
Abstract
Abstract
Antibiotic resistance has been demonstrated during the entire duration of antibiotic use even before medical utilization. Increasing resistance within virtually all microbes continues to be a problem. Infection with antibiotic resistant microbes has demonstrated significantly increased morbidity, death, and health-care–associated costs. Given increasing antibiotic resistance, multiple novel agents and approaches are being investigated, including antimicrobial lipids. Sphingosine and ceramide have been demonstrated to play a pivotal role in the innate immunity of the epidermis, oral mucosa, and respiratory epithelium; their role is being investigated currently in uroepithelium. Ceramide has been shown to be pivotal in the regulation of mammalian defense against Pseudomonas aeruginosa and Staphylococcus aureus pathogens commonly encountered in pneumonia. On the other hand, sphingosine appears to be equally pivotal and directly involved in pathogenic defense and has been demonstrated to “rescue” mammals from P. aeruginosa infections. Within this review, we will discuss the role of sphingolipids within innate immunity, pathogen invasion, and bacterial infection. We will discuss the antimicrobial activity of sphingosine and possibility for commercial use as an antimicrobial in the post-antibiotic era.
A
While the above example focuses on only S. aureus, multiple drug-resistant (MDR) strains of Pseudomonas aeruginosa, Enterococcus faecium, Klebsiella pneumonia, and other pathogenic bacterium have also been identified [8–10,14]. Antibiotic resistance can occur from chromosomal mutation, expression of a latent gene after exposure, or through the exchange of genetic material via conjugation or transduction [16]. Bacterial resistance occurs because of destruction or inactivation of the antibiotic agent, alteration of the target site of the antibiotic agent, or efflux of the antibiotic agent [15,16]. Antibiotic-resistant bacterial infections are associated with higher number of deaths, longer hospitalizations, and higher cost in both gram-positive and gram-negative organisms [17–20]. It is estimated that nosocomial infections will develop in 2 million persons per year, with approximately 50%–70% of infections being MDR strains. Of those infected with MDR strains, approximately 77,000–90,000 patients will die each year [21]. From 1999–2005, hospitalizations because of methicillin-resistant S. aureus has more than doubled from approximately 127,000 to 278,000 patients [22].
Given increasing antibiotic resistance, multiple novel agents and approaches are being investigated. Promising novel approaches include the development of antimicrobial peptides, new vaccines, antibacterial bacteriophages, antisense antimicrobial agents, and antimicrobial lipids [23–28]. In this review, we will describe sphingosine and its analogues in innate immunity, bacterial infections, and as a potential therapeutic option to combat bacterial infections in the post-antibiotic era.
Sphingolipids Structure and Regulation
Sphingolipids are a class of lipids ubiquitous in eukaryotic cells that were believed initially to be structural components of cell membranes but have been found to regulate multiple cell functions via signal transduction [29,30]. In the 1880s, J. L. W. Thudichum first bestowed the “sphingosine” backbone of sphingolipids name because of its difficult to characterize (i.e., sphinx-like) properties [31]. The function and mechanisms of the sphingolipid class remained elusive until the 1980s when sphingosine was discovered to inhibit protein kinase C [32]. Subsequently, sphingolipids have been implicated in the induction of apoptosis, immunity, motility, proliferation, growth, differentiation, and infection [29–31].
Sphingolipids are composed of a long chain hydrocarbon backbone, with sphingosine, sphinganine, and phyosphingosine being the most common (Fig. 1). All sphingolipids have a fatty acid attached to carbon-2 and a head group attached to the hydroxyl group of carbon-1. More than 300 sphingolipids are known today with distinct head groups [30,31,33]. The hydrophobic head group and hydrophobic backbone are responsible for the amphipathic nature of sphingolipids and make the molecule unable to diffuse passively across the leaflets of the lipid bilayer. Sphingomyelin is the most abundant sphingolipid molecule in mammalian cells and forms microdomains within the plasma membrane. These microdomains are held in place via hydrophobic van der Waal interactions with cholesterol [34,35].
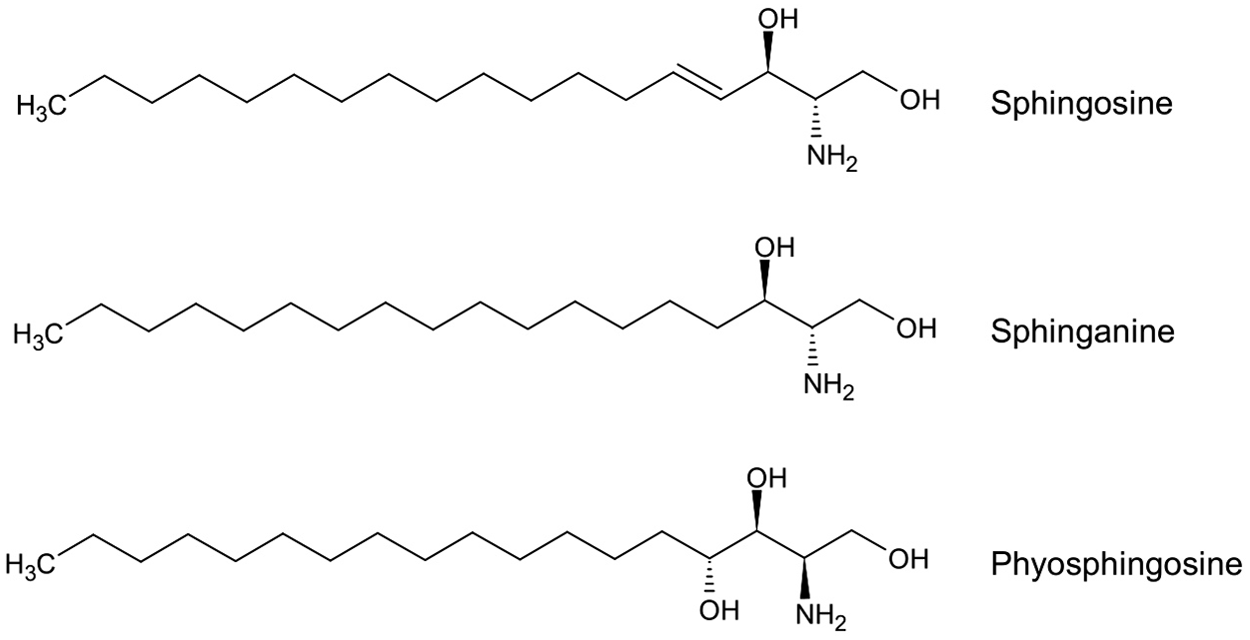
The structure of the most common backbones used in sphingolipid synthesis: Sphingosine, sphinganine, and phytosphingosine.
The synthesis of sphingolipids starts in the endoplasmic reticulum (ER) but transitions to the Golgi apparatus on differing sides of the plasma membrane of these organelles [33]. Via a series of reactions, sphinganine is synthesized from serine and palmitoyl-CoA [36]. Sphinganine subsequently will undergo further enzymatic reactions to form ceramide. Once ceramide is synthesized, multiple reactions can occur to create sphingomyelin, glucosylceramide and its derivatives, sphingosine, and more complex sphingolipids (Fig. 2) [31,33,37]. Within the sphingolipid family, ceramide and sphingosine-1-phosphate (S1P) are two of the best-studied sphingolipids. Ceramide has been demonstrated to promote apoptosis and arrest cell growth, while S1P has been implicated in the prevention of ceramide-induced apoptosis and stimulates cell proliferation [38,39]. The play between ceramide and S1P has led to the phrase “sphingosine rheostat” and demonstrates the fluid nature and opposing effects of ceramide/S1P balance [37,40]. Increased ceramide concentrations have been observed in conditions associated with increased cellular stress: Ischemia, radiation, inflammation with the release of pro-inflammatory mediators, and oxidative stress, to name a few [36,41].

Simple overview of sphingolipid metabolism and sphingolipids most observed in response to a bacterial pathogen.
An important enzyme to note is sphingomyelinase, which converts sphingomyelin to ceramide. There are several sphingomyelinase isoenzymes including the lysosomal acid-sphingomyelinase (ASM) and the secreted ASM, which have their highest enzyme activity at acidic pH [42,43]. The lysosomal and secreted ASM regulate sphingolipid turnover and degradation. When ASM is activated via stressors, a release of ceramide occurs within lysosomes and on the outer leaflet of the plasma membrane with the consecutive formation of small ceramide-enriched membrane domains that spontaneously fuse into ceramide-enriched membrane platforms and become signaling units [34,35,44]. Ceramide-enriched membrane platforms cluster receptor molecules on the plasma membrane and allow for signal transduction [44–46]. Evidence has pointed to the development of a “signalsome,” which not only facilitates the transmission of signals but also amplifies them by greater than 100-fold [47].
Formation of ceramide-enriched membrane platforms has been demonstrated after stimulation by CD95, CD40, tumor necrosis factor (TNF), interleukin (IL)1, platelet activating factor; infection with Neisseria gonorrhoeae, P. aeruginosa, Neisseria meningitides, S. aureus, Escherichia coli, and pathogenic mycobacteria; and UV-light irradiation, gamma-ray irradiation, and treatment with cisplatin [45,47–62].
Sphingolipids in Innate Immunity
Sphingolipids have been implicated in the function and signaling of the innate immune system. Receptors to S1P are present on dendritic cells, eosinophils, macrophages, mast cells, and NK cells. Binding of S1P to its receptor effects chemotaxis, differentiation, and effector cell response [63]. Within immune cells, sphingolipids and sphingolipid metabolism derivatives act as second messengers. Sphingolipids also function as ligands to bind to G protein-coupled receptors when released from activated immune cells [64,65].
Ceramide
Hinkovska-Glavheva et al. [66,67] demonstrated that ceramide modulates the phagocytic function of polymorphonuclear leukocytes (PMN). Phagocytosis mediated by Fcγ receptor binding induced plasma membrane-associated neutral sphingomyelinase, which subsequently increased ceramide formation [66]. Within the PMN, ceramide may also be metabolized to C1P, which activates phospholipase A2 [67]. Given these findings, the sphingomyelin-ceramide balance may exert a regulatory effect on phagocytosis in PMNs. Ceramide has also demonstrated involvement in the oxidative properties of PMNs; oxidative burst is inhibited. Ceramide appears to inhibit phospholipase D; in turn, phosphatidic acid is decreased and unable to activate nicotinamide adenine dinucleotide phosphate (NADPH) oxidase [32,68–70].
Apoptosis is necessary to resolve inflammation, and delayed apoptosis has been demonstrated in autoimmune disorders [71]. Intrinsic and extrinsic pathways exist for apoptosis control, although the downstream products of both pathways converge on varying caspases. The extrinsic pathway is initiated by various death receptors (Fas, TNF-related apoptosis-inducing ligand receptor, and TNF receptors) [71,72]. During the initial apoptotic events, sphingomyelin is cleaved via ASM-mediated hydrolysis, and ceramide is generated. Ceramide-enriched membrane platforms form and cluster signaling molecules required for apoptosis [37–41,72]. Ceramide accumulation has also been demonstrated to form channels within the mitochondrial membrane. These channels are large enough to release pro-apoptotic proteins, such as cytochrome c, from the intermembrane space of the mitochondria and allow apoptosis to proceed [73,74]. Bcl, an antiapoptotic protein, can disassemble these ceramide channels. This appears to be a highly regulated process [75].
Jaffrezou et al. [76] demonstrated a role for ceramide within the nucleus during apoptosis. Using isolated nuclei, ionizing radiation induced an accumulation of ceramide, neutral sphingomyelinase, and features of apoptosis [77]. Taken together, there are many possible mechanisms of sphingolipid induced apoptosis.
Lipopolysaccharide (LPS) is a major surface membrane component of most gram-negative bacteria and acts as an endotoxin [78]. It stimulates macrophages and triggers release of TNFα, IL-1, IL-6, and other signaling molecules [64] and induces macrophages to produce nitric oxide (NO), prostaglandin E2, and cyclo-oxygenase-2 (COX-2) via the activator protein-1 (AP-1) and nuclear factor κB (NFκB) pathways [79–82]. Hsu et al. [81] demonstrated that ceramide inhibited the response of macrophages to LPS by reducing the activation of AP-1 and NFκB.
Dendritic cells have receptors for CD40, TNFα, and IL-1, and stimulation results in ceramide accumulation as dendritic cells are induced to differentiate. Exogenous ceramide causes dendritic cells to down-modulate antigen uptake capabilities and ultimately reduces antigen presentation [83].
Within the innate immunity, natural killer (NK) cells are the primary cytotoxic cells, and activation has been demonstrated by direct contact with target cells. NK cells kill target cells by the Fas/CD95 receptors or cytoplasmic granule toxins after contact is made to induce apoptosis [84]. Activation of the Fas antigen results in an increase in ceramide levels, sphingomyelinase activity, and overall cytotoxicity. Ceramide-enriched membrane platforms occur resulting in the propagation of the apoptotic signal [45,47,85,86].
Ceramide accumulation has been shown to stimulated NADPH oxidase, xanthine oxidase, and NO synthase within endothelial cells, smooth muscle cells, and macrophages. This activation in turn, stimulates the release of reactive oxygen species (ROS) and the superoxide radical [87].
Sphingosine
Studies have demonstrated that the oxidative properties of PMNs are also affected by sphingosine. Sphingosine inhibits protein kinase C, which disrupts the NADPH oxidase pathway and thus the production of oxidative burst products [32,88,89].
Sphingosine-1-phosphate
Sphingosine-1-phosphate has been implicated in leukocyte adhesion and migration. Increased levels of intercellular adhesion molecule 1 (ICAM-1) and IL-8 were observed in human lung epithelial tissue after stimulation with S1P [90]. S1P lyase deficient mice demonstrated decreased recruitment of PMNs in response to inflamed tissues. The chemotaxic IL-23/IL-17/granulocyte-colony stimulating factor pathway appeared to be disturbed, resulting in the poor PMN trafficking [90]. Schwab et al. [91] administered 2-acetyl-4-tetrahydroxybutylimidazole to induce lymphopenia in mice and demonstrated greater than 100-fold increase in S1P in lymphoid tissue and inhibition of S1P lyase activity. Vogel et al. [92] demonstrated that S1P lyase (the enzyme that degrades S1P) knockout mice displayed lymphopenia, myeloid cell hyperplasia, and T-cell sequestration in lymph tissue. Heterozygous mice for S1P lyase demonstrated reduced lymphopenia and myeloid cell hyperplasia, but T-cell development/trafficking remained blunted. The above studies demonstrated the S1P is critical in the development and trafficking of immune cells.
Triggering of the FcɛRI receptor on mast cells results in sphingosine kinase activation, and sphingosine is metabolized to S1P. The S1P in turn activates various signaling molecules that lead to the release of TNFα and IL-5 transcription [93,94]. S1P is also exported out of the cell and exhibits autocrine transactivation of S1P receptors, which in turn leads to mast cell degranulation and chemotactic motility [95].
Sphingosine-1-phosphate has been demonstrated to mediate endocytosis and migration in mature dendritic cells, but does not appear to effect antigen processing [96].
In addition to receptor-mediated effects of S1P, sphingosine kinase also produces intracellular S1P after stimulation with a variety of growth factors and likely acts intracellularly [97]. These intracellular targets and mechanisms remain elusive to investigators, and further research is necessary. Intracellular S1P has been demonstrated to induce calcium release from the endoplasmic reticulum and is an integral messenger in the TNF-α, protein-kinase C, and prohibin 2 signal transduction pathways [98].
While ceramide demonstrates pro-apoptotic properties, S1P often antagonizes ceramide and promotes survival and growth.
Sphingolipids in Bacterial Infections
Grassmé et al. [99] were the first to identify a link between ASM and ceramide in bacterial infections. Neisseria gonorrhoeae, a common pathogen that causes a sexually transmitted disease, invades mucosal epithelial cells and fibroblasts by interaction of opacity-associated proteins with heparin sulfate proteoglycan (HSPG) [99,100]. By binding to HSPG receptors, ASM is activated. This activation releases ceramide to act as second messengers. N. gonorrhoeae bacteria were unable to invade cells that had undergone pharmacologic inhibition of ASM or ASM deficient cells. With restoration of ASM activity in deficient cells via plasmid transfection, the pathogenicity of N. gonorrhoeae was restored [99]. Additional studies have demonstrated other receptors—carcinoembryonic antigen-related cellular adhesion molecule receptors (CEACAM) and scavenger receptor expressed on endothelial cells (SREC-1)—are sites for the invasion of N. gonorrhoeae and use the sphingomyelinase-ceramide pathway [101,102]. Similar pathways were demonstrated with N. meningitidis infection [57].
Another important bacterium that relies on the ASM-ceramide pathway is S. aureus. After infection of human endothelial cells with S. aureus, ASM was activated, ceramide was released, and apoptosis occurred [103]. Apoptosis occurred via the activation of cellular caspases, release of cytochrome c, and the stimulation of Jun-NH2-terminal kinase. In ASM deficient cells or on pharmacologic inhibition of ASM, S. aureus triggered apoptosis did not occur, again demonstrating the underlying importance of the ASM-ceramide pathway [103]. Alpha toxin, a virulent pore-forming toxin secreted by S. aureus, activated ASM and released ceramide [104]. Endothelial cells incubated with alpha-toxin demonstrated increased ASM activity and ceramide production; the same relationship was seen when cells were inoculated with S. aureus. When endothelial cells were inoculated with alpha-toxin deficient S. aureus bacterium, ASM activity was unchanged [104]. The ASM-ceramide system has been implicated in the destruction of tight junction proteins in endothelial cells after S. aureus infection [105]. Endothelial cells demonstrated the destruction of tight junctions after incubation with alpha-toxin alone; when treated with amitriptyline, an inhibitor of ASM, tight junction morphology was preserved [104].
While E. coli is found in normal bacterial flora, opportunistic pathogenic strains exist and cause a variety of infectious diseases. Like many gram-negative bacteria, E. coli contains and releases LPS, which stimulates the ASM-ceramide pathway [64,78–82,106]. When dendritic cells were exposed to E. coli, apoptosis occurred; high concentrations of endogenously administered LPS mimicked this effect in immature dendritic cells. Apoptosis was because of the activation the ASM-ceramide pathway in dendritic cells [62].
The ASM-ceramide pathway has been extensively studied in the setting of P. aeruginosa bacterium. Grassmé et al. [107] demonstrated that the CD95/CD95 ligand were essential for cellular apoptosis during P. aeruginosa infection. It was further demonstrated that the creation of a ceramide-enriched membrane platform was required for the CD95/CD95 ligand pathway to function [47]. The creation of ceramide-enriched membrane platforms was also demonstrated to be essential for internalization of P. aeruginosa, possibly via cystic fibrosis (CF) transmembrane conductance regulator (CFTR). When ASM was inhibited, a massive inflammatory response, release of IL-1, and ultimately septic death occurred in mice after inoculation with P. aeruginosa [55]. Alveolar macrophages demonstrate activation of ASM, subsequent ceramide production, and development of ceramide-enriched membrane platforms after inoculation with P. aeruginosa. The membrane platforms are necessary for the activity of NADPH-oxidase, subsequent generation of reactive oxygen species, and elimination of bacteria, and if the infection persists, activation of JNK with subsequent apoptosis [108–111].
Many other pathogens have demonstrated reliance on the ASM-ceramide pathway within mammalian cells. Table 1 summarizes the different sites and roles of ceramide [47,55,57,62,99–111,136].
Different Roles of Ceramide after Bacterial Infection
Sphingolipids and Innate Defense of the Skin
The epidermis produces free fatty acids that have been implicated in cutaneous host defense since the 1940s [112]. Accumulation of lipids in lamellar granules occurs with development and differentiation of epidermal cells. Once maturing cells reach the boundary of the stratum corneum and granular layer, the lamellar granules are exocytosed, releasing phospholipids, cholesterol, and glycolipids. Hydrolytic enzymes within this layer produce ceramide from sphingomyelin and glucosylceramides from phosphoglycolipids [113]. Ceramidases are also present and break down ceramide into free sphingoid bases. These sphingoid bases are often sphingosines, dihydrosphingosines, 6-hydroxysphingosines, and their backbone ranges from 16–20 carbons in length [113–116]. Miller et al. [117] demonstrated that free fatty acids provided the most antimicrobial activity against Staphylococcus compared with polar lipids and glycosphingolipids. When essential fatty acid deficient (EFAD) mice were exposed to S. aureus, a large inoculum could be used; isolated lipids from EFAD mice proved more lethal against multiple bacterial [118]. Further examination of lipid composition demonstrated less triglycerides and more free fatty acids in the EFAD deficient mice. After the lipids of EFAD mice were fractioned out by thin-layer chromatography, glycosphingolipid and phospholipids demonstrated highly effective killing activity against S. aureus [117].
In 1992, Bibel et al. [119] first began further understanding the antimicrobial effects of sphingosine in the skin. Lipid screening demonstrated a profound effect of sphingosines (ranging from 0.19–50 mcg/mL lipid) against S. aureus in vitro compared with ethanol. The antimicrobial effects of sphingosines were consistent in in vitro assays against Propionibacterium acnes, Streptococcus pygoenes, Brevibacterium epidermidis, Micrococcus luteus, E. coli, Serratia marcescens, and P. aeruginosa. A significant progressive reduction in bacteria was demonstrated with increasing amounts of sphingosines in all assays. When sphinganine was applied to living skin and subsequently inoculated by various bacteria, a significant reduction (0.94–3.5 log at 200 mcg.cm2) in bacteria was demonstrated [119].
Further, Fischer et al. [120] determined the minimum inhibitory concentrations (MIC) of various lipids: Sphingoid bases—D-sphingosine, phytosphingosine, dihydrosphingosine; and fatty acids: sapienic acid, and lauric acid—on various bacteria. All sphingoid bases were active against E. coli, Fusobacterium nucleatum, S. aureus, Streptococcus sanguinis, Streptococcus mitis, Cornebacterium bovis, Cornebacterium striatum, and Cornebacterium jeikeium (MIC 0.5–15.6 units) Fatty acids, on the other hand, demonstrated worse antimicrobial activity; lauric acid was antimicrobial against F. nucleatum, S. sanguinis, and S. mitis, (MIC 2.1-15.6) while sapienic acid was antimicrobial against F. nucleatum (MIC 6.5 units) Interestingly, neither sphingoid bases nor fatty acids demonstrated antimicrobial effects against P. aeruginosa and S. marcescens (MIC >500 units) Sphingosine and phytosphingosine killed S. aureus, S. sanguinis, S. mitis, and C. striatum within 0.5–6 hours. While the mechanism of antimicrobial activity is not yet fully understood, antimicrobial lipids may disrupt the cell wall, alter the cytoplasmic membrane, or penetrate the cytoplasmic membrane and alter the cytoplasm of the bacterium [120].
Atopic dermatitis (AD) is a chronic inflammatory disease associated with dry, scaly patches of skin; patients exhibit an increased susceptibility to bacterial colonization as well. Patients with AD demonstrate an alkalization of their epidermis [121]. Bibel et al. [119] demonstrated that the ideal environment for sphingosine's antimicrobial properties is in an acidic environment with rapid loss of activity at a pH of 8. Patients with AD demonstrate marked reduction of ceramide and sphingosine within the stratum corneum. A reduction in the activity of acid ceramidase was observed. Increased bacterial susceptibility is likely because of the reduction of ceramide, sphingosine, and acid ceramidase confounding one another [120,121]. Murata et al. [122] discovered sphingomyelin deacylase within the stratum corneum. Over-expression of sphingomyelin deacylase is present in patients with AD [123], likely contributing to the difference in ceramide and sphingosine compared with healthy controls. Sphingomyelin deacylase catalyzes the conversion of sphingomyelin to sphingosylphosphorylcholine [122]. The higher level of sphingomyelin deacylase likely competitively competes with sphingomyelinase for sphingomyelin and thus leads to decrease ceramide and sphingosine [123].
Sphingolipids and Innate Defense of the Oral Cavity
The oral mucosa and vermillion border of the lip contain sebaceous glands and follicles without terminal hairs. Like the skin, sebaceous glands secrete sebaceous lipids [124]. It has been reported previously that neutral lipids, phospholipids, and glycolipids are present within human saliva [125]. The majority of human saliva is made up of neutral lipids, with 1%–5% of the total lipid mass from phospholipids and glycolipids [126]. After further analysis, the neutral lipids that make up saliva include cholesterol, fatty acids, triglycerides, cholesterol esters, wax esters, and squalene; the composition closely resembles that of the lipids secreted by sebaceous glands at the skin [123]. Brasser et al. [127] demonstrated that free sphingosine was also present in saliva.
With evidence of sphingosine and free fatty acids in saliva and oral mucosa, antimicrobial properties are likely present. In the oral cavity, S. mitis, S sanguinis, and F. nucleatum are often present [128]. Sphingoid bases and fatty acids demonstrated antibacterial properties in the 2011 study published by Fischer et al. [120]. The antimicrobial effects of prior sphingoid bases and fatty acids [120] were found to be present against Porphyromonas gingivalis, a common pathogen implicated in periodontitis [129]. To further mechanistically evaluate the antimicrobial effects, electron microscopy was performed after sapienic acid and phytosphingosine incubation with P. gingivalis. Disruption of the bacterial plasma membrane and ultrastructural damage were observed after infection. Sapienic acid treated P. gingivalis demonstrated unregulated proteins involved in glycolysis, amino acid metabolic processes, microbial metabolism, polypeptide acquisition and degradation, adhesion, and virulence [129].
Sphingolipids and Innate Defense of the Lungs
Pathogenesis of P. aeruginosa
The respiratory surface is a critical barrier against pathogens, similar to skin function. The respiratory tract is exposed to innumerable bacteria, particles, and toxins daily while exchanging mass quantities of oxygen and carbon dioxide. Respiratory epithelial cells line the alveoli and bronchi, providing a framework for its barrier function. Mechanisms of protection include complex tight adhesions between cells, mucocilliary clearance, fluid and mucin secretion, antimicrobial peptides, and recruitment/retention of immune cells [130]. Secreted antimicrobial peptides include lysozymes, beta-defensins, lactoferrin, and surfactant proteins A and D [131]. Within the milieu of cells, alveolar macrophages, lymphocytes, and neutrophils play an important role in the defense of the respiratory system [130].
Within the respiratory tract, P. aeruginosa is a common opportunistic pathogen and can produce crippling morbidity and death [132]. Infection with P. aeruginosa leads to the release of pro-inflammatory cytokines (IL-1, IL-6, and IL-8), internalization of the bacterium, and ultimately apoptosis of the cell. [55]. In patients with CF, P. aeruginosa is an important pathogen; patients with CF demonstrate increased susceptibility to P. aeruginosa infections [133]. The role of sphingosine and the ASM-ceramide pathway within the respiratory tract has been extensively studied during P. aeruginosa pathogenic invasion.
Previous research demonstrated that CD95/CD95 ligand interactions were crucial for the immune response to P. aeruginosa and concomitant apoptosis of infected cells. With neutralization of the CD95/CD95 ligand system in lpr or gld mice, P. aeruginosa caused profound sepsis and death within 48 hours [107]. CD95 had been demonstrated previously to activate ASM and subsequently cause ceramide accumulation on stimulation. During CD95 stimulation, rapid clustering of receptors was demonstrated; in ASM deficient cells, clustering did not occur. In cells with a deficiency of acid ceramidase (metabolizes ceramide), CD95 clustering was enhanced [134]. Further observation demonstrated that genetic deficiency of ASM negated the apoptotic effects of CD95 binding, but with exogenous ceramide application, apoptosis was restored. These results demonstrate that the ASM-ceramide clustering is crucial for the apoptotic function of CD95 [134]. Further, by blocking formation of the membrane platforms, P. aeruginosa was no longer able to induce apoptosis, and there was a greater than eight-fold increase in IL-1β production.
Further studies demonstrated that infection with P. aeruginosa stimulated ASM containing intracellular vesicles to fuse with the extracellular leaflet and co-localize to the membrane platforms propagating ceramide production [56]. Internalization of P. aeruginosa seems to induce NFκB nuclear translocation, which is ultimately responsible for apoptosis [135]. A functional ASM is also required to activate NADPH oxidase and to allow the formation of reactive oxygen species that kill bacteria, as discussed above. In addition, ASM-activity and ceramide are certainly required to form sphingosine, which function is discussed further below,
It is assumed that the beneficial effect of cell death against the infection, which seems to be counterintuitive, is mediated by an exfoliation of infected cells into the lumen. Death of the cells releases the intracellular pathogen and allows the immune system to attack the pathogen. In addition, apoptotic cells might be simply transported out of the body by the mucociliary clearance. In CF mice, a reduction of mucociliary clearance, deposition of deoxyribonucleic cid (DNA) fragments, and increased adherence of P. aeruginosa within the respiratory mucosa was observed [136,137].
Given patients with CF have increased susceptibility to P. aeruginosa, much research has focused on what mechanisms differ from healthy persons. Patients with CF have a mutation in the CFTR transmembrane protein resulting in failure of the protein to reach the plasma membrane. The chronic airway inflammation and susceptibility to pathogens that patients with CF experience is incompletely understood. [136]. Pier et al. [138] demonstrated that CFTR is necessary for P. aeruginosa binding, internalization, and clearance. Using murine epithelial cells, wild-type cells demonstrated significantly enhanced ingestion of P. aeruginosa compared with cells lacking CFTR or CFTR deficient cells transfected with cDNA of ΔF508 CFTR, the product of a known genetic mutation in patients with CF. Pier et al. [138] further went on to identify that the first extracellular domain of CFTR acts as the cellular receptor for the binding and internalization of P. aeruginosa. With pharmacologic blocking of this domain, P. aeruginosa was unable to be internalized in epithelial cells. [139]
Further identification of raft proteins involved in internalization of P. aeruginosa includes Src-like kinases, Lyn, and major vault protein (MVP) [140–142]. When Lyn, an Src-like kinase, was inhibited, internalization of P. aeruginosa, the release of cytokines, and apoptosis was inhibited. Lyn activation activated phosphatidyl-inositol-3-kinase and subsequently Akt to regulate internalization of P. aeruginosa and stimulate respiratory oxidation [140,141]. MVP is recruited to ceramide rafts on P. aeruginosa binding to CFTR, and deficiency leads to poor internalization of the bacteria [142]. The direct function of these proteins in is yet to be defined in ceramide-enriched membrane platforms, and further research is ongoing.
Sphingosine as a preventative and treatment adjunct
Sphingosine was first described as a possible preventative measure during P. aeruginosa lung infection in 2014 [143]. Initial experiments demonstrated sphingosine was abundant in human nasal epithelial cells of healthy individuals but was virtually absent in nasal cells isolated from patients with CF. The same relationship of sphingosine content was demonstrated in tracheal and bronchial epithelial cells between wild type and CF mice or ceramide synthase deficient (CerS2) mice. Overall, CF mice appeared to have a 75% reduction in sphingosine levels compared with wild type mice with a corresponding accumulation of ceramide. Application of acid ceramidase to the surface of the isolated CF tracheal epithelial cells normalized sphingosine levels via ceramide hydrolysis. In CF mice, inhalation of acid ceramidase or sphingosine increased sphingosine levels in the tracheal epithelial cells [143]. The CF and Cer2S mice demonstrated significantly increased response to P. aeruginosa inoculation with increased bacterial counts, signs of pneumonia and sepsis, and massive cytokine release. When CF and Cer2S mice were exposed to inhaled sphingosine or FTY720, a sphingosine analog, 1 hour before P. aeruginosa inoculation, a protective effect was observed. Sphingosine and FTY720 were also administered 1 hour after P. aeruginosa inoculation and completely blocked infection in CF and Cer2S mice [143].
Further research at the University of Cincinnati has demonstrated that burn-injured and aged mice have similar reductions in sphingosine as observed in CF mice. This imbalance of sphingosine results in increased susceptibility and death to P. aeruginosa in both cohorts [144,145]. When burn-injured mice were pre-treated with sphingosine or neutral ceramidase before bacterial inoculation, bacterial levels were significantly reduced and death rates improved. The IL-6 and IL-10 levels also returned to baseline after treatment with sphingosine or natural ceramidase in burned mice [144]. In aged mice, treatment with inhaled sphingosine 30 minutes before P. aeruginosa inoculation demonstrated significantly decreased levels of bacteria within bronchoalveolar lavage and lung tissue specimens compared with untreated aged mice; bacterial counts were the same in the young mice cohort. Sphingosine did not seem to enhance Pseudomonas clearance in young, healthy mice [145].
Tabazavareh et al. [146] took the findings of CF and P. aeruginosa infections discussed above to further investigate the relationship between CF and S. aureus. In vitro incubation of S. aureus with sphingosine induced rapid death of the bacterium while incubation with ceramide had no antimicrobial effect. Further, inhalation of sphingosine 30–40 minutes before inoculation of CF mice with S. aureus prevented infection. These findings, combined with those of P. aeruginosa, establish the antimicrobial role of sphingosine and the possible future use in clinical practice to prevent and treat pneumonia in the CF and critically ill population.
Martin et al. [147] investigated the delivery and toxicity of increasing sphingosine doses to healthy mice; overall, no adverse effects were demonstrated. First, lipid colloid suspensions with sphingosine and attached fluorofluor demonstrated good penetration in the trachea and bronchus. The inhalation delivery method appears to work and deliver sphingosine to the intended target. Second, DNA fragmentation in bronchial epithelium, pulmonary neutrophil recruitment, and morphologic changes in bronchial epithelium were not demonstrated after sphingosine administration. This shows that sphingosine is overall a safe and non-toxic molecule.
Seitz et al. [148] has developed a novel method to coat endotracheal tubes with sphingosine, again evolving the delivery method of sphingosine. Sphingosine and D-ribo-phytosphingosine, a sphingosine analog, coated endotracheal tubes significantly reduced bacterial adherence of S. aureus, P. aeruginosa, and Acinetobacter baumannii after in vitro incubation. When compared with commercially available silver-coated endotracheal tubes, sphingosine- and phytosphingosine-coated endotracheal tubes were superior against S. aureus. This may provide future therapeutic use in the prevention of ventilator-associated pneumonia.
Sphingolipids and Staphylococcus Bacteremia
The ASM-ceramide pathway has also been implicated in S. aureus sepsis and infection. Severe pulmonary edema often develops in patients with severe sepsis from infection with S. aureus; this has also been shown to occur in wild type mice after infection. The ASM deficient mice injected with intravenous S. aureus demonstrated reduced lung edema compared with wild type mice. Using Evans blue dye, a marker of endothelial cell integrity, wild type mice demonstrated marked leakage of the dye into the lungs while ASM deficient mice displayed minimal leakage. Further, hematoxylin and eosin staining demonstrated thickened hyaline walls and cellular infiltrates in the wild type mice but virtually normal morphology in the ASM deficient mice [105]. Murine endothelial cells infected with S. aureus demonstrated increased ASM activation and subsequent ceramide formation. Indirect immunofluorescence of endothelial cells demonstrated marked degradation of tight junction proteins; inhibition of ASM via amitriptyline mitigated this effect. The destruction of tight junction proteins by S. aureus is a result of ASM activation and directly contributes to the pulmonary edema observed in S. aureus sepsis [105]. Becker et al. [104] took the previous study a step further and demonstrated that alpha toxin alone was responsible for the severe lung edema and marked degradation of tight junctions via the ASM-ceramide activation.
The alpha-toxin produced by S. aureus also activates the ASM-ceramide pathway in macrophages. This activation triggers the release of IL-1β and TNF-α; in ASM deficient macrophages and ASM- inhibited (via amitriptyline) macrophages, cytokine release was reduced. [149] ASM-ceramide plays a pivotal role in S. aureus pathogenesis and virulence within a murine model.
Sphingolipids and Innate Defense of the Bladder
Research is currently under way at The University of Cincinnati looking at the role sphingolipids may play within bladder epithelium and infection. As outlined above, research has demonstrated that sphingosine plays a role in innate immunity during infection in the skin, oral cavity, and respiratory epithelium. Human uroepithelial cells from patients with recurrent urinary tract infections demonstrated decreased sphingosine, increased ceramide, and decreased acid ceramidase content compared with uninfected patients [150]. Unpublished research demonstrates that sphingosine was present at the uroepithelium in wild type mice and that this accumulation was ceramidase dependent. Wild type mice were then inoculated with E. coli, and sphingosine was administered via catheter into the bladder. A significant reduction in bacterial content was demonstrated in sphingosine treated mice.
Future Directions
This review demonstrates the elusive nature of sphingolipids in bacterial infections; there is an important balance between sphingolipid molecules within signaling. Ceramide and ASM are necessary for the pathogenesis of bacteria and also the host response to bacteria. While physiologic concentrations of ceramide are required for NADPH oxidase activation and ROS release, as well as controlled release of cytokines, constitutively increased levels of ceramide concentrations—as in CF or after burn—are harmful and sensitize the host to infection with P. aeruginosa and S. aureus. This sensitization is thought to be because of increased adhesion, internalization, spontaneously increased cell death, release of DNA into the bronchial lumen, and reduction of mucociliary clearance. A lack of ceramide is also harmful, because it prevents the release of reactive oxygen intermediates, apoptosis, and cytokine release. Thus, cellular levels of ceramide must be tightly regulated to allow for an efficient immune response.
Endogenous sphingosine has been demonstrated to have antimicrobial properties in the skin, oral cavity, respiratory tract, and urinary tract. The most exciting and promising effects of endogenous sphingosine administration can be observed with inhaled sphingosine and pulmonary infection. Sphingosine has been shown to be non-toxic in mammals at varying concentrations. Further research needs to continue to assess and develop treatment modalities for sphingosine in pulmonary, skin, mucosal, and urinary tract infections. The future is bright for sphingolipids in antimicrobial applications in the post-antibiotic era.
Footnotes
Author Disclosure Statement
No competing financial interests exist.