
Abstract
Abstract
Background:
Despite significant advancements in surgical protocols and biomaterials for orthopedics, peri-prosthetic joint infection (PJI) remains a leading cause of implant failure. Staphylococcus aureus nasal colonization is an established risk factor for PJI, with methicillin-sensitive S. aureus a leading cause of orthopedic implant-related infections. The purpose of these in vitro studies was to investigate the antibacterial activity of a tailored bacteriophage cocktail against planktonic and biofilm-associated S. aureus.
Methods:
The S. aureus strains (n = 30) were screened for their susceptibility to a library of S. aureus-specific bacteriophage (n = 31). Five bacteriophage preparations that demonstrated bactericidal activity against >90% of S. aureus strains tested were combined as a StaPhage cocktail and assessed for their antibacterial activity toward planktonic and biofilm-associated S. aureus, with biofilms established on three-dimensional-printed porous titanium scaffolds.
Results:
StaPhage treatment immediately after bacterial inoculation inhibited growth of S. aureus by >98% in eight hour cultures when multiplicity of infection of phages to bacteria was greater than 1:1 (p < 0.01). Viable bacterial numbers within biofilms on titanium surfaces were significantly reduced (6.8 log10 to 6.2 log10 colony forming units [CFU]; p < 0.01) after exposure to the StaPhage cocktail, in vitro. No significant reduction was observed in biofilms exposed to 100 times the minimal inhibitory concentration of cefazolin (log10 6.81 CFU).
Conclusions:
Combined, these data demonstrate the in vitro efficacy of S. aureus-specific bacteriophage cocktails against S. aureus growing on porous titanium and warrant further in vivo studies in a clinically relevant animal model to evaluate the potential application of bacteriophage in the management of PJI caused by S. aureus.
P
The formation of bacterial biofilms is intrinsic to the pathogenesis of PJI [5–7]. The ability of bacteria such as S. aureus to form biofilms on orthopedic implants makes diagnosis and treatment of PJI extremely challenging [3,7]. Biofilms are complex microbial communities encased in a self-produced polysaccharide matrix that serves to protect bacteria from the host immune response and antibiotic agents, thus facilitating persistent infection and increased likelihood for the emergence of antibiotic-resistant bacterial strains [5]. The alarming global rise in antibiotic-resistant bacterial strains is driving an urgent need for the development of innovative strategies to combat bacterial infections [8]. Bacteriophage therapy offers an alternate strategy for the management of PJI, particularly where there are limited effective antibiotic agents available.
Bacteriophages, or phages, are naturally occurring viruses that infect only bacteria without harming eukaryotic cells [9]. Lytic phages infect and replicate within a specific host bacterium causing lysis and death of the bacterial cell and release of large numbers of newly assembled virions that can attack more target bacteria, perpetuating the cycle for as long as there are live bacterial target cells present. Therefore, unlike antibiotic agents that decrease in concentration below the surface of bacterial biofilms, phages can penetrate into biofilms making them particularly useful for managing chronic, persistent bacterial infections [10].
Phage cocktails, mixtures of individual phage types, have been used to broaden the spectrum of activity against pathogenic bacteria such as S. aureus [11]. Phage therapy has been standard clinical practice for management of bacterial endocarditis, periodontitis, gastrointestinal, respiratory, bone and joint, and urinary tract infections in Eastern Europe for almost a century [12]. Although the clinical application of phages declined in Western countries with the advent of antibiotic agents, phage therapy is becoming increasingly considered globally as an attractive adjunct or alternative treatment to antibiotic agents in response to the increase in antimicrobial resistance [9,12].
Staphylococcus aureus nasal colonization is an established risk factor for PJI after arthroplasty [13]. Molecular studies have demonstrated the genetic relatedness of S. aureus strains from surgical infections with those colonizing skin in the pre-operative period [13,14]. Methicillin-sensitive S. aureus (MSSA) causes approximately 45% of PJI cases, with MSSA infections 2.5 times more common than PJI caused by methicillin-resistant S. aureus (MRSA) [3,15]. Despite antibiotic sensitivity, the ability of MSSA strains to establish biofilms on the surface of orthopedic materials makes eradication extremely challenging. Therefore, the purpose of the current study was to investigate the antibacterial activity of S. aureus-specific phages against planktonic and biofilm-associated MSSA strains recovered from the skin of patients undergoing total joint replacement, as proof-of-concept in vitro studies for future potential therapeutic application of phage for management of PJI caused by S. aureus.
Methods
Bacteria and bacteriophages
A panel of S. aureus strains (n = 30) recovered from anterior nares of patients (n = 94) undergoing total knee or hip arthroplasty was used in the current study after approval from the Institutional Ethics Committee. The S. aureus strains were isolated from pre-operative nasal swabs and identified using standard microbiologic and biochemical techniques. Methicillin resistance was confirmed using a rapid immunochromatographic PBP2a culture colony test (Alere, Australia). Glycerol stocks of S. aureus isolates were stored at −80°C until further analysis.
The S. aureus-specific phages (StaPhage) (n = 31) preparations used in the current study were a kind gift by Dr. L. Elliot (AusPhage Pty Ltd). Lytic StaPhage were isolated from sewage water, triple plaque purified, and characterized using methods described previously [16]. Host sensitivity range was determined for each preparation using spot testing of serially diluted lysates against a panel of veterinary and clinical isolates of S. aureus as part of the routine isolation procedure [17]. Phage classification was confirmed by morphology using transmission electron microscopy (TEM, Tecnai 12, FEI, The Netherlands). The StaPhage preparations provided were from the family Myoviridae (Fig. 1a, b) and contained 1 × 107–9 plaque-forming units (PFU) per mL.
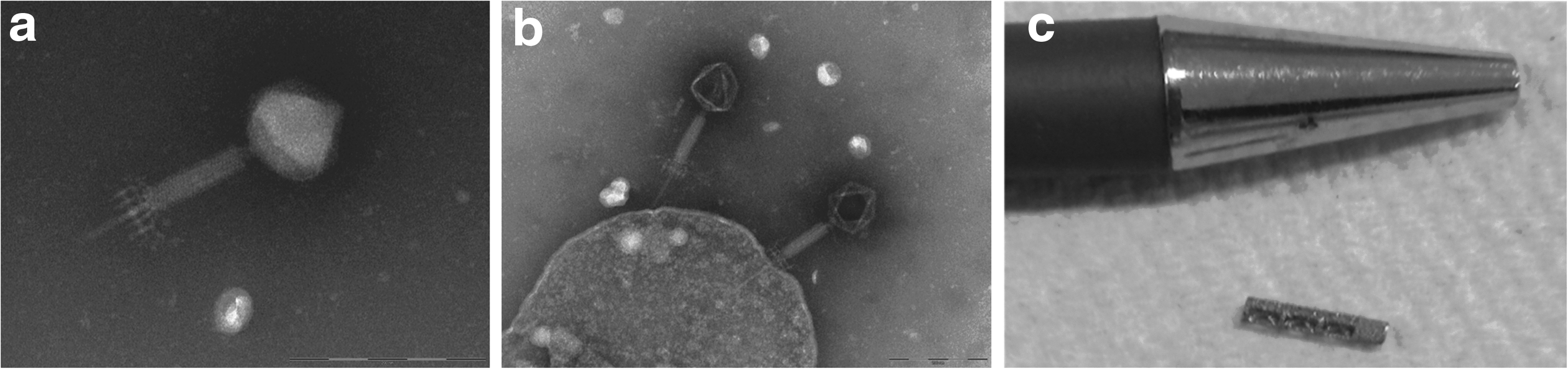
Transmission electron microscopy (TEM, Tecnai 12, FEI, The Netherlands) image of representative phage preparations, (
S. aureus sensitivity screening
Akin to antimicrobial susceptibility testing of bacterial isolates, phage sensitivity screening was performed using standard spot tests [17]. The S. aureus strains (n = 29 MSSA, n = 1 MRSA) were screened against a panel of individual StaPhage preparations (n = 31) to identify phage preparations with a broad spectrum of lytic activity for subsequent inclusion in a StaPhage cocktail. Propagating strains of S. aureus were used in parallel to test strains to confirm viability, activity, and concentration of StaPhage preparations. Bacterial sensitivity was considered where confluent, semi-confluent, opaque lysis, or individual plaques were observed in the bacterial lawn. Data are presented as the percentage of S. aureus strains tested that demonstrated sensitivity to each individual StaPhage preparation.
Phage efficacy against planktonic S. aureus
Based on in vitro sensitivity screening assays, five lytic StaPhage preparations (StaPh_1, StaPh_3, StaPh_4, StaPh_11, and StaPh_16) were selected for further analysis. To compare the inhibitory effect of individual StaPhage preparations with a pooled cocktail of the individual preparations on bacterial growth, two MSSA strains (ORI16_C02N and ORI16_025) were cultured in the presence of StaPhage at a multiplicity of infection (MOI) of 10 (phage):1 (bacteria). To compare the effect of MOI on inhibition of planktonic growth of S. aureus, the StaPhage cocktail was added to cultures of two additional MSSA strains (ORI16_002 and ORI16_096) at MOI 0, 0.01:1, 0.1:1, 1:1, or 1:10. Individual StaPhage preparations or a cocktail containing an equal concentration of the five pooled StaPhage preparations were added to log-phase S. aureus cultures (105 CFU), at the indicated MOI.
Growth kinetics (37°C, 50 rpm) were monitored over an eight hour period with hourly measurement of optical density (OD, 600 nm). Data are expressed as mean OD or, where indicated, as percentage reduction in bacterial growth compared with untreated cultures according to the formula: (ODuntreated − ODtreated)/ODuntreated *100. To enumerate viable bacteria, parallel culture plates were prepared, and at four hours, aliquots were removed from untreated and treated cultures, serially diluted, and plated onto tripticase soy agar (TSA) with colonies enumerated after overnight incubation.
Biofilm-forming capacity of S. aureus
The S. aureus strains (n = 10) were screened for their ability to form biofilms, where each isolate was categorized on strength of biofilm formation in comparison with a reference S. aureus strain of known biofilm-forming capacity (ATCC 25923) [18]. Biofilms were established in microtiter plates using log phase suspensions of S. aureus strains (5 × 104 CFU/mL, tryptic soy broth (TSB) with 0.5% glucose). After incubation (48 h, 37°C), biofilm formation was assessed using a colorimetric crystal violet technique and measurement of OD, 590 nm [19]. The cutoff optical density (ODc) for each plate was defined as three standard deviations above the mean OD of the blank wells. Thresholds to distinguish weak, moderate, and strong biofilm formation were set at 2 × ODc or 4 × ODc, respectively.
Phage efficacy against biofilm-coated titanium surfaces
Titanium is commonly used in prosthetic joint components, with three-dimensional (3D)-printed titanium increasingly being used in orthopedic implants [20]. To assess the efficacy of phages against biofilms growing on a clinically relevant orthopedic material, two MSSA strains (ORI16_C02N and ORI16_C02J) that demonstrated strong biofilm-forming capacity on polystyrene surfaces were selected for further analysis of their ability to colonize the surface of customized 3D-printed, porous titanium cylindrical scaffolds (Fig. 1c, 70% porosity, 5 mm × 1.6 mm) designed for use in our rat model of knee implant surgery (a kind gift from R. Wood, Stryker Orthopaedics).
ORI16_C02N was isolated from the anterior nares of a patient who had undergone total knee arthroplasty (TKA) with uncomplicated recovery; however, the patient presented with acute-onset septic arthritis in the operated knee three years post-TKA. The ORI16_C02J was recovered from synovial fluid of the implanted knee of this patient before debridement with implant retention and intravenous flucloxacillin therapy. Staphylococcal single neucleotide polymorphism based-typing [21] confirmed MSSA status, with both strains found to be of the same clonal complex, CC78, with identical binary profiles for pvl, cna, sdrE, pUB110, and pT181.
Biofilms were established by placing sterile implants into log phase suspensions of S. aureus (104 CFU) for 48 hours, with a media change performed after four and 24 hours. Cefazolin is frequently used as a first-line antibiotic agent for management of post-surgical infections caused by MSSA in orthopedics [3]. To compare in vitro efficacy of cefazolin and StaPhage cocktail, biofilm-coated implants were left untreated or treated with cefazolin (50 mg/mL, 100X minimum inhibitory concentration [MIC]) or StaPhage cocktail (2 × 106 PFU) for 48 hours, with media changed after 24 hours of culture.
After 48 hours of treatment, implants were sonicated (5 × 5 min rounds, 100% power, XUB12 Grant Ultrasonic bath) in recovery media (phosphate buffered saline [PBS], pH 7.2 with 0.1% Triton-X-100 and 0.15% ethylenediaminetetraacetic acid [EDTA]). Recovery media were centrifuged (3000g, 10 min, 4°C), bacteria re-suspended in sterile PBS, and serial 10-fold dilutions plated in duplicate onto TSA to enumerate colonies after 24 hours. Data are expressed as log10 CFU per implant.
Scanning electron microscopy
StaPhage-treated, cefazolin-treated, and untreated biofilm-coated titanium implants were fixed in 2.5% glutaraldehyde in 0.1M sodium cacodylate buffer, sputter-coated with gold, and visualized by scanning electron microscopy (SEM, JSM5410LV, Jeol Ltd).
Statistical analysis
Statistical analyses were performed using GraphPad Prism for Mac software (version 7). All experiments were conducted in triplicate or quadruplicate. Results are expressed as mean ± standard error (SE). Two-way analysis of variance (ANOVA) with Holm-Sidak multiple comparison test was used to compare in vitro antibacterial activity of StaPhage preparations between S. aureus strains. Comparison between treatment groups was performed using one-way ANOVA with the Levene test for equality of variances and Tukey post-hoc analysis for pairwise comparison. Differences corresponding to p < 0.05 were considered significant.
Results
Sensitivity of S. aureus nasal isolates to phage
Phage sensitivity screening was performed to identify StaPhage preparations with broad activity against S. aureus strains for use in subsequent in vitro efficacy assays. Approximately half of the StaPhage preparations tested (17 of 31) demonstrated lytic activity against >90% of S. aureus strains tested (Fig. 2). Of the 30 S. aureus strains isolated from the anterior nares of TKA patients, one strain was identified as MRSA positive (ORI16_041). Twenty-five of the 31 (80.6%) StaPhage preparations demonstrated lytic activity against this MRSA strain. Given that MSSA is more commonly associated with PJI [3,15], however, further investigations of the effectiveness of StaPhage preparations were based on the MSSA strains recovered.
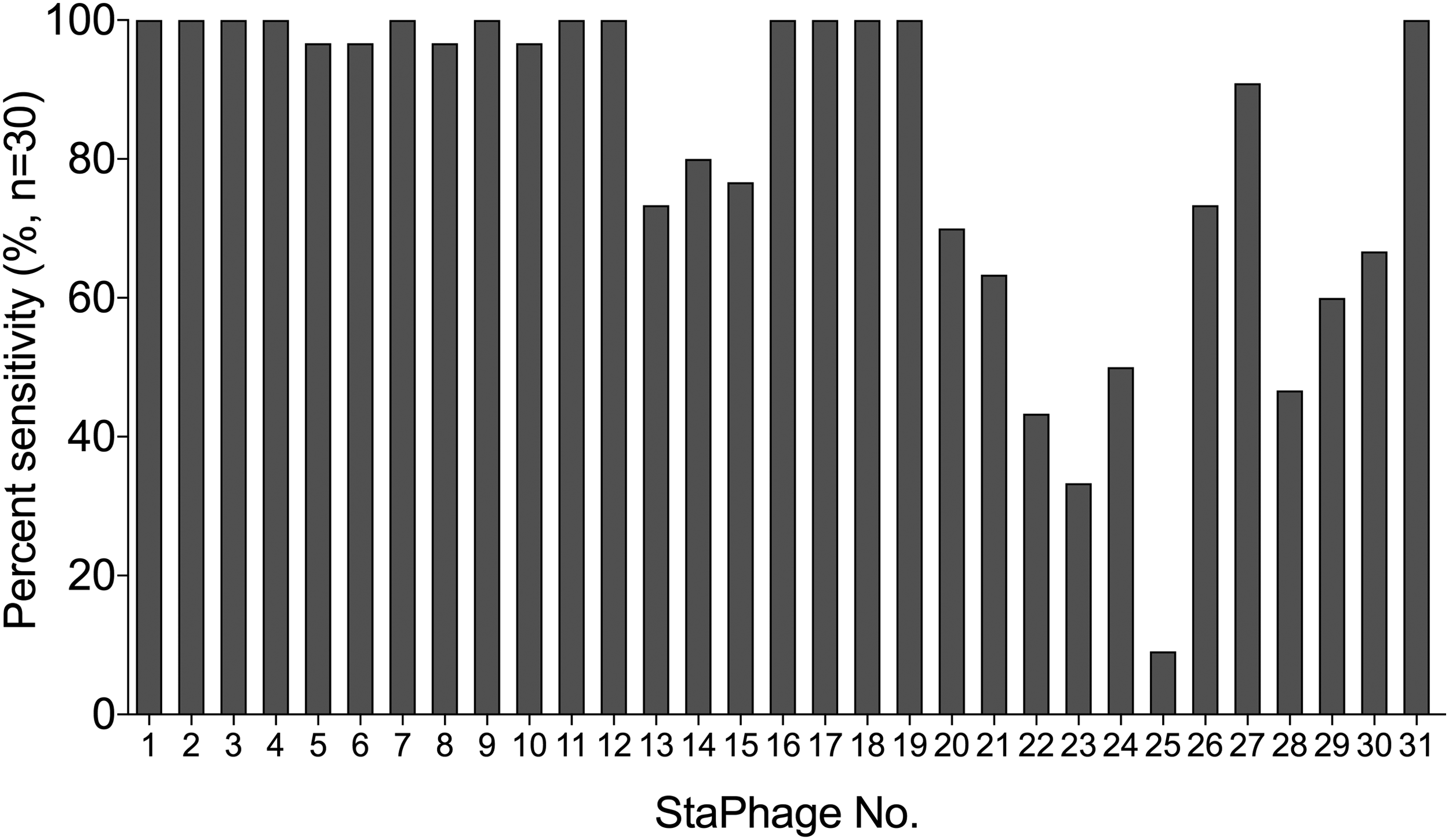
Phage sensitivity screening was performed against a panel of Staphylococcus aureus strains (n = 30) isolated from pre-operative nasal swabs of patients undergoing joint replacement surgical procedures. More than 90% of the S. aureus strains screened showed sensitivity toward 17 of the 31 phage preparations tested.
Inhibition of S. aureus growth
To compare the inhibitory effect of individual StaPhage preparations and a pooled cocktail on bacterial growth, two MSSA strains (ORI16_C02N and ORI16_25) were cultured in the presence of StaPhage at an MOI of 10:1. Differences were observed in the antibacterial activity of individual StaPhage preparations against the two S. aureus strains (Fig. 3a). In vitro bacterial growth of ORI16_C02N and ORI16_025 was inhibited by up to 98.3% and 88.3% in the presence of at least one StaPhage preparation, respectively. There was no additive effect observed, however, on the reduction in bacterial growth of ORI16_C02N or ORI16_025 under these in vitro conditions when individual StaPhage preparations were pooled as a cocktail.

A dose-response inhibition effect was observed on in vitro growth of two additional MSSA skin-associated strains, ORI16_002 and ORI16_096, after addition of the StaPhage cocktail at increasing MOI (Fig. 3b,c). Bacterial growth was reduced by 98.9% and 98.5% for ORI16_002 (p < 0.01) and ORI16_096 (p < 0.01), respectively, where the StaPhage cocktail was added at MOI 1:1. Enumeration of viable bacterial numbers confirmed a significant decrease in ORI16_002 (p < 0.01) and ORI16_096 (p < 0.01) in the presence of StaPhage cocktail at an MOI 10:1 (Fig. 3c). Combined, data demonstrate that StaPhage is effective at inhibiting in vitro growth of S. aureus when cultured in the presence of MOIs greater than 1:1.
Anti-biofilm activity of StaPhage cocktail
Having demonstrated the efficacy of StaPhage in inhibiting planktonic growth of S. aureus in vitro, we subsequently sought to investigate the antibacterial activity of StaPhage toward S. aureus biofilms. Five of 10 (50%) of the S. aureus strains screened demonstrated strong in vitro biofilm-forming capacity on polystyrene surfaces (Fig. 4). Two MSSA strains derived from a TKA patient in whom PJI had developed (ORI16_C02N, ORI16_C02J) were selected for further analyses based on previous demonstration of their in vitro susceptibility to the StaPhage cocktail in planktonic form, the strength of in vitro biofilm formation, and in vitro susceptibility to cefazolin in planktonic form (Fig. 5a).
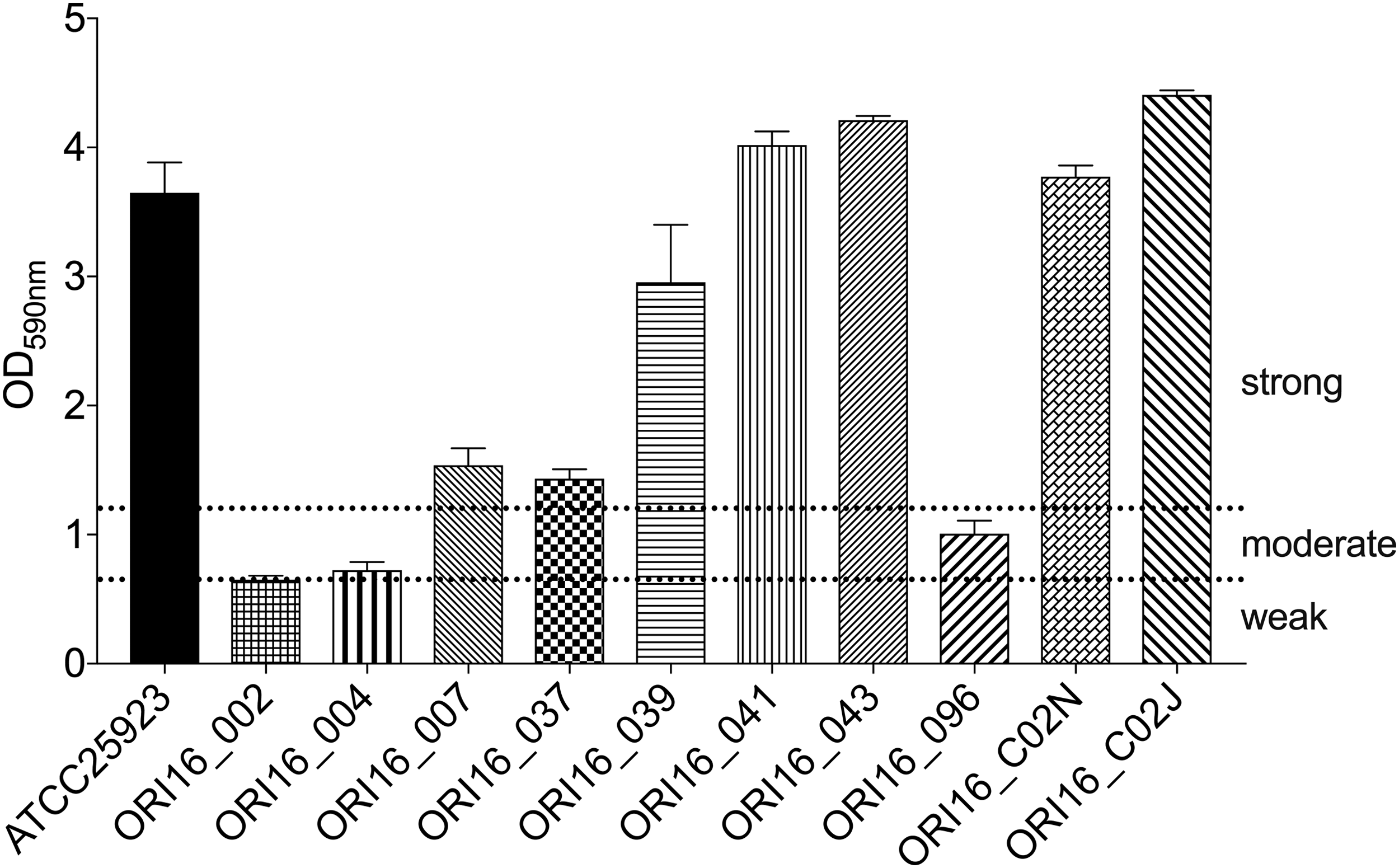
Biofilm formation of S.taphylococcus aureus strains on polystyrene surfaces after 48 hour growth at 37°C, in vitro. Biofilm formation was quantified by measuring the optical density at 590 nm (OD590) of dissolved crystal violet. Means and standard error of the mean values of OD590 values of five replicate wells for each bacterial strain are shown. A S. aureus reference strain (ATCC25923) with known biofilm-forming ability was included as a positive control. The cutoff OD (ODc) was defined as three standard deviations above the mean OD of the negative control. Biofilm production was classified as follows: ODc < OD <2 × ODc = weak biofilm production; (2 × ODc) < OD <4 × ODc = moderate biofilm production; and 4 × ODc < OD = strong biofilm production.
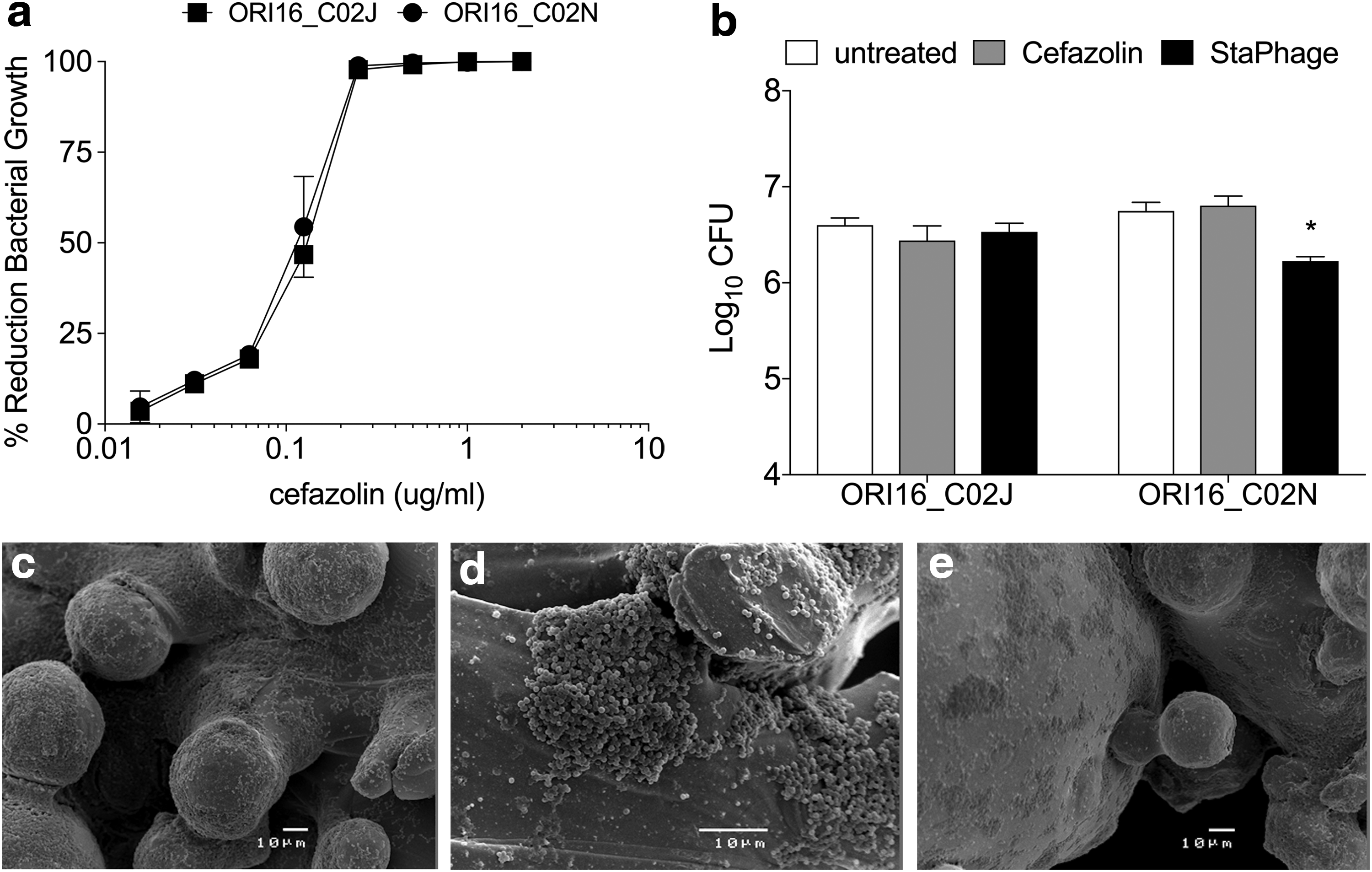
Anti-biofilm activity of StaPhage cocktail.
After 48 hours in culture with titanium implants, levels of ORI16_C02N and ORI16_C02J within biofilms reached 6.3 and 6.2 log10 CFU, respectively. Although both S. aureus strains were shown to be sensitive to cefazolin, viable bacterial numbers recovered from untreated and cefazolin-treated (50 mg/mL, 100X MIC) biofilm-coated titanium implants were comparable for ORI16_C02N (6.81 log10 CFU, p = 0.52) and ORI16_C02J (6.44 log10 CFU, p = 0.49) (Fig. 5b). Despite demonstrating effectiveness of the StaPhage cocktail in inhibition of planktonic growth of ORI16_C02J and ORI16_C02N, differences were observed in the anti-biofilm activity of StaPhage toward the two S. aureus strains. Bacterial numbers in ORI16_C02J biofilms exposed to the StaPhage cocktail in vitro or untreated were similar (p = 0.86). In contrast, a significant decrease (3.3-fold, p = 0.004) was observed in bacterial numbers remaining on ORI16_C02N-coated implants after exposure to the StaPhage cocktail compared with untreated implants. Microbiological data were confirmed with SEM images of untreated, cefazolin-treated, and StaPhage treated biofilms of ORI16_C02N on titanium implants (Fig. 5c–e), demonstrating reduction in the thickness and area covered by S. aureus biofilm after 48 hours for implants exposed to the StaPhage cocktail.
Discussion
Despite significant improvements in surgical techniques and advancements in biomaterials for orthopedics, infection remains a leading cause of revision of total knee and hip arthroplasty [4]. Methicillin-sensitive S. aureus is the organism most commonly associated with PJI, with skin colonization an established risk factor for infection after arthroplasty [3,13,15]. The global threat of antibiotic resistance has ignited interest in the use of bacteriophages as an alternate management option for biofilm-related infections, including PJI. We have conducted proof-of-principle in vitro studies demonstrating potential application of lytic bacteriophages in the management of S. aureus biofilms growing on porous titanium, a material commonly used in the manufacture of orthopedic implants.
Phages are highly abundant in the environment and are relatively inexpensive to isolate and purify [9]. Despite the use of phages for medical, veterinary, agricultural, and aquaculture application for almost a century, there have been no reported adverse safety concerns for mammalian cells [9,12,22]. Their specificity, safety for mammalian cells, and self-propagating characteristics are key characteristics for which phages are being increasingly being considered for the management of biofilm-related infections outside of Eastern Europe, where phage therapy has been used since 1921 [9].
The first report of clinical application of staphylococcal phages was for the successful management of skin infections [9]. Since then, staphylococcal phages have also been investigated for the management of bacteremia, eye, respiratory, gastrointestinal, urinary tract, and bone infections [23]. More than half of the individual phage preparations screened in the current study possessed lytic activity against the S. aureus strains recovered from arthroplasty patients. Five representative phages preparations were shown to prevent the planktonic, in vitro growth of MSSA and an MRSA strain.
The wide host spectrum of many of the individual phage preparations screened suggests their polyvalent nature and potential use in anti-staphylococcal cocktails. Notably, differences were observed in the spot test screening and the lytic activity of the phage preparations in broth cultures, highlighting the importance of the screening technique selected for determining the phage host range. These differences may reflect distinct mechanisms of action contributing to bacterial cell lysis. Plasticity in phage host range has also been demonstrated and may evolve over time or under different selective pressures, rather than being a fixed characteristic [24].
Often phage cocktails containing three or more individual phage preparations are used to widen the spectrum of anti-bacterial activity and the success rate of phage therapy for chronic bacterial infections [9,10,12]. In the current in vitro studies, we did not observe an additive effect on inhibition of planktonic S. aureus growth when individual phage preparations were combined as a cocktail. Although the most effective phage preparation differed for each S. aureus strain investigated, typically one of the five individual preparations tested exerted >90% reduction in bacterial growth when added alone at the time of bacterial seeding, suggesting an increased adsorption efficiency for the given strain of S. aureus. Inclusion of a “high-performing” preparation within a cocktail format would eliminate target bacterial cells rapidly, thus masking less obvious effects of additional phages on inhibition of planktonic S. aureus growth, in vitro. Nevertheless, the highly lytic impact of phages against planktonic growth of S. aureus strains was demonstrated with a 6 log decline of CFU within 4 hours of phage delivered at an MOI of 10:1. The reported optimal MOI for in vivo effectiveness of phages is between 1 and 10 [22], which is in accordance with our observations in the current study.
While the pathogenesis of PJI remains poorly understood, biofilm formation is considered pivotal [6,7]. Biofilm-forming capacity of S. aureus is thought to be influenced by clonal variation, rather than gene regulation, and is not influenced by antibiotic resistance [25,26]. Consistent with the literature [25], we observed differences in the capacity of S. aureus associated with skin carriage to form biofilms in vitro using a polystyrene microtiter assay. Porous titanium is used widely in orthopedic implants, and 3D-printing is being increasingly used for manufacture of patient-specific orthopedic implants [20]. To investigate the anti-biofilm activity of StaPhage against biofilms growing on clinically relevant orthopedic materials, we utilized 3D-printed porous titanium implants designed for a rat model of knee implant surgery.
Eradication of biofilms with conventional antibiotic therapy is extremely difficult, with MICs of 100 to 1000 times higher required for effectiveness against bacteria growing in biofilms than against bacteria in planktonic form [27], increasing the risk for collateral tissue damage. Data demonstrated that StaPhage more efficiently killed bacteria on the surface of implants than cefazolin, despite use of 100X MIC and despite demonstrating cefazolin sensitivity of these strains in planktonic form. The tolerance of MSSA biofilms to cefazolin is consistent with findings of Urish et al. [28], who demonstrated persister bacterial cells at supra-therapeutic levels of cefazolin, irrespective of biofilm depth.
Cefazolin is used widely for pre- and peri-operative prophylaxis in orthopedic surgical procedures and is the antibiotic agent of choice for management of post-operative infections caused by MSSA. Clinical failure of cefazolin has been described for high inoculum (>107 CFU) endocarditis and osteomyelitis infections, is associated with type-A and C beta-lactamase-producing MSSA strains, and is independent of clonal type [29,30]. A significant proportion of MSSA strains recovered from patients with osteomyelitis possesses the type A beta-lactamase variant, although cefazolin prophylaxis itself does not appear to exert a selective pressure for persistence of these MSSA strains in vivo [31,32].
While beta-lactamase typing of these strains was not performed, it is possible that the dramatic contrast in cefazolin sensitivity of planktonic and biofilm-associated ORI16_C02N and ORI16_C02J observed in the current study is a reflection of this high inoculum effect. Nevertheless, these data highlight the challenges of eradicating MSSA biofilms on orthopedic materials with cefazolin and warrant exploration of novel strategies to manage PJI.
Importantly, the StaPhage cocktail significantly lowered the bacterial biomass on titanium implants and the viability of biofilm-embedded bacteria, suggesting it also exerts lytic activity against slow-growing, persister cells. Reduced activity of the StaPhage cocktail was observed against ORI16_C02J compared with ORI16_C02N biofilms, highlighting S. aureus strain differences in susceptibility to StaPhage. This may reflect differences in phage infectivity rates, dependent on the mode of bacterial growth and presence and accessibility of phage adsorption receptors. The complexity of these interactions and the implications they have on therapeutic dosing of phages in terms of the tailoring of MSSA strain-specific phage cocktails, concentration administered, frequency of dosing, and route of administration highlight the importance of clinically relevant in vivo studies to more accurately evaluate efficacy of this approach [33].
Emergence of bacterial resistance and host immunogenicity has been suggested as potential limitations for phage therapy. Importantly, phage efficacy is independent of the antibiotic resistance phenotype of the target bacterial strain [12]. While there is potential for bacteria to develop resistance to phage through modification or shielding of cell surface receptors, these responses are often detrimental to bacterial virulence and survival in vivo [12].
Administration of phages has been shown to stimulate host immune responses, with the strength and type of response influenced by the route of administration and the dosing protocol used [9]. Oral and topical administration may induce the production of anti-phage neutralizing antibodies, while intravenous administration of phages has potential to stimulate innate and adaptive immune responses [9]. Nevertheless, safety has been demonstrated in patients with immunosuppression and autoimmune disorders [22] and indeed, indirect stimulation of cell-mediated immune responses after administration of phage may further facilitate bacterial clearance in vivo [9].
To minimize the potential for phage resistance and host immunogenicity, phage therapy centers in Eastern Europe deliver patient-tailored therapy within a controlled environment, use customized phage cocktails containing three or more individual preparations to broaden target specificity, and adopt short-term dosing regimes to avoid extended phage exposure and thus selective pressure for phage-tolerant bacterial strains [11]. Regulatory framework for the therapeutic application of phages in countries outside of Eastern Europe is yet to be established [11].
Discordance in the classification of phages underpins continued debate as to whether their use in medicine should adhere to current stringent legislative frameworks in place for pharmaceuticals or whether frameworks should be adapted to enable inclusion of customized, patient-specific phage preparations that are not marketed, but are distributed in a controlled way through approved national treatment centers [11]. Currently, application for administration of phages can be sought for compassionate use for patients with life-threatening bacterial infections. There is growing impetus, however, for the approval of phages for wider medical application with clinical trials currently under way to evaluate S. aureus and Pseudomonas aeruginosa phages to manage burn, rhinosinusitis, and cystic fibrosis-associated infections [12].
Given the well-documented disparity between antimicrobial efficacy using in vitro and in vivo systems [34], an extensive comparison of StaPhage with additional antibiotic agents and antibiotic combinations was not conducted in the current study. Substantive in vivo work using a clinically relevant animal model of PJI will be essential to characterize efficacy, the potential for development of bacterial and host resistance, and safety before consideration of clinical application. These complexities cannot be understood using in vitro studies.
Each phage preparation used in the current study was distinct as determined by a unique host range susceptibility pattern using spot testing of serially diluted lysates. While sequencing of each lysate has not been performed, formal genetic characterization of the phage preparations would form an essential component of the safety evaluation before progression to clinical application. An additional limitation of the current in vitro study was the small number of S. aureus strains screened for sensitivity to the StaPhage cocktail. The purpose of the current study, however, was to demonstrate feasibility of customizing phage cocktails to a given strain, rather than create a one-size-fits-all therapy.
Conclusion
The current proof-of-principle studies demonstrated antibacterial activity of phages against planktonic and biofilm-associated S. aureus strains growing on porous titanium scaffolds. Compared with cefazolin treatment, bacterial numbers were significantly reduced within S. aureus biofilms growing on titanium surfaces after exposure to a phage cocktail, in vitro. These preliminary data provide support for in vivo studies to further investigate the potential use of custom phage cocktails for management of orthopedic implant-related infections caused by S. aureus.
Footnotes
Acknowledgment
This research was supported by internal funds of the Orthopaedic Research Institute of Queensland and James Cook University.
Author Disclosure Statement
Titanium implants used in this study were provided in-kind by Stryker Orthopaedics. Bacteriophage preparations used in this study were provided in-kind by AusPhage Pty Ltd. LE is the Director of AusPhage Pty Ltd. LE was not directly involved in the conduct of experiments, data acquisition, or data analysis. The Orthopaedic Research Institute of Queensland (JM, AG, KH, MW, PM) receives annual funding donations from ARGO to support the offer of a surgical Research Fellowship, and from Stryker and Arthrex to support the salary of a Research Coordinator and Orthopaedic PHO position.