modulating the influx of neutrophils into the lung [12]. This would appear to emphasize the need for greater numbers of the circulating iNKT cells to be activated, thereby interacting with the circulating innate cells, including neutrophils, in cases of sepsis of non-abdominal origin.

No association was noted in patients with abdominal sources of sepsis between the numbers of circulating iNKT cells (
The iNKT cells appear to play a more local-regional role in abdominal-origin sepsis. We have demonstrated previously that, after CLP, it was the activated abdominal iNKT cells that played a key role in modulating the abdominal macrophages in response to sepsis [13]. We believe that our data support the contention that after activation and exodus from the liver, these activated iNKT cells are needed within the peritoneal cavity rather than in the circulation. Furthermore, iNKT cell blockade leads to reduced expression of macrophage co-stimulatory molecules CD40 and CD80 accompanied by decreased maturation of splenic macrophages [18].
Furthermore, the abdominal cavity displays a unique and independent cytokine and chemokine response to a variety of insults, a response distinct from that seen in the rest of the body [14]. This study was followed by a further observation from the same investigators that abdominal cavity inflammation was associated with an increase in the production of stromal cell-derived factor 1 alpha (SDF-1α), a chemokine important for the inward migration of CXCR-4-positive cells [15]. In that study, the omentum was an abundant source of SDF-1α. It is noted that CXCR-4 is expressed by a variety of cells, including lymphocytes. Activation of the SDF-1α/CXCR4 axis results in perivascular recruitment of lymphocytes from the circulation. Thus, the authors proposed that the omentum played a key role in the recruitment of cells into the peritoneal cavity [15]. These findings were in keeping with prior observations that the licensing of liver iNKT cells for both potentiated random and directional migration in a patrolling fashion in response to infection is regulated at a local tissue level.
After sepsis, morbidity and death often are driven by the resultant end-organ damage, a major contributor to which is acute lung injury [16]. Although we did not observe any association between iNKT cell numbers of activation and death in patients with abdominal-origin sepsis, we did notice a distinct association in sepsis of non-abdominal origin between increased shock and death and increased numbers of circulating and activated iNKT cells. The higher numbers of circulating and activated iNKT cells may be correlated with the severity of illness and therefore the presence of shock. In pulmonary viral infections, iNKT cells dampen the inflammatory response within the lung. Further, iNKT cells affect pulmonary infiltrating inflammatory monocytes directly, thereby limiting tissue destruction [19]. Thus, it is proposed that in the absence of iNKT cells, increased pulmonary inflammation would be seen, potentially leading to a greater risk of death from acute lung injury [20]. This would be in keeping with our data showing lower iNKT cells numbers in patients who died. This view is supported by evidence from the recent H1N1 pandemic, wherein a higher mortality rate was noted in patients who exhibited decreased numbers of circulating iNKT cells [19].
To our knowledge, this is the first time that a particular inflammatory response, specifically innate regulatory T-cell changes in the frequency, number, and activation status, has been assessed with respect to abdominal vs. non-abdominal sources of sepsis. Our data suggest that the future of therapy for sepsis will involve targeting these individualized iNKT cell responses.
Footnotes
Acknowledgments
Portions of this work were supported by grants from the Armand D. Versaci Research Scholar in Surgical Sciences Award (to J.S.Y. and S.F.M.), the Shock Society's Junior Faculty Fellowship Award (D.S.H.), and R01-GM046354 (A.A.).