
Abstract
Abstract
Background:
Trauma, major elective surgery, and overt sepsis can lead to a cascade of immunological change. A subset of these patients will have a degree of immune suppression that leads to hyporesponsive innate defenses, increasing the risk of infective co-morbidity and death. This article is an overview of monocyte impairment in the high-risk surgical patient. Specifically, our primary focus is on observations made pertaining to monocyte function and pathophysiological mechanisms underpinning this impairment. Clinical factors influencing monocyte function are also discussed.
Methods:
A Pubmed search was conducted to review aspects of monocyte impairment in the surgical patient. Search terms included “monocyte impairment,” “immunoparalysis,” and “endotoxin tolerance” cross-referenced against terms including “trauma,” “major surgery,” and “sepsis.”
Results:
Findings revealed a broad variety of monocyte defects reported in surgical patients. They ranged from altered cytokine responses, particularly ex vivo TNF-α production, to impaired antigen presentation such as depressed HLA-DR expression. The latter is the most commonly described marker of secondary infection and death. Studies of underlying mechanisms have commonly utilized a model of endotoxin tolerance with in vitro monocytes, revealing a complex array of dysregulated pathways. For our purposes, endotoxin tolerance and monocyte impairment are sufficiently similar entities to permit further study as a single subject. In the high risk patient, microRNAs (also referred to as miRNA or miR) are emerging as potential biomarkers that may modify such pathways. Creation of a reliable impaired human monocyte model could be important to all such considerations.
Conclusion:
Impairment of monocyte function continues to be predictive of nosocomial infection, multi-organ failure, and death in some surgical patients. However, the optimal marker that could identify a patient as high risk early enough, and whether it might guide potential therapy, still is yet to be proven.
I
Contrary to their null hypothesis, systematic observations of host defense mechanisms in ill patients prompted the recognition of the importance of the impaired monocyte [5]. Specifically, a subset of approximately 15% of patients exhibited subnormal monocyte function and were at greater risk of developing infection and dying [6]. This is illustrated in Figure 1, which highlights that the differences in monocyte HLA-DR expression, a cell surface marker, become increasingly predictive of a patient's outcome after days four to six following major trauma.

Percent of monocyte HLA-DR positivity and outcomes in trauma patients. Outcomes are labeled as an uncomplicated recovery (O), survivors with post-traumatic infection (□) and non-survivors (Δ) ([With permission from Hershman MJ, Cheadle WG, Wellhausen SR, et al. Monocyte HLA-DR antigen expression characterizes clinical outcome in the trauma patient. Br J Surg 1990;77:204–207.)
The ultimate goal for laboratory and clinical investigators is not just to identify such “high-risk” patients but also to augment host defenses. This principle is illustrated in Figure 2, where the addition of paternal muramyl dipeptide (MDP) in a rat model of a contaminated wound promotes containment, as opposed to permissive progression to bacteremia and the subsequent systemic deleterious consequences [7].
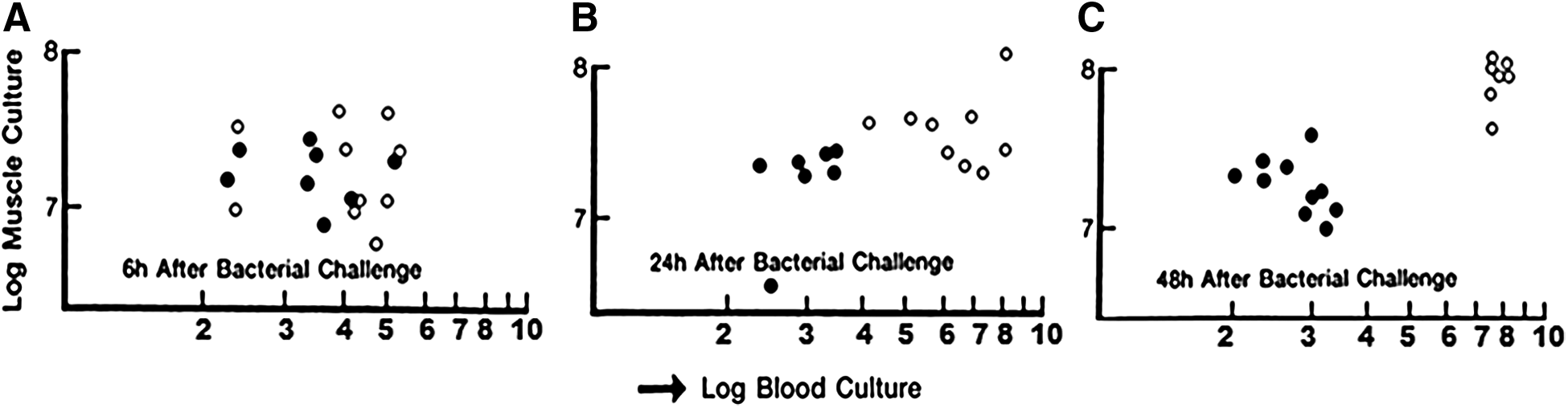
Muramyl dipeptide (MDP) augmentation of bacterial containment in a model of surgical incision contamination [7]. Infection in skeletal muscle is contained better by MDP (●) than by placebo (O). As time passes from 6 h after contamination in A, note MDP produces constant levels of localized bacteria where placebo treated animals experienced escape of bacteria into blood by 48 h (C). (With permission from Polk HC, Jr., Lamont PM, Galland RB. Containment as a mechanism of nonspecific enhancement of defenses against bacterial infection. Infect Immun 1990;58:1807–1811.)
The central factor that determines patients' fates following a major insult remains unclear. In a group of patients given the same insult, many will be “aces” and survive, but a subset will be “dogs” and not survive (Fry DE, personal communication, 1988). Firstly, genetic predisposition can lead to subnormal host defenses. Secondly, the heterogeneous nature of an insult and its severity can also be decisive. Thirdly, epigenetic change, such as in microRNA, may influence post-translational modification of important pro- and anti-inflammatory signaling. Finally, prompt and attentive care with timely surgical intervention and precise non-operative care can massively shift the balance for some patients.
The Role of the Healthy Monocyte
The monocyte is central to the innate immune system of the surgical patient. Its key roles are cytokine production, antigen presentation, and phagocytosis (Fig. 3). Early recognition of PAMPs and DAMPs from invading microbes and damaged tissue activates monocytes through surface receptors, such as extracellular toll-like receptors, and intracellular receptors, such as NOD-like receptors, RAGE receptors, and certain toll-like receptors.
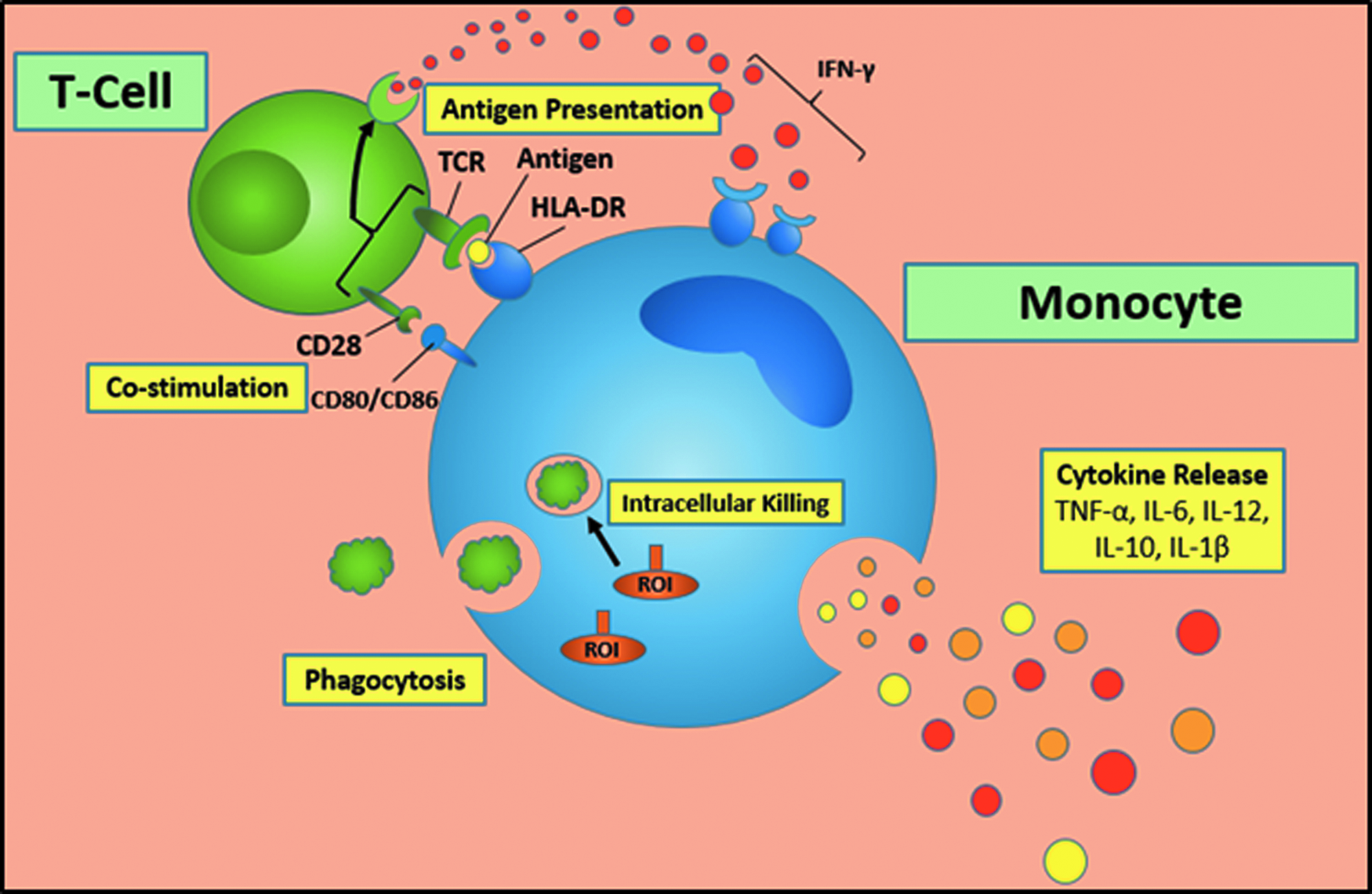
Basic functions of the monocyte. Color image is available at www.liebertpub.com/sur
This leads to the infiltration of neutrophils, traditionally the first leukocyte on the scene. These polymorphonuclear cells have the most potent phagocytic ability, which are followed by infiltrating circulating monocytes because of chemokines. These can differentiate and thus expand the number of tissue macrophages. Throughout this process, monocytes secrete cytokines, which have systemic consequences on remote organs such as the liver, gastrointestinal system, and endothelium, especially that in lung parenchyma. Furthermore, this cell–cell communication leads to a crucial link to the adaptive immune system through complex T-cell interactions. The monocyte and other antigen presenting cells allow fragments of invading bacteria, such as LPS, to be processed in vesicles and externalized through MHC II complexes, such as HLA-DR, that bind to T-cell receptors on receptive T-cells. This bi-directional interaction can also promote monocyte/macrophage activation through endogenous IFN-γ from T-cells and natural killer cells. However, in patients with subnormal immune systems or persistent antigen stimulation, this process can be blunted through the promotion of negative co-stimulatory molecules such as PD-1/PDL-1 surface molecules.
Invading microbes initially evoke a T-helper (Th) 1 response from CD4 T-cells. In this type of response, T-cells secrete IFN-γ and IL-2, further promoting and activating monocytes and macrophages in a pro-inflammatory manner. Whereas this response is linked to the “M1” pro-inflammatory macrophage, the alternative Th2 response (associated with allergens and helminths) secretes IL-10 and IL-4, promoting the “M2” anti-inflammatory macrophage.
Cytokines are secreted proteins that act in a paracrine manner to influence the differentiation and response of surrounding immune cells. Of the multitude of cytokines that monocytes secrete, the most clinically studied include the following:
• Pro-inflammatory cytokines ○ TNF-α—promotes inflammation and endothelial activation ○ IL-6—acute phase response promoting fever and T/B cell growth ○ IL-1β—fever, activation of monocyte, macrophage, T-cell activation • Anti-inflammatory cytokines ○ IL-10—suppresses monocyte function
Cytokines influencing monocyte behavior is discussed below. Although monocytes do phagocytize bacterial pathogens, defects in their phagocytic ability appear to be less important to the fate of the surgical patient when compared with altered cytokine release and antigen presentation.
Monocytes are now considered to be of three types [8]:
1. The classical monocyte accounts for the majority of circulating monocytes, expressing high CD14, medium HLA-DR, but not CD16 (CD14++ CD16-). These cells produce both TNF-α and IL-10. 2. Non-classical monocytes express lower CD14, but high CD16 and HLA-DR (CD14+CD16++). They secrete TNF-α, but not IL-10. 3. Finally, intermediate monocytes express high CD14, but low CD16 (CD14++ CD16+). This subset has gained increasing recognition over the last 5 y and is noted to increase IL-10 production. It has been correlated with sepsis, myocardial infarction, and early but not metastatic colorectal cancer [9].
It is worth noting that monocytes, macrophages, and dendritic cells are all myeloid-derived with largely similar expressions of toll-like receptors. Although this review focuses on the monocyte, many of the concepts discussed regarding physiology, pathology, and therapy can be extended to all the above.
Clinical Scenarios and the Monocyte Response
Major trauma and burns
As post-traumatic care (and indeed all critical care) improved, it became apparent that some survivors of major trauma were susceptible to later infection. Evidence grew that this may be a consequence of depressed host defenses [10]. Our systematic study of various facets of the innate immune system revealed that monocyte antigen presenting ability was highly predictive of infection following major trauma [5]. It was apparent that not only low absolute monocyte HLA-DR expression, but also the inability to up-regulate this MHC II surface marker in response to endotoxin was also predictive of morbidity and death. Since then, studies focusing on HLA-DR as a marker of monocyte function demonstrate that this remains consistently predictive for poor outcomes following trauma and burns (Table 2) [5,6,11–25]. Increasingly, CD14+ HLA-DR- cells are described as “myeloid derived suppressor cells,” and this phenotype has also been observed in the orthopedic patient following intramedullary nailing following fracture of the tibia [26].
Authors' group
Review article
The revolution of cytokine-related research prompted another aspect of monocyte function to be examined: Ex vivo cytokine production in response to endotoxin, both in “whole blood” and in isolated peripheral blood mononuclear cells. In general, early high IL-6 production and subsequent lower TNF-α production appear to characterize this dysregulation [27]. Furthermore, the role of the anti-inflammatory IL-10 has long been debated, but it remains that IL-10 acts to inhibit pro-inflammatory pathways. Although a homeostatic balance is critical, IL-10 can promote a suppressed monocyte phenotype [28].
Findings of the Glue Grant 1 project relating to monocyte function
A highly funded multi-center project, the Glue Grant, aimed to take a “systems approach” to the host inflammatory response to trauma and burns, primarily focusing on gene expression studies. This design was to lessen the reductionist approach of multiple groups focusing on one single aspect of the host defenses, which was seen as likely to fail given the redundancy of the immune system. It is worth noting that the patients studied were critically ill, generally being hypotensive, having received blood products, and demonstrating some form of organ dysfunction. Selected papers from this effort pertaining to monocyte impairment are discussed below.
Xaio et al. [29] sequentially studied genome wide expression of messenger RNA in 167 severely injured trauma patients up to 28 d after injury. In addition, patients with greater than 20% burns and healthy individuals, with and without low dose LPS treatment, were analyzed. Results reflected a change in over 80% of gene expression, termed a “genomic storm.” This response to trauma was non-specific, with 98% of the same genomic changes observed in burn patients and 88% observed in experimental endotoxemia. A complicated recovery was associated with the up-regulation of innate genes including IL-6 and IL-10, with a simultaneous down-regulation of humoral immunity genes such as those relating to antigen presentation. Importantly, the authors demonstrated that contrary to conventional thinking, leukocyte genomic responses of both pro- and anti-inflammatory pathways were simultaneous rather than a so-called “second hit” or insult causing the anti-inflammatory component [29]. Laudanski et al. [30] examined both monocyte and T-cell pathways of 18 trauma patients using high throughput screening of gene expression. Interestingly, the response to trauma was specific to the cell type, with only 11% of pathways common to both monocytes and T-cells, such as a down-regulation of the transcription factor NFκB. This further demonstrated a down-regulation of antigen presentation genes such as HLA-DR and CD86, with an increase in monocyte negative co-stimulatory genes such as PDL2. In keeping with the down-regulation of antigen presenting pathways in monocytes, T-cells demonstrated molecular choreography with reciprocal increases in genes relating to apoptosis and anergy [30].
In a more recent study from the same dataset, 8% of the 1595 subjects included had proven gram-negative bacteremia. These 127 patients had an increased risk of death (21%) compared with those without infection (11%). Gene expression of these patients was compared with those whom did not contract infection, showing certain pro-inflammatory pathways to be similar 12 h following injury. However, at 96 h after injury, IL-1β and IL-2RB were markedly decreased in the former group. Furthermore, genes relating to monocytes, such as HLA-DR and CD163, were similarly increased at 12 h after injury, but at 96 h, a substantial decrease in the these genes was apparent in the group that went on to develop gram-negative bacteremia [31]. This project has increased our understanding of the pathways involved in host defenses following major trauma. Perhaps not surprisingly given the recognition of such complexity, the surgeon remains ill-equipped to confidently or specifically modulate this response.
Inflammatory networks following trauma
Since the Glue Grant project, other groups have adopted a ‘systems biology’ approach to better understand inflammatory networks after major trauma. The Billiar group studied 472 survivors of blunt trauma for circulating plasma cytokines and chemokines. The findings were analyzed using emerging computational methods, which reflected that various patterns were demonstrable in the first 24 h, much earlier than the diagnosis of infection made 7 d after injury. Raised markers in the patients who later developed infection include HMGB1, IL-6, and IL-1RA, to name a few. Dynamic Bayesian networks inferred that IL-6 was affected by various chemokines such as MCP-1 and MIG [32,33].
Major elective surgery
Major abdominal operations and general anesthesia have long been associated with post-operative immunosuppression. Factors such as increasing age, prevalence of diabetes mellitus, and immunosuppressive medications (neoadjuvant chemotherapy, glucocorticoids, and monoclonal antibodies such as anti-TNF-α) increase the relevance of this concept today. However, the prevalence of impaired monocyte function also extends to the patients of neurosurgeons, transplant surgeons, and cardiac surgeons [34–36]. More recently, Albertsmeier et al. showed that major abdominal surgery leads to an expansion of CD14+ HLA-DR−cells. This group of monocytes secreted TNF-α and IL-6, but when co-cultured with T-cells, led to decreased T-cell IFN-γ secretion [37]. As discussed later, studies in the last few years have demonstrated how surgical approaches can also influence monocyte behavior. Furthermore, dysregulation of pro-inflammatory pathways in monocyte subsets, such as NFκB in intermediate monocytes, led to greater TLR and IL-6 expression that was associated with increased infective complications [13,18].
Overt sepsis
Major infection is the most common clinical event associated with monocyte impairment, namely severe sepsis and septic shock. Although the majority of studies use flow cytometry to quantify either the percentage of monocytes expressing HLA-DR or mean fluorescence intensity, more recent studies show that gene expression of HLA-DR is also predictive of infective complications [12,38]. Furthermore, bench-to-bedside studies of monocyte impairment reveal dysregulated gene expression patterns, such as S100A8/9, TNFAIP6, and IRAK-M (as well as TNF-α, IL-10 and HLA-DR), which have been suggested to “monitor efficacy” of IFN-γ therapy [39–41].
There has been increased interest over the past decade in the differential expression of negative co-stimulatory molecules in patients with sepsis. This has been in part because of the recognition of T-cell anergy and apoptosis as a driver of immune suppression, which is also observed in some cancers [42]. Monocyte co-stimulatory antigens such as CD86 and PDL-1 correspond to T-cell CD28 and PD-1, respectively, which may be considered “on” and “off” switches in addition to the central HLA-DR–T-cell receptor interaction. The potential importance of negative co-stimulatory molecules has been highlighted by reported improved mortality rates in murine models of surgical sepsis from PD-1/PDL-1 blockade [43,44]. Various clinical observations have found increased monocyte PDL-1 and increased T-cell PD-1 expression in infected patients. In one study of 64 patients with septic shock, of which 42% had an abdominal source, patients had increased expression of monocyte PDL-1 and T-cell PD-1 compared with healthy control groups, which was also predictive of secondary nosocomial infection [25]. In another study of 90 critically ill surgical patients, blood samples withdrawn within 24 h of admission to the intensive care unit (ICU) showed increased monocyte PD-1 expression, correlating with APACHE II scores [45]. In addition, certain PD-1 polymorphisms have been associated with greater mortality rates in 221 patients diagnosed with sepsis or septic shock, of which lung was the most common source [46].
Clinical Factors Influencing Monocyte Function
The practicing surgeon caring for sick surgical patients may adversely influence monocyte function unknowingly, as summarized in Table 3 [47–56]. Experimental models have shown that failing to protect patients from unintentional hypothermia may promote the pro-inflammatory response through increased microRNA-155 expression and subsequent greater TNF-α and lower IL-10 production [47]. The once popular strict glucose control in critical care environments may risk harmful hypoglycemic episodes, but also blunt cytokine and bactericidal properties of the monocyte [48]. Supplemental oxygen, to an inspired oxygen concentration of 80%, appears to have mixed effects on the monocyte and increases reactive oxygen species in particular [49]. The immunosuppressive effects of blood transfusion have been recognized for many years, but recent studies implicate small circulating biomolecules, such as protein-bound or exosomal microRNA, which may also inhibit monocyte HLA-DR expression [51]. The role of minimally invasive surgery on monocyte function has remained controversial [57]. Recent studies have suggested that minimally invasive approaches by laparoscopy or in some cases robotics are less immunosuppressive, as compared with major open colorectal and liver resectional surgery [52–56].
Defining and Studying the Impaired Monocyte
Clinical studies have shown that monocyte impairment is not common in most surgical patients and pertains to only some previously unsuspected patients. Therefore, it becomes difficult to adequately fund clinical studies to examine the mechanisms behind this pathology. Over the last 15 years, both in vitro and in vivo murine models have been used to study endotoxin tolerance. Endotoxin tolerance was discovered by Beeson, who found that repeated injection of endotoxin led to progressively decreased febrile response [58]. We now know that exposure to endotoxin, often LPS, stimulates various negative regulators to inhibit a subsequent response when compared with a naïve cell. This has been shown to be akin to the impaired monocyte response observed in the critically ill surgical patient [59]. We consider monocyte impairment and endotoxin tolerance in monocytes to be a similar phenomenon. Various studies have shown “cross-tolerance” in that the same molecular mechanisms behind such an impairment also occur with other so-called “priming” agents that include the following:
• PAMPs/DAMPs ○ Lipoteichoic acid (TLR2) ○ Poly (I:C) (TLR3) ○ High mobility box group 1 (TLR2 & 4) • Cytokines ○ IL-1β ○ IL-6 ○ TNF-α ○ IL-10 [60]
Common models include isolated primary cells such as CD14 monocytes and PBMCs as well as cell lines such as THP1 and U937 monocytes and RAW 264.7 macrophages. In vivo models expose mice to low dose intraperitoneal LPS followed either by ex vivo stimulation or in vivo cecal ligation and puncture. Human in vivo models have used subcutaneous LPS followed by ex vivo leukocyte stimulation [61]. The most accepted in vitro outcome of the LPS pre-treated “impaired” cell is reduced TNF-α production compared with naïve cells, measured either by supernatant protein secretion by ELISA or gene expression of TNF-α messenger RNA. Other cytokines have been reported variably [60]. Although reported less frequently, this approach is believed to represent monocytes with reduced antigen presenting capabilities [62,63].
Pathophysiological mechanisms behind the impairment of the monocyte response are complex and influence various molecular pathways (Fig. 4) [60]. In toll-like receptor signaling, the MyD88-dependent pathway is inhibited at various levels, such as sMyD88 and IRAK-M that inhibits TRAF6. Furthermore, A20 increases and thus inhibits TRAF6. It is at this level that Pellino-1, a positive regulator, is decreased in impaired cells [64]. There is variable reporting on the dysregulation of the IκK subunits, but recently, monocyte impairment has been associated with disruption of the interaction between IRAK1 and IκK-γ (also known as NEMO) [65].
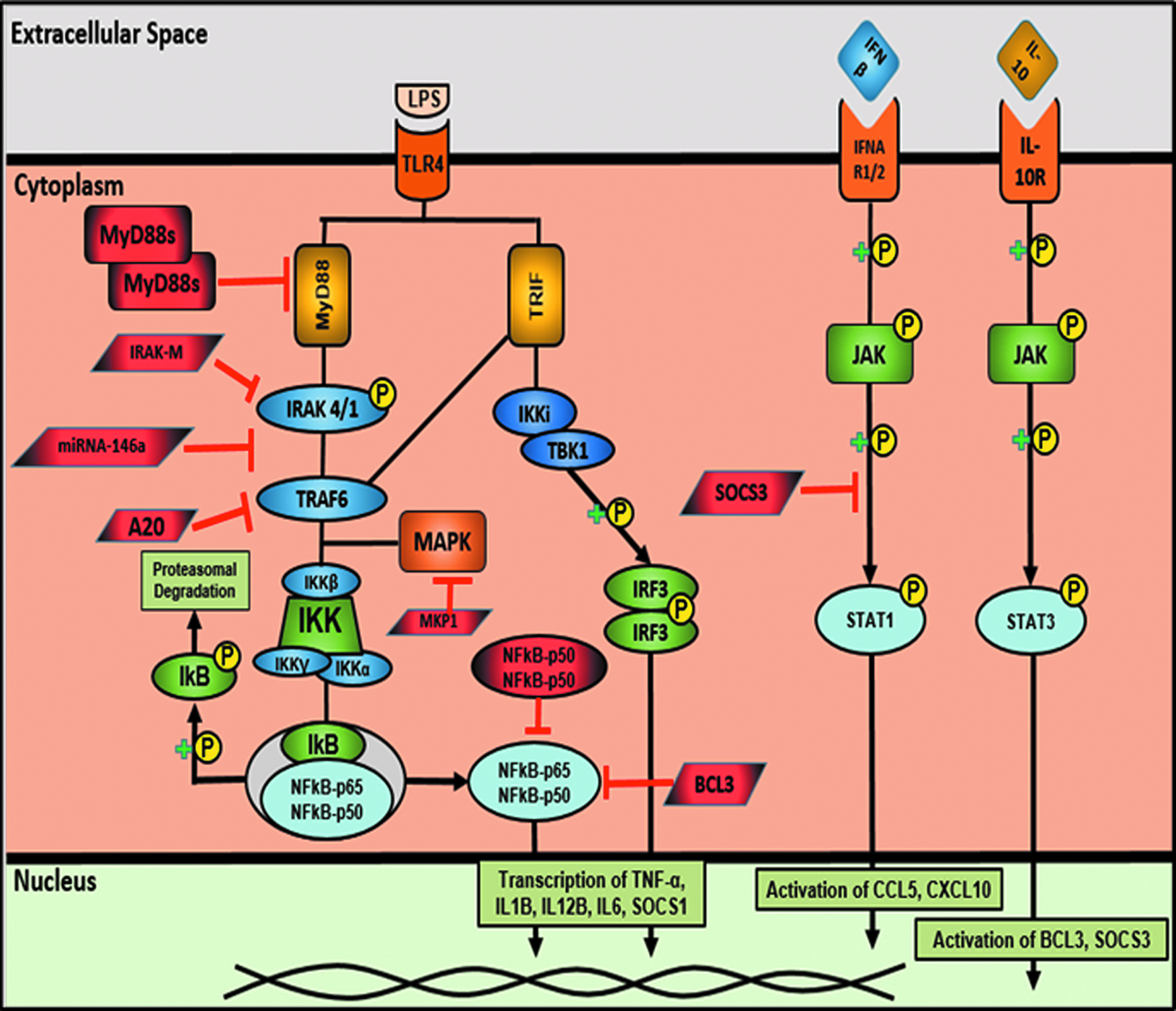
Negative regulators involved in monocyte impairment (endotoxin tolerance). (With permission from Biswas SK, Lopez-Collazo E. Endotoxin tolerance: New mechanisms, molecules and clinical significance. Trends Immunol 2009;30:475–487.) Color image is available at www.liebertpub.com/sur
This leads to subsequent increases in concentrations and decreased degradation of IκB in impaired monocytes, which act as the “brakes” in the NFκB pathway. Therefore, there is decreased p65 phosphorylation and subsequent translocation to the nucleus, in part explaining the decreased gene expression of pro-inflammatory cytokines. Furthermore, the non-canonical NFκB pathway has been reported to be involved, with impaired cells having increased expression of RelB, c-Jun, and the homodimer of p50-p50 of NFκB rather than the more commonly expressed p-65-p50 heterodimer [66].
The MAPK pathway is also dysregulated, with evidence of decreased TAK1 phosphorylation in impaired cells, which leads to decreased MAPK p38 phosphorylation [65]. Other known negative regulators that are not discussed here include SOCS1, SOCS3, BCL3, PPAR-γ, and CREB [60].
MicroRNAs are short, non-coding RNA molecules involved in post-transcriptional regulation of gene expression. Various microRNAs are dysregulated in monocyte impairment, with microRNA-146a being the most studied. MicroRNA-146a increases as a physiological response to LPS and subsequently inhibits TRAF6 and IRAK1. Murine knock-out or antagomiR-mediated transfection rescues the impaired response. Further microRNAs shown to influence monocyte impairment include let-7a, miR-223, 221, 9, 150 and 132 [60].
However, despite the manipulation of numerous negative regulators demonstrated in vitro to improve the monocyte response, none of these are known to have gained scientific momentum or reached the bedside. Such complexity highlights the redundant nature of our evolved immune system. Although in vitro monocyte cell culture allows molecular pathways to be elucidated, these models are limited by the lack of surrounding cells that are obviously present in vivo. Furthermore, murine studies of sepsis, such as CLP, have for many years raised unfulfilled hope in potential immunotherapy for the impaired monocyte, but they have not yet translated to clinical success. One reason might be that the immune system is not completely conserved between human beings and other species such as mice. Murine models are overtly oversimplified—there are no additional parameters such as blood transfusion, fluid resuscitation, antibiotic therapy, and co-morbidity. Finally, the genetic variation in the patient population is obviously not represented in the narrow pool of mice and cell lines.
Conclusion
Despite a plethora of other biomarkers associated with hard outcomes in surgical patients, observations focusing upon monocyte function have stood a considerable test of time. Many of these, including monocyte HLA-DR, have been reproducible in their ability to reflect underlying immunological impairment following major insults. We have observed that even this marker does not fully stratify patients until 4–6 d after the initial insult, which may be too late for a potential therapy to influence the patient trajectory. MicroRNA assessment could provide an earlier marker or indeed be part of a panel of markers that reflect the true high-risk patient. Despite the daunting task, the search for earlier and reliable markers for patient identification and monitoring response to therapy is warranted through tailored, personalized immunotherapy to shift the balance in favor of the high-risk or immunodeprived surgical patient.
Footnotes
Author Disclosure Statement
No competing financial interests exist.