
Abstract
Abstract
Background:
This study aimed to determine the effects of probiotic Lactobacillus plantarum on colitis in mice with multiple drug-resistant bacteria, and to judge the intervention function of probiotics in the resistance of pathogenic bacteria.
Methods:
Male BALB/C mice were divided into four groups. The colonic tissues were collected for pathology observation, permeability, transepithelial electrical resistance (TER), and tight junction protein expression detection at death, and the feces were collected for detecting drug susceptibility of MDR-PA.
Results:
MDR-PA mice developed severe intestinal inflammation and tissue damage in which paracellular permeability was increased, in conjunction with decreased expression and redistribution. LP administration attenuated colitis of MDR-PA mice. The drug susceptibility diameter of MDR-PA has increased.
Conclusion:
Probiotics can treat diarrhea of mice by MDR-PA and protect the intestinal barrier function. Probiotics can weaken the resistance of multiple drug resistance in Pseudomonas aeruginosa to some extent.
I
This study aims to determine the effects of probiotic Lactobacillus plantarum LP-Only (LP) on colitis in mice, a model of intestinal infection with multiple drug-resistant bacteria, and to judge the intervention function of probiotics in the resistance of pathogenic bacteria.
Patients and Methods
Bacterial strain and cultivation
The probiotic bacterial strain used in the study is LP, which was originally isolated from the feces of healthy infants on the basis of having desirable probiotic properties. Strain LP-Onlly was identified as L. plantarum based on the pattern of carbohydrate fermentation and 16S rDNA (GenBank accession NO. AY590777) and was deposited at China General Microbiological Culture Collection Center with accession number CGMCC1258. Lactobacillus plantarum was expanded from a lyophilized stock (including 10% skim milk, 1.5% fucose, 0.5% glycerol, 2% sorbitol, 1% malt dextrin, (wt/vol)) in deMan-Rogosa-Sharpe (MRS) medium (MRS, Oxoid, UK) at 37°C for 18 h under anaerobic conditions created by anaerobic bags (Anaerocult A, Merck, Damstadt, Germany) in anaerobic jars (BBL Becton Dickson, Cockeysville, MD, USA). Bacteria were harvested by centrifugation at 4000r/min for 10 min, and washed twice with sterile phosphate-buffered saline (PBS, pH 7.5). The pellet of viable cells resuspended in PBS was adjusted spectrophotometrically to contain 109cfu/mL and stored at 4°C for use.
Multiple drug-resistant Pseudomonas aeruginosa (MDR-PA) was isolated from clinical strains of Shanghai Sixth People's Hospital microorganism inspection center. It was all resistant to penicillin, cephalosporin, hydrocarbon enzymes, and beta lactamase inhibitor. It was identified by drug-sensitive test automatic microbial analysis system.
Animals
Male BALB/C mice, age 6 wks (S&p Bi kai laboratory animal co., LTD, Shanghai, China) were used. The mice were housed under specific pathogen-free conditions at a constant temperature and in a 12:12 h light:dark cycle in Shanghai Jiao Tong University Medical School, and were given free access to water and consumed a commercial sterile diet (SLACOM Inc., Shanghai, China). The protocol was approved by the Animal Care and the Ethics Committee (Shanghai Jiao Tong University, Shanghai, China).
Reagents
All chemicals were obtained from Sigma (St Louis, MO, USA), unless otherwise indicated.
Experimental design
Mice exhibiting no overall initial differences in body weight were equally randomized into four groups: Normal control (NC mice without LP treatment and MDR-PA infection), MDR-PA (MDR-PA mice with MDR-PA infection, without LP treatment), MDR-PA+LP (MDR-PA+LP mice with MDR-PA infection and LP treatment), and LP group (LP mice with LP treatment, without MDR-PA infection) (n=10). Mice in the MDR-PA groups were administered free drinking water with ceftriaxone sodium as a dose of 8 g/kg/d per mice for 4 d. Water was changed once a day. Mice were maintained by gavage with MDR-PA as a dose of 108cells/d per mice for 1 wk. Mice in the probiotic groups were administered by gavage with a dose of 109cells/d per mice for 3 wks, whereas the other two groups consumed PBS only. Mice were maintained for another 1 wk without LP treatment. Mice were sacrificed by cervical dislocation at the end of experiment. The colonic tissues were collected for pathology observation and inflammation score, permeability, transepithelial electrical resistance (TER), and tight junction protein expression detection at death after 3 wks treatment of LP, and the feces were collected at 1 wk intervals throughout the experiment for the analysis of drug susceptibility of MDR-PA.
Medicine sensitive experiment
Fecal samples for MDR-PA analysis were aseptically collected at an interval of 1 wk, and were dispersed in 2.5mL Ringer's buffer supplemented with 0.05% (wt/vol) cysteine hydrochloride, then serially diluted tenfold. 100 mL of the appropriate dilutions was spread onto Blood agar medium, and incubated anaerobically for 24 h at 37°C. The strain was observed for cell morphology and Gram staining was carried out. Biochemical characteristics of the strain were determined using Microscan Autoscan-4, according to the manufacturer's instructions. The drug sensitivity tests of bacteria were tested by K-B paper diffusion method, which vaccination on M-H AGAR plate with French merry Emmanuel VITEK2 AST - GN09 susceptibility card.
Histopathology
At death, the colon tissues were fixed in 10% neutral phosphate buffered formalin, routinely processed, sectioned at 6 mm, and stained with hematoxylin and eosin for light microscopic examination. Each segment of the colon (cecum, ascending, transverse, and descending colon) was assigned a histological score ranging from zero to four based on the criteria adapted from Berg [5].
Enzyme-linked immunosorbent assay
The colonic concentrations of the cytokines TNF-α and IFN-γ were determined by using ELISA kits (BD Pharmingen, Oxford, UK). Briefly, the colon tissues were thawed, cut, and homogenized by using an Ultra-Turrax homogenizer (IRK-WERKE, Germany) in 800mcl of chilled homogenization buffer containing phosphate-buffered saline (PBS), 2% (vol/vol) fetal calf serum, and 0.5% (wt/vol) cetyltrimethylammonium bromide. The homogenate was microcentrifuged at 17,940 g for 15 min at 4°C. The protein content of the supernatant was determined by a BCA protein estimation kit (Pierce, Rockford, IL). TNF-α and IFN-γ concentrations in colonic supernatants were measured according to the manufacturer's instruction and are expressed as picograms per milligram tissue.
Immunofluorescence
The localization of ZO-1, occludin, and claudin-1 proteins was determined by immunofluorescence confocal microscopy. The colon segments were immediately removed, washed with PBS, mounted in the embedding medium (Tissue-Tek, Sakura), and stored at −80°C until use. Frozen sections were cut at 10mcm, and mounted on the slides. The slides were incubated in pre-warmed (95°C) 10mM sodium citrate buffer for 5 min and then cooled to room temperature. The nonspecific background was blocked by incubation with 5% bovine serum albumin plus 5% newborn bovine serum in PBS for 30 min at room temperature. The sections were incubated with goat polyclonal antibody against ZO-1, rabbit polyclonal antibodies against occludin, and mouse monoclonal antibody against claudin-1 at 4°C overnight and afterward with phalloidin-fluorescein isothiocyanate at room temperature for 1 h. The sections were probed with respective Cy3-conjugated secondary IgG antibodies (Jackson ImmunoResearch Laboratories). The nuclei were counterstained with 4′-6-diamidino-2-phenylindole. Next, the slides were washed and mounted in Vectashield mounting medium (Vector Laboratories, Burlingame, CA). Slides incubated without any primary antibody were used as negative controls. Confocal analysis was performed with an LSM510 laser scanning confocal microscope (Zeiss, Thornwood, NY).
Assessment for the barrier function of tight junctions
The barrier function of tight junctions was determined by measuring TER and paracellular permeability of nonionic mannitol, according to the method adapted from Madsen [6]. For the evaluation of TER, the freshly isolated and striped colonic mucosae were mounted in a modified Ussing chamber (EasyMount Chamber; Physiologic Instruments, San Diego, CA) with an exposed area of 0.3cm2. The tissues were bathed on both sides with 3 mL of oxygenated Krebs buffer, which was maintained at 37°C by a heated water jacket and gassed with 95% O2-5% CO2. The tissues were voltage clamped at 0mV to monitor short-circuit current (Isc) by using a Dual Voltage Clamp amplifier (VCC MC2; Physiologic Instruments) connected via a PowerLab8SP (AD Instruments Pey, Castle Hill, Australia) to a computer. The current deflection (ΔIsc) was caused by applying a 1-mV pulse for 0.5 s at 60 s intervals under the short-circuit condition. The TER was calculated according to Ohm's Law (i.e., TER=PD/ΔIsc), where PD is expressed as millivolts (mV), Isc as microamperes per square centimeter (mcA/cm2), and TER as ohms times centimeters squared (Ω·cm2). For the evaluation of paracellular permeability, 3% (wt/vol) mannitol was added to the mucosal side of the isolated colon after a 30 min equilibration period. Then 200 mcl of fluid was collected from the serosal side every 30 min for 120 min. The mannitol concentration was determined by high-performance liquid chromatography (HPLC) (Waters, Milford, MA) with a SK-801 shodex(8.0×300mm2, 5 mcm; Showa Denko, Tokyo, Japan) attached to a refractive index detector (Waters). The mobile phase was HPLC grade water, the flow rate was 1mL/min, and the column temperature was maintained at 80 °C.
Statistical analysis
All statistical analyses were performed by use of GraphPad Prism 5.0. Numerical data with a normal distribution were expressed as means±SD, and multiple groups were compared by one-way analysis of variance followed by least significant difference post hoc test. Comparisons between two groups were done by Student t-tests. Spearman rank correlation coefficients were computed for correlation analyses. The χ2 test was used for the comparison of categorical data among the groups. A p value of less than 0.05 was considered statistically significant.
Results
Effects of LP on histopathological manifestation of colitis in MDR-PA mice
Histological analysis showed that colitis developed in all MDR-PA mice, in which mucosal ulceration, erosion, and a large number of lymphocyte and neutrophil infiltration in the lamina propria were observed (Fig. 1). The inflammatory score was substantially greater in MDR-PA mice than in NC mice (p<0.01; Fig. 2). However, after 3 wks treatment of LP, only mild inflammatory cell infiltration was observed in MDR-PA mice, and the inflammatory score in MDR-PA mice was substantially reduced by LP treatment (p<0.01; Fig.2).

Effect of probiotic LP on colonic inflammation.
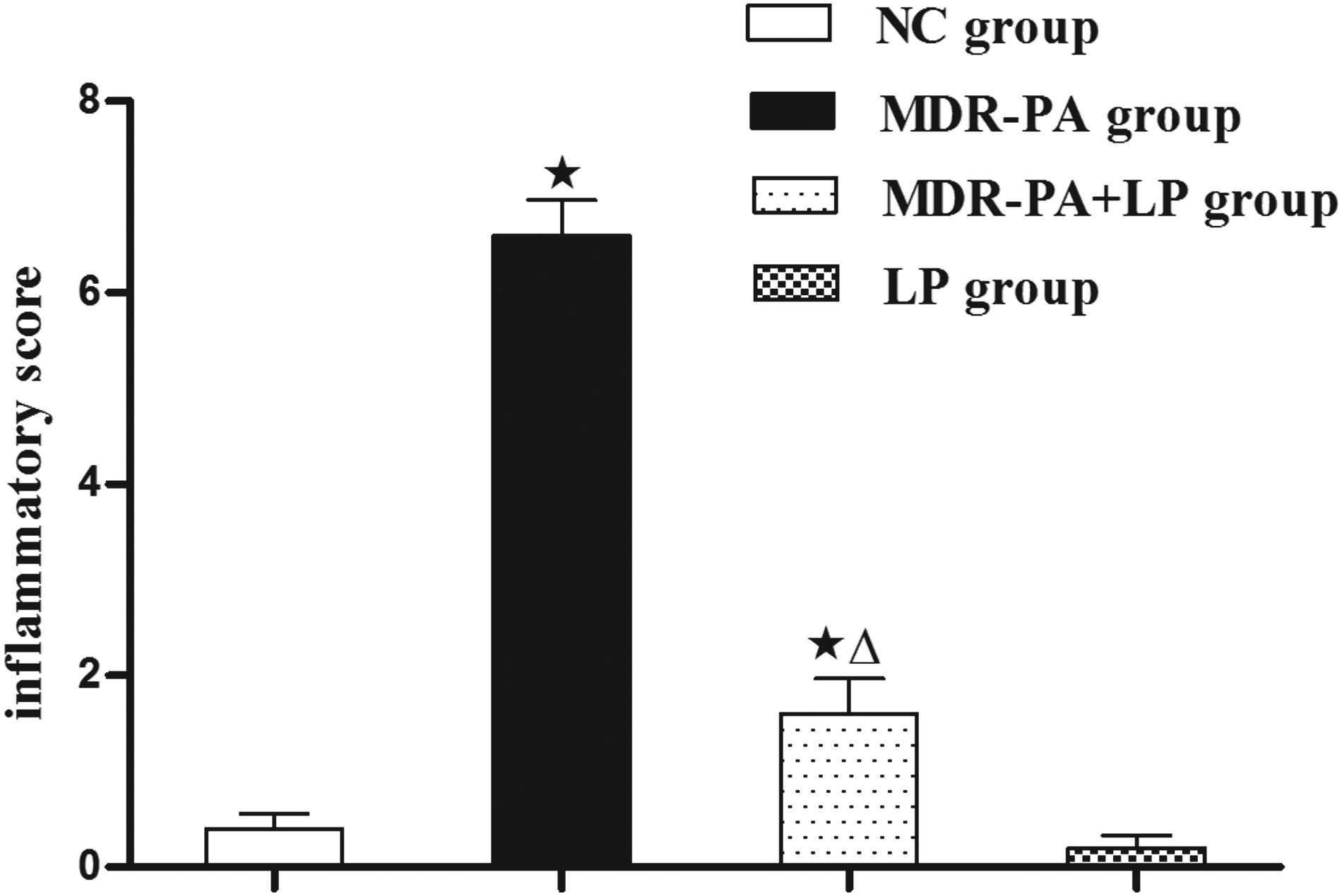
Lactobacillus plantarum (LP) attenuates colitis in MDR-PA group mice. Colonic inflammation scoring in NC group, mice treated with LP (LP group), MDR-PA mice (MDR-PA group), and MDR-PA mice treated with LP (MDR-PA+LP group). Error bars represent the mean±standard deviation (SD), n=10, ⋆p<0.01 compared with NC; ▵p<0.01 compared with MDR-PA.
The medicine sensitive experiment results of MDR-PA
The drug susceptibility results of multiple drug resistance of P. aeruginosa were almost no obvious changes after 2 wks treatment of LP. After 3 wks treatment of LP, the drug susceptibility diameter of MDR-PA increased compared with before, such as MEM10, TZP110, ATM-30 and CAZ-30. (Fig. 3).
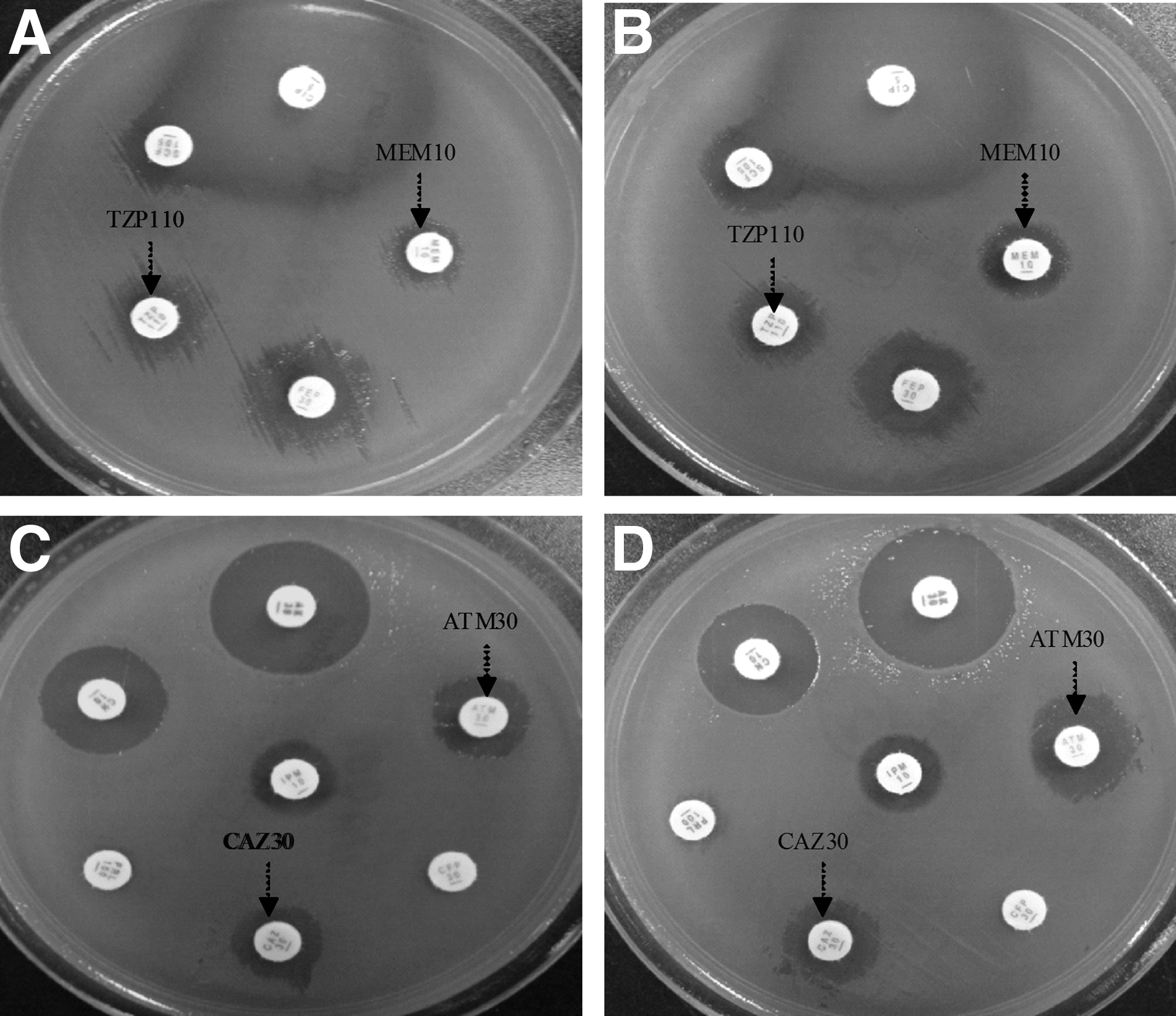
The results of drug susceptibility of MDR-PA. The drug susceptibility diameter of MEM10 and TZP110 in MDR-PA+LP mice
LP treatment reduces expression of proinflammatory cytokines in MDR-PA mice
The expression of TNF-α and IFN-γ in the colonic mucosa was substantially increased in MDR-PA mice, when compared with the concentrations in NC mice (334.45±69.74 vs. 135.18±33.46 pg/mg, p<0.001, and 533.25±112.21 vs. 208.08±37.07 pg/mg, p<0.001, respectively) (Fig. 4). However, the expression concentrations of TNF-α (157.58±45.85 pg/mg) and IFN-γ (259.08±89.29 pg/mg) in MDR-PA mice with the 3-wk LP treatment were substantially reduced, compared with the concentrations in MDR-PA mice (both p<0.001), although they were still greater than those in NC mice (p<0.05 and p<0.05, respectively). Although there was also a decrease in the expression of TNF-α (119.18±26.76 pg/mg) and IFN-γ (184.98±34.53 pg/mg) in LP-treated NC mice, the decrease did not reach the statistically significant concentration (Fig. 4).
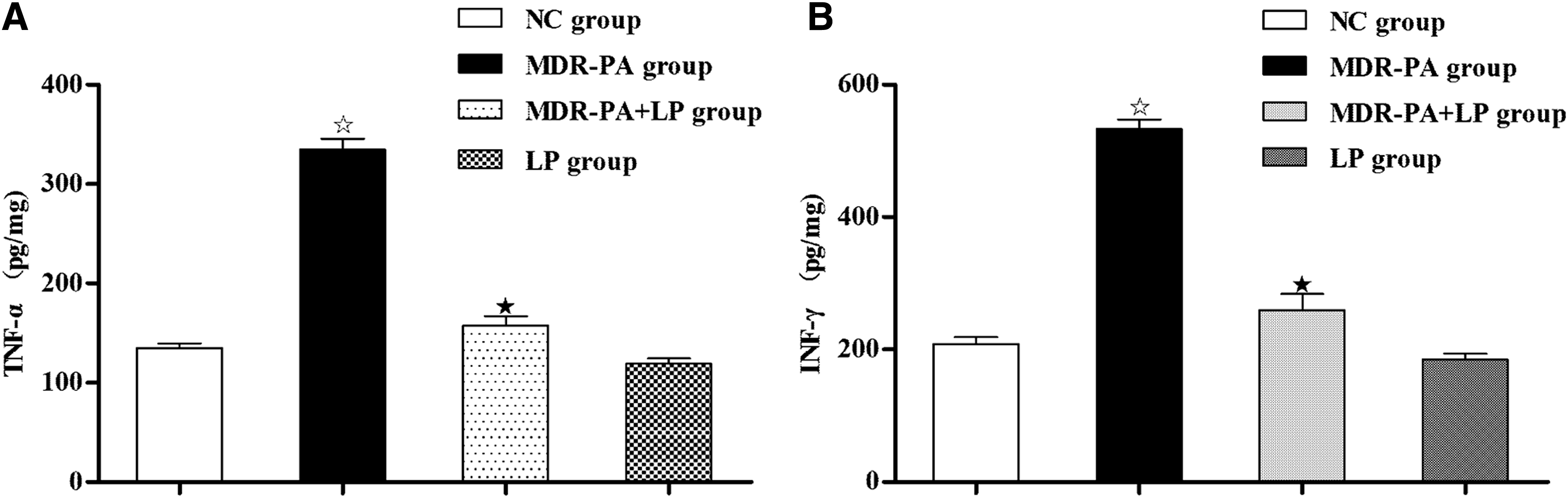
Effects of probiotic LP on production of proinflammatory cytokines, TNF-α
LP treatment reduces paracellular permeability
MDR-PA mice exhibited a substantial increase in the cumulative permeation of mannitol through the colonic mucosa, compared with NC mice (2.06±0.41 vs. 0.39±0.24%; p<0.001; Fig. 5a). In accordance with the increase in mannitol permeability, a substantial decrease of TER was observed in MDR-PA mice (39.89±10.40 Ω·cm2 vs. 74.08±4.61 Ω·cm2; p<0.001; Fig. 5b). However, these changes in MDR-PA mice were completely prevented by the 3-wk treatment with LP.
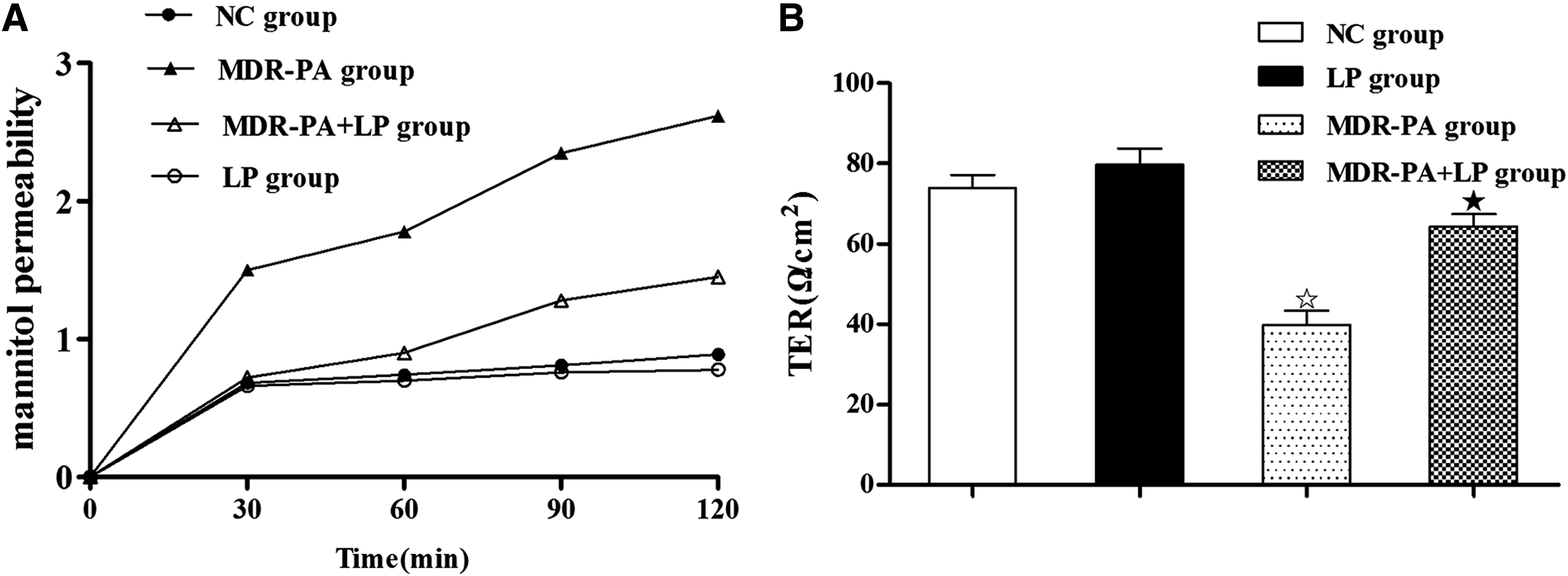
Effects of probiotic LP on colonic paracellular permeability.
LP treatment restores the expression and distribution of ZO-1, occludin, and claudin-1 proteins in MDR-PA mice
To identify whether there are changes in the apical junction protein expression and distribution, immunofluorescence assay of ZO-1, occludin, and claudin-1 were performed on the colonic tissue. Immunofluorescence assay demonstrated that the network of ZO-1, occludin, and claudin-1, which were predominantly localized along the apical cellular border, remained intact in NC mice. However, in MDR-PA mice, ZO-1, occludin, and claudin-1 staining in apical cellular border appeared to be decreased and discontinuous, with the redistribution of ZO-1, occludin, and claudin-1. All these changes of the proteins were prevented by LP treatment. (Fig. 6).
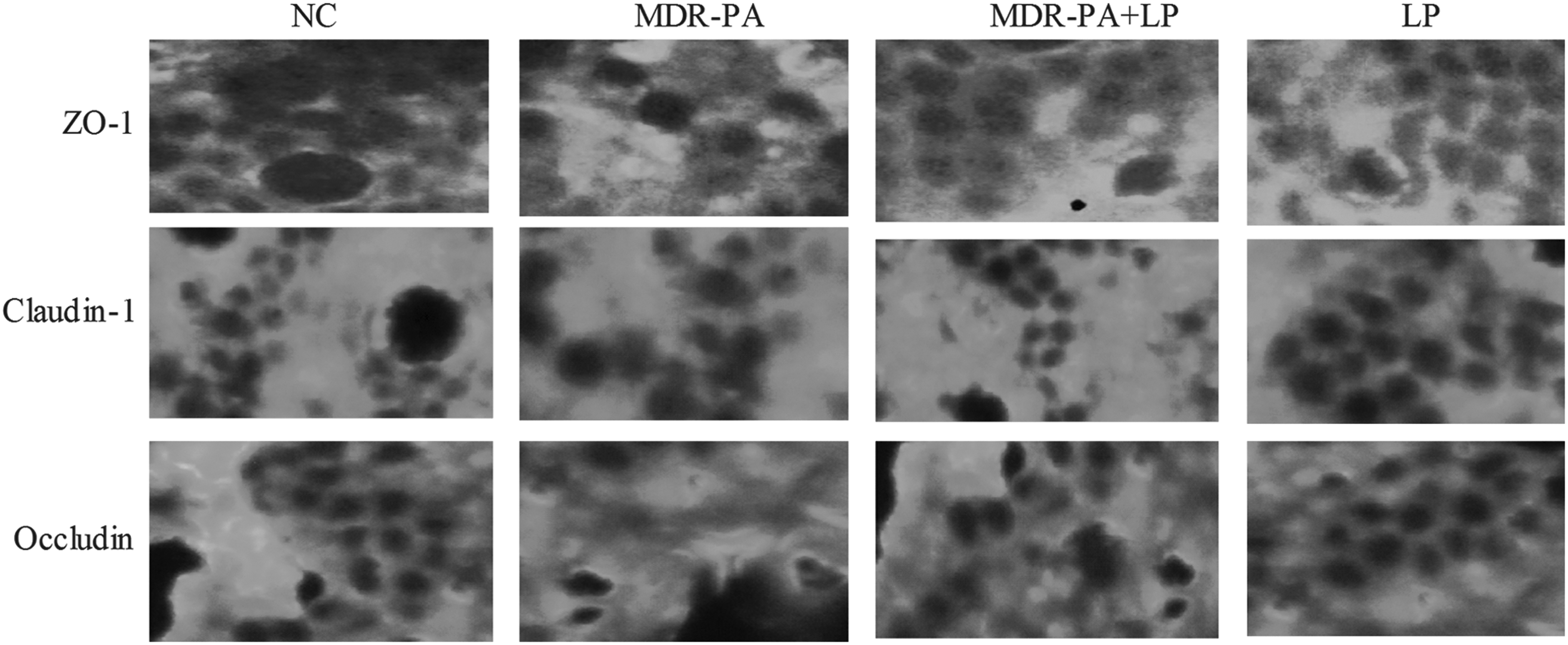
Effects of probiotic LP on the distribution and expression of intercellular junction proteins in the colon. Immunofluorescence for intercellular junction proteins including ZO-1, occluding and claudin-1 in the colonic epithelium of NC, LP, MDR-PA, and MDR-PA+LP mice (n=4 in each group).
Discussion
In recent years, more and more attention is being focused on clinical infection caused by P. aeruginosa. Pseudomonas aeruginosa is a known opportunistic pathogen. It is found in soil, water, skin flora, and most man-made environments throughout the world. It thrives not only in normal atmospheres, but also in hypoxic atmospheres, and has, thus, colonized many natural and artificial environments. It frequently causes septicemia, lower respiratory infection, indwelling catheter, intestinal infection after surgery and serious infections in burn patients [1,3,7–9]. Because of inherent characteristics, P. aeruginosa is to aminoglycosides, fluoroquinolones, cephalosporins, carbapenems, tetracyclines, monobactams, and sulfonamides, etc. Thus, there is no drug resistant sufficient solution strategy to solve the drug resistance of P. aeruginosa.
It has been demonstrated that probiotics can effectively reduce the bacteria toxicity, and increase the therapeutic effect of antimicrobial drugs. Lkhagvadorj et al. [10] found that the synbiotic treatment was effective for the treatment of intestinal infection caused by four MRSA strains with different toxin productions. Several studies performed either in murine models or in human beings have suggested that the administration of probiotics improves intestinal microbial balance and the modulation of immune functions and that probiotics may be interesting agents to eradicate or prevent VRE intestinal colonization [11,12]. Our experiment mainly studied the effect of LP on intestinal infection in multiple drug-resistant bacteria mice.
The integrity of the intestinal epithelial cells plays an important role in preventing the bacteria translocation. Epithelial cells in the small intestine are a type of brush border cell that are joined together by tight junctions; these were mainly constituted by ZO-1, 2, 3, occludin, claudins, cingulin, ezrin, and other additional components, to form a polymer impermeable membrane. These cells have a brush border surface to increase their absorptive surface area, thus making them more efficient. Tight junctions have a dual function as a dynamic intestinal epithelial mechanical barrier. It can not only prevent potentially harmful substances or pathogens from entering the body, but allows in nutrients, ions, and water. When the tight junctions' variation, reduce or missing, the gap of intestinal epithelial permeability will increase. Thus, bacteria, endotoxin, and macromolecular substances can pass into the systemic circulation, causing various diseases.
Several in vitro studies have demonstrated that pretreatment of intestinal epithelial cells with probiotics prevents the increase of epithelial paracellular permeability and the decrease in TER induced by pathogenic Escherichia coli or cytokines, resulting in the repair of the intestinal epithelial barrier [13,14]. This effect correlates with the dynamical regulation of apical junctional complex, particularly tight junctions consisting of tight junction proteins. Studies have also shown that probiotic bacteria prevent the decline in phosphorylation and the redistribution of tight junction proteins caused by pathogenic infection in intestinal epithelial cells [13,15]. Furthermore, Qin et al. [16] showed that LP prevented the abnormal expression and distribution of tight junction proteins, including claudin-1, occludin, JAMA-1, and ZO-1 proteins induced by enteroinvasive Escherichia coli in Caco-2 cells. Moreover, studies in vivo have demonstrated that probiotic therapy results in changes in the expression of tight junction proteins [17,18]. A recent study showed that treatment with VSL#3 (a probiotic compound) completely prevented the decreased expression and redistribution of occludin, ZO-1, and claudin-1, −3, and−5 in dextran-sodium-sulfate-induced colitis in mice [17]. Chen et al. [19] found that treatment of IL-10-/- mice with LP prevented the decreased expression of ZO-1, occludin, and claudin-1, and the redistribution of ZO-1, occludin, and β-catenin, resulting in a substantially increased TER and decreased paracellular permeability. In the present study, we investigated the effects of probiotics on the changes of apical junction complex proteins, including ZO-1, occludin, and claudin-1 in the colon of MDR-PA mice. We found that treatment of MDR-PA mice with LP prevented the decreased expression of ZO-1, occludin, and claudin-1, and the redistribution of ZO-1 and occludin, resulting in a substantially increased TER and decreased paracellular permeability. In addition, the positive correlation between paracellular permeability and inflammatory score further suggests that alterations in the intercellular junction may play an important role in probiotic therapy for intestinal inflammation.
Resta-Lenert et al. [14] found that both TNF-a and IFN-γ substantially reduced TER and increased epithelial permeability, effects prevented by Streptococcus thermophilus (ST) and Lactobacillus acidophilus (LA) or bacteroides thetaiotaomicron (BT). A Janus kinase (JAK) inhibitor synergistically potentiated effects of ST/LA or BT on TER and permeability, but p38, ERK1, 2, or PI3K inhibition did not. Finally, only probiotic-treated epithelial cells exposed to cytokines showed reduced activation of SOCS3 and STAT1, 3. Arribas et al. [20] found that pre-treatment with L. fermentum may exert its protective action against LPS-induced organ damage in mice by a combination of several actions including its antioxidant properties and by reduction of the synthesis of the pro-inflammatory TNF-α and IL-6. In our study, the expression of proinflammatory cytokines (TNF-α and IFN-γ) were increased in MDR-PA mice, whereas LP treatment prevented the increase in the proinflammatory cytokine production.
Our study showed that intestinal contents in mice after LP intervention germiculture for 24 h, were all visible for the growth of P. aeruginosa. After 3 wks treatment of LP, the drug susceptibility diameter of MDR-PA had increased compared with before such as MEM10ug, TZP110, ATM−30, and CAZ-30. Because it is short of statistics significant, we speculate about probiotics, which may change the resistance of bacteria. However, the process may be related to many factors such as environmental (flora quantity, ph, temperature, in vivo and in vitro, etc.), length of time of probiotics intervention, and the concentration of probiotics, etc. In addition, the different stages of colitis and different doses of LP may contribute to the different effects of LP on the change of bacteria resistance qualitative; however, further investigation is needed to clarify this issue.
In conclusion, probiotics can treat diarrhea of mice intestinal infection by multiple drug-resistant P. aeruginosa and protect the intestinal barrier function. Probiotics can weaken the resistance of multiple drug resistance in P. aeruginosa to some extent.
Footnotes
Author Disclosure Statement
We declare that all the authors have no conflict of interest.