
Abstract
Background:
Acute intestinal ischemia–reperfusion injury (AIIRI) is a devastating clinical condition relevant to multiple diseases processes, including sepsis, trauma, transplantation, and burns. An AIIRI is a contributor to the development of multiple organ dysfunction syndrome (MODS). Oncostatin M (OSM)/oncostatin M receptor (OSMR) signaling is an unrecognized and novel candidate pathway for the mediation of MODS. In this study, we hypothesized that OSM mediates the injury mechanism of AIIRI leading to MODS.
Methods:
Wild-type (WT) and OSMR-knockout (OSMR-/-) C57BL/6 mice underwent AIIRI using a well-established model of selective occlusion of the superior mesenteric artery (SMA). Serum cytokine concentrations were measured using a multiplex detection system. Further tissue analysis was conducted with polymerase chain reaction, enzyme-linked immunosorbent assay, Western blots, and histologic review.
Results:
Survival was significantly higher in WT than in OSMR-/- groups at 30 minutes of ischemia with 2 hours of reperfusion (100% versus 42.9%; P = 0.015). No significant differences in the degree of local intestinal injury was seen in the two groups. In contrast, the degree of lung injury, as evidenced by myeloperixodase activity, was lower in OSMR-/- animals in the early AIIRI groups. There was a greater degree of renal dysfunction in OSMR-/- mice. Oncostatin M mediated interleukin (IL)-10 upregulation, with WT animals having significantly lower IL-10 concentrations (52.04 ± 23.06 pg/mL versus 324.37 ± 140.35 pg/mL; P = 0.046).
Conclusion:
Oncostatin M signalling is essential during acute intestinal ischemia–reperfusion injury. An OSMR deficiency results in decreased early lung injury but increased renal dysfunction. There was a significantly increased mortality rate after AIIRI in mice with OSMR deficiency. Augmentation of OSM may be a novel immunomodulatory strategy for AIIRI.
Multiple organ dysfunction syndrome (MODS) is a major contributor to morbidity and death in patient populations worldwide. The syndrome is relevant to many disease processes, such as sepsis and trauma [1].
The gastrointestinal tract is an important defense barrier for the body, but in critically ill patients, it can become a seriously pathogenic source of bacteria and inflammatory mediators leading to a septic response such that it is a critical contributor to MODS [2-4]. Acute intestinal ischemia–reperfusion injury (AIIRI) is an adaptive response to shock; the mortality rate ranges from 60%–90%, a value that has not changed since the 1940s. This high mortality rate is attributable not only to local organ injury but also to the systemic inflammatory response and distant organ injury [5, 6].
The pathogenesis of MODS remains poorly understood but is related to a common pathway that leads to the dissemination of systemic mediators that result in progressive distant organ injury [7, 8]. These mediators are regulated further by local mediators that are activated by local ischemia or injury. Many mediators of MODS have been examined, such as interleukin (IL)-6 [9], inducible nitric oxide synthase [10, 11], and reactive oxygen species [12].
Oncostatin M (OSM)/oncostatin M receptor (OSMR) signaling is an unrecognized and novel candidate pathway for the systemic inflammatory response and mediation of MODS. The protein belongs to the IL-6 family of cytokines (IL-6, IL-11, IL-27, leukemia inhibitory factor [LIF], OSM, ciliary neurotrophic factor [CNTF], cardiotrophin [CT]-1, cardiotrophin-like cytokine [CLC]) that all signal through GP130 [13-17]. Oncostatin M was first investigated for its role in inhibiting the growth of A375 melanoma cells [18]. The protein has since been found to be a multi-functional cytokine with roles in hematopoiesis [19], modulation of cell growth [20,21], remodelling of the extracellular matrix [22], and mediation of the acute-phase response in both the liver and kidney [23,24]. Oncostatin M has been demonstrated to regulate the production of pro-inflammatory cytokines such as IL-6 [25]. It is produced by several cell types, including polymorphonuclear leukocytes and monocytes, which are innately involved in the response to cellular injury [26, 27].
Investigation of the role of OSM in MODS has been scant to date. Limited clinical studies have shown that the concentration of OSM is upregulated in patients with septic shock [28]. The OSM gene was identified as one of the genes differentially upregulated in early sepsis compared with “sterile systemic inflammatory response syndrome” (SIRS) [29]. The functional aspect of OSM signalling in AIIRI or MODS has not been explored to a significant extent. In view of the functional roles that have shown OSM to have an important systemic inflammatory role, OSM may play a significant part in inflammatory signalling in SIRS and MODS.
In this study, we hypothesized that OSM mediates the mechanism of AIIRI leading to MODS. Using OSM receptor-knockout (OSMR-/-) mice, we examined the functional role of OSM in a superior mesenteric artery (SMA) occlusion model of AIIRI.
Material and Methods
Animals
Male wild-type (WT) C57BL/6 mice were obtained from Charles River Laboratories (Ottawa, Ontario, Canada). The OSMR homozygous knockout (OSMR-/-) mice were obtained from the line of B6.129S-Osmr<tm1Mtan> heterozygotes, developed by Tanaka et al. and bred by Charles River Laboratories [19]. All animals were housed in the Health Sciences Laboratory Animal Services (HSLAS) facility at the University of Alberta in conventional housing rooms. They were kept in pathogen-free conditions with 12:12-hour light:dark cycles and fed ad libitum on a standard murine diet and tap water. All animal experiments were reviewed and approved by the Animal Care and Use Committee of the University of Alberta.
Intestinal ischemia: Reperfusion model
The SMA model of ischemia–reperfusion injury has been well-described in the literature. Animals were anesthetized using intra-peritoneal injections of ketamine (120 mg/kg) and xylazine (16 mg/kg). An appropriate depth of anesthesia was maintained with regular intra-peritoneal injections of those drugs throughout the procedure.
A midline laparotomy was created, and bowel was lateralized to expose the mesenteric vessels. The SMA was isolated, and a 10-g micro-vascular clamp was applied to induce ischemia for 30 or 60 minutes. The clamp was removed to allow reperfusion for either 2 or 4 hours. In the sham-treatment group, laparotomy, exposure, and dissection of the SMA were performed as in the experimental groups, with no ischemia or reperfusion period.
The WT and OSMR-/- animals were divided into four experimental groups: sham treatment (n = 4, each genotype), 30 minutes of ischemia with 2 hours of reperfusion (n = 8, each genotype), 30 minutes of ischemia followed by 4 hours of reperfusion (n = 7, each genotype), and 60 minutes of ischemia with 2 hours of reperfusion (n = 10 for WT, n = 14 for OSMR-/-). Animals were euthanized at the end of the reperfusion period at 2 or 4 hours except in the event of death prior to the endpoint.
Myeloperoxidase activity assay
Tissue specimens (40 mg) were homogenized and processed in hexadecyltrimethylammonium bromide (HTAB) buffer using a previously described protocol [30,31]. Samples were sonicated for 10 seconds, followed by freezing in liquid nitrogen and thawing. Samples then were centrifuged, with the cycle repeated three times with collection of pooled supernatant liquid. Protein concentrations were determined in duplicate using the Bradford assay with Coomassie Plus reagent (Thermo Fisher Scientific, Waltham, MA, USA). Myeloperoxidase (MPO) activity was measured by the H2O2-mediated oxidation of the substrate o-dianisidine hydrochloride. The MPO reaction was carried out for 30 minutes and examined using a SpectraMax plate reader (Molecular Devices, Sunnyvale, CA, USA) at 460 nm. Activity was normalized to milligrams of protein.
Plasma cytokine measurement
Blood samples were obtained from all mice at the end of reperfusion. Blood from the inferior vena cava was collected in heparinized syringes and centrifuged to collect plasma. Undiluted plasma samples were used for measurements of all cytokines. Plasma concentrations of OSM were measured by an enzyme-linked immunosorbent assay (R&D Systems, Minneapolis, MN). Additional cytokines were measured using a multiplex detection system for IL-1β, IL-6, IL-10, interferon (IFN)-γ, and tumor necrosis factor (TNF)-α (Bio-Rad Laboratories, Hercules, CA). Samples were measured using a Luminex Magpix system (Luminex Corporation, Austin, TX) according to the manufacturer's protocol.
Histologic examination
Tissue samples were fixed in buffered 10% Formalin solution at room temperature for at least one week. Fixed tissues were processed and embedded in Paraplast (Thermo Fisher). Embedded tissues were sectioned at a thickness of 3 to 5 mcm. Slides were stained using a routine hematoxylin and eosin (H&E) stain and reviewed by a pathologist in a blinded fashion. The Park-Chiu score was used to grade the degree of intestinal injury [32,33]. Histologic lung injury was evaluated using a method described by Slutsky et al. [34]. In brief, this method assesses multiple histologic features, including alveolar distention, alveolar collapse, peri-vascular hemorrhage, alveolar hemorrhage, peri-vascular edema, polymorphonuclear leukocyte infiltration, alveolar membranes, and alveolar edema. Scoring for each feature is from 0 to 3 (0 = normal, 1 = mild, 2 = moderate, 3 = severe). Aggregate scores were reported out of a maximum of 24.
Statistical analysis
Quantitative data are expressed as mean ± standard error. Statistical analysis was performed by ANOVA followed by the Tukey-Kramer test for post-hoc pairwise comparisons. Non-parametric analysis for histologic injury scores was performed using the Kruskal-Wallis test, followed by the Tukey-Kramer method for ranked pairwise comparisons. Statistical significance denotes a p value <0.05. Statistical analysis was performed using the STATA 12 (StataCorp, College Station, TX, USA) statistical software package.
Results
OSMR deficiency results in differential mortality rate compared with wild-type in intestinal ischemia: Reperfusion injury
The superior mesenteric artery (SMA) occlusion model to induce AIIRI is a well-established model that results in time-dependent local and distant organ injury [31]. In our study, groups of 30 minutes/2 hours, 30 minutes/4 hours, and 60 minutes/2 hours were used (nomenclature: ischemia–reperfusion time). The model allows comparison of organ injury at the designated reperfusion endpoints (i.e., 2 hours and 4 hours). Although this was not intended as a survival study, there was a notable differential survival outcome in the WT and OSMR-/- groups in reaching the reperfusion endpoint in that it decreased with increasing ischemia and reperfusion times (Fig. 1). In the WT animals, survival ranged from 100% in sham-treated groups to 60.0% in the 60 minutes/2 hours group. For OSMR-/- mice, survival ranged from 100% for sham treatment to 28.9% in that group. There was no significant difference between genotypes at the shortest ischemia and reperfusion time (30 min/2 h). In the 30 minute/4 hours group, the survival of WT and OSMR-/- mice was 100% and 42.9%, respectively (P = 0.015). The survival rates in the 60 minutes/2 hours group were 60.0% and 28.6% for WT and OSMR-/-, respectively, which was near statistical significance (P = 0.058). Death occurred during the reperfusion phase in all animals except for one mouse in the WT 60 minutes/2 hours group, which died during the ischemia phase.
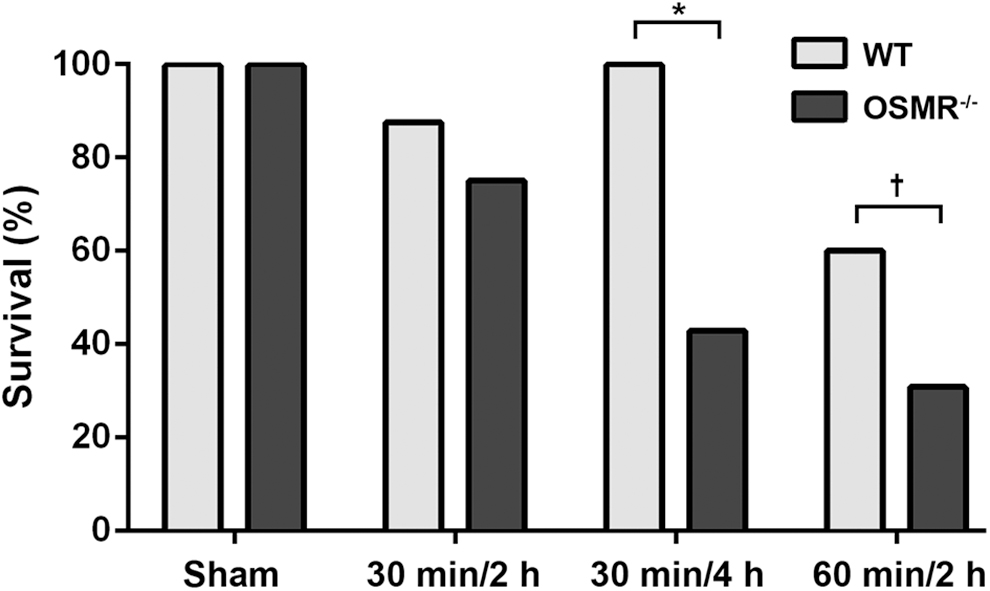
Survival outcome in wild-type (WT) and oncostatin M receptor -knockout (OSMR-/-) mice undergoing acute intestinal ischemia–reperfusion injury. Survival declined in a time-dependent fashion with increasing ischemia and reperfusion times. Survival rates diverged in the two groups. *P = 0.015; †P = 0.058.
For the remainder of the analysis, only animals reaching the pre-determined reperfusion endpoint (in each allocated group) were used for tissue, histology, and serum analysis. Exclusion of the non-surviving animals was done to avoid significant confounding from the various durations of the reperfusion phase.
Local intestinal injury after intestinal ischemia: Reperfusion injury
Intestinal injury increases in a time-dependent fashion with increasing ischemia and reperfusion times. Myeloperoxidase activity assay was used as a surrogate marker of injury and neutrophil infiltration, as there was time-dependent increase in MPO activity with increasing ischemia and reperfusion times (Fig. 2A). With WT mice, there was a 2.3-fold and 2.8-fold increase in MPO activity at 30 minutes/4 hours and 60 minutes/2 hours, respectively, compared with sham-treated animals, which was not statistically significant. In OSMR-/- animals, there were no significant changes in MPO concentrations at 30 minutes/2 hours, 30 minutes/4 hours, and 60 minutes/2 hours groups. There was a statistically significant difference in MPO concentrations in the 60 minutes/2 hours groups between WT and OSMR-/- mice (p = 0.044).
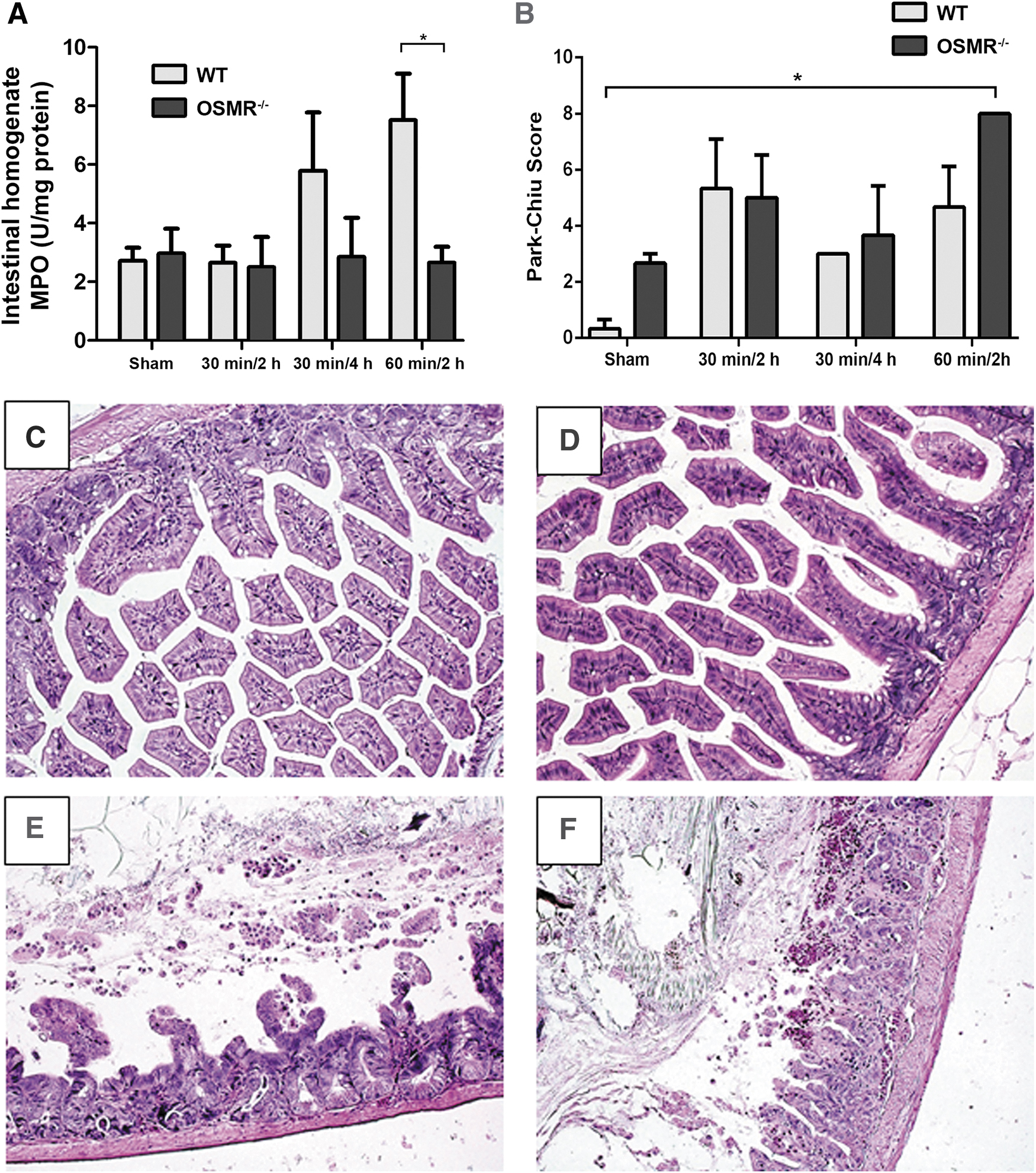
Degree of intestinal injury was ameliorated with intact oncostatin M (OSM) signalling. (
Increasing durations of ischemia and reperfusion time resulted in more evidence of histologic intestinal injury, which correlated with a higher tissue injury score that was most pronounced in the 60 minutes/2 hours group (p < 0.05). There were no statistically significant differences between WT and OSMR-/- within each AIIRI group (Fig. 2B). Interestingly, the difference in MPO activity in the 60 minutes/2 hours group was not demonstrated in histologic tissue injury scores. Increasing degrees of AIIRI were associated with histologic changes, including epithelial necrosis and sloughed villi (Fig. 2C).
Multiple organ dysfunction after AIIRI
Distant organ injury is a key component of the AIIRI model. Lung injury was used as one marker for distant organ involvement. The WT mice demonstrated increased MPO concentrations in lung tissue with ischemia–reperfusion (Fig. 3A). The largest degree of increase was seen in the 30 minutes/2 hours group, which was significantly higher in WT animals than in OSMR-/- mice (P < 0.05). In OSMR-/- mice, the peak in lung MPO occurred later, in the 30 minutes/4 hours group. This was confirmed with histologic evidence of lung injury with increasing severity of AIIRI reflected in lung tissue, seen with alveolar hemorrhage and inflammatory cell infiltrate (Fig. 3C). There was no statistically significant difference in histologic injury scores between WT and OSMR-/- mice (Fig. 3B).

Lung injury in response to intestinal ischemia–reperfusion injury. (
Renal impairment is often seen in MODS. Using the blood urea nitrogen concentration as a marker for renal dysfunction, AIIRI led to an increase in BUN in both WT and OSMR-/- genotypes (Fig. 4). The BUN concentration was higher in OSMR-/- than in WT in all pairs of experimental groups and in the sham-treated group (p = 0.017). Limited renal dysfunction was seen in the 30 minutes/2 hours groups in both WT and OSMR-/-. The largest magnitude of difference in BUN was seen in the 30 minutes/4 hours group, but this was not statistically significant.
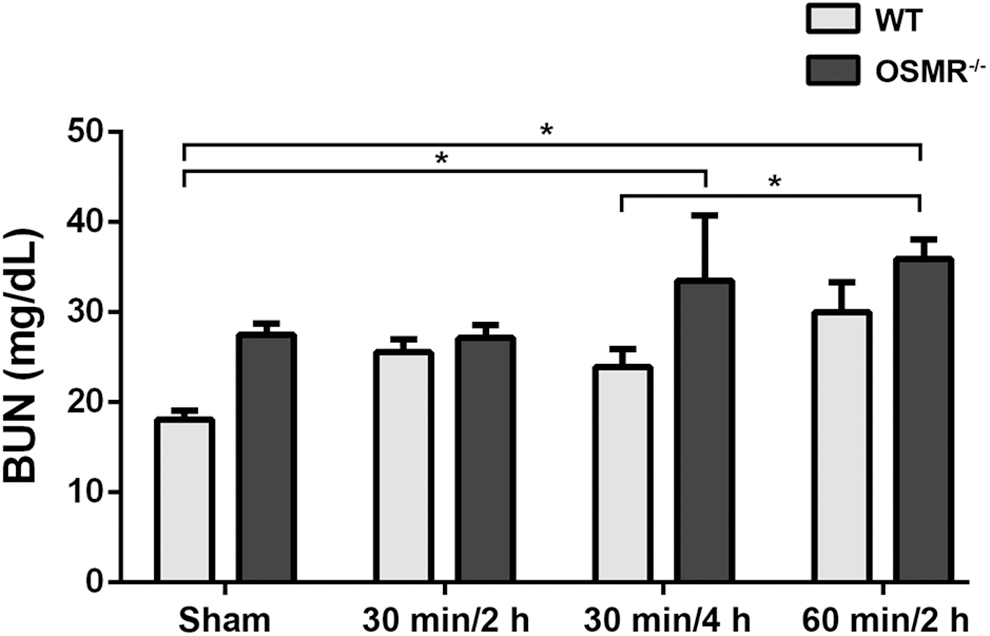
Blood urea nitrogen (BUN) concentrations after intestinal ischemia–reperfusion injury. Concentration was higher in all oncostatin M receptor-/- groups than in wild-type groups. *P < 0.05.
Plasma OSM concentrations in acute intestinal ischemia: Reperfusion injury
Plasma OSM concentrations were measured in all groups (Fig. 5). The OSM concentrations were lower in WT animals, with a peak at 30 minutes/4 hours. The plasma concentrations in OSMR-/- groups were higher, including peak values at 30 minutes/4 hours (94.91 ± 22.48 pg/mL versus 31.58 ± 15.42 pg/mL; p = 0.022). The largest difference in OSM concentrations was in the 30 minutes/4 hours group, which was the group with the greatest mortality rate divergence between the two genotypes.
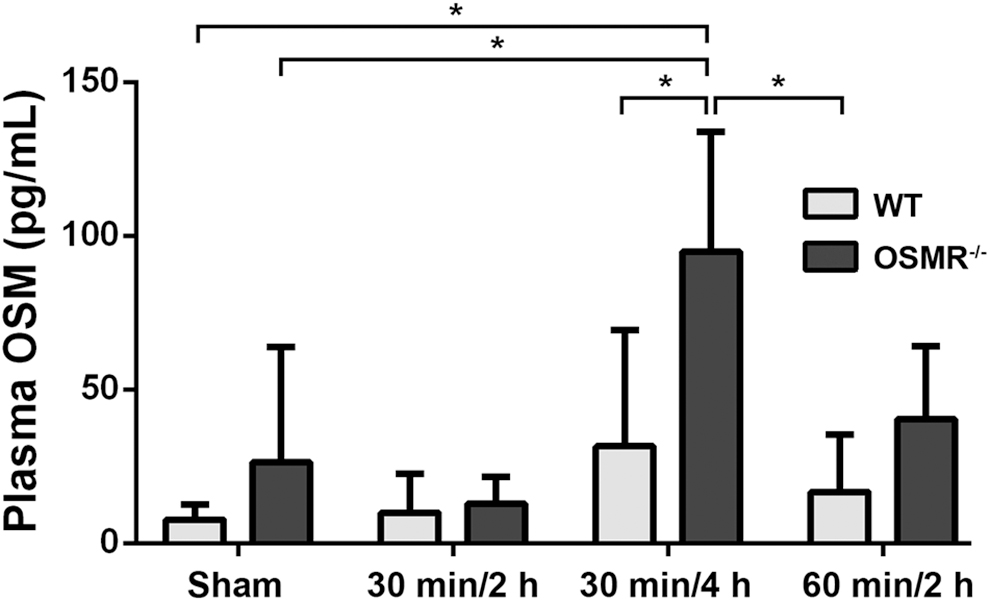
Plasma oncostatin M (OSM) concentrations in murine intestinal ischemia–reperfusion injury. Concentrations peaked at the 30 min/4 h in both wild-type (WT) and OSM receptor (OSMR)-/- groups. Plasma OSM concentrations were higher in OSMR-/- groups than in WT groups. *P < 0.05.
Reduced pro- and anti-inflammatory cytokine concentrations in OSMR-knockout mice
Systemic cytokine concentrations in plasma were measured in all groups (Table 1). Peak plasma concentrations depended on the specific cytokines measured. There was a statistically significant difference in IL-10 plasma concentration in the 30 minutes/4 hours group between WT and OSMR-/- (52.04 pg/mL versus 324.37 pg/mL; p = 0.046), which was the ischemia–reperfusion group that showed the largest divergence in mortality rate.
Plasma Cytokine Concentration (expressed in pg/mL) in Wild-Type (WT) and Oncostatin M Receptor (OSMR-/-)-Knockout Mice after Acute Intestinal Ischemia Reperfusion Injury
Wild type and oncostatin M receptor (OSMR) knockouts were subjected to various times of intestinal ischemia and reperfusion (groups are denoted by ischemia time/reperfusion time). Statistical significance is defined as P < 0.05.
Abbreviation: NS = not significant.
The peak plasma concentrations of each of these cytokines were variable, with the peak differences between groups occurring in the 30 minutes/2 hours, 60 minutes/2 hours, and 30 minutes/2 hours groups for TNF-α, IL-6, and IFN-γ, respectively. There were no significant differences in plasma concentrations of IL-1β and IL-17 between the WT and OSMR-/- animals. There was a large variability in peak plasma concentrations of TNF-α, IL-6, and IFN-γ in OSMR-/- groups, with a general up-regulation in all concentrations compared with sham-treated mice. Although there was a non-statistically significant trend to higher concentrations of TNF-α, IL-6, and IFN-γ in OSMR-/- groups, the large variability did not result in statistical significance.
Discussion
The investigation of the role of OSM in SIRS and MODS is a novel area of research. In this study, OSMR-/- animals were used to assess the functional deficiency of OSM signalling in AIIRI. The main findings of this study are that the OSM concentration rises early, and OSM deficiency results in more deaths. The OSMR-deficient animals had decreased lung injury at earlier time points but great renal impairment at longer ischemia–reperfusion times. The mechanism of these findings is related to increased anti-inflammatory cytokine concentrations after AIIRI in mice deficient in OSM signalling.
In humans, OSM signals through type I (gp130-LIFRβ heterodimer) and type II (gp130-OSMRβ heterodimer) receptors [35-37]. In mice, OSM signalling occurs only through the type II receptor, which is ablated with deficiency of OSMR in the knockout mice. In previous studies using OSMRβ knockout, the predominant differences from WT phenotypes were reduced peripheral erythrocyte and platelet counts [19]. In a separate study, OSMR-/- animals were found to have impaired hepatic repair after injury [38].
This is the first study to examine OSM signalling in AIIRI. Although there was a higher mortality rate with OSMR deficiency, the mortality difference was not fully explained by the differences in either local intestinal or distant organ injury. There was greater intestinal injury, as measured by MPO, in the WT 60 minutes/2 hours group compared with OSMR-/-. There was significantly greater renal injury, as represented by BUN concentration, in OSMR-/- groups at 30 minutes/4 hours and 60 minutes/2 hours compared with the WT animals.
The strong induction of OSM expression in OSMR-deficient groups with AIIRI suggests a role for OSM during the early time points of reperfusion. The survival divergence between WT and OSMR-/- animals was significant in the 30 minutes/4 hours groups, which was consistent with the highest peak in plasma OSM concentrations. Deficiency of the OSMR led to a stronger induction of OSM systemically, reflecting an appropriate negative feedback inhibition of OSM signaling in OSMR-/- animals. This effect is likely mediated by SOCS3, which is strongly induced by OSM, and strongly inhibits OSM signaling [16,39]. With the shorter ischemia time point of 30 minutes/2 hours, there was no difference in the mortality rate, with significantly lower lung MPO activity in OSMR-/- animals.
The lack of correlation between MPO activity and tissue injury with OSMR-/- corresponds to the literature establishing the role of OSM in neutrophil transmigration, activation, and adhesion [40]. Neutrophils are critical mediators of early tissue injury and release OSM early through degranulation of intracellular stores promoting local inflammation [26]. Also, OSM is abundantly available in other cell types, such as macrophages and T lymphocytes, which were not studied in this model [21]. As neutrophil infiltration throughout the intestinal wall is anticipated during AIIRI [41], the lack of MPO activity along with high expression of OSM in OSMR-/- implicates OSM in neutrophil recruitment in the early time points of reperfusion. The temporal nature of these findings illustrates that time-dependent inhibition of the OSMR axis may be beneficial in early intestinal ischemia-induced organ injury.
The limitation of the study is the exclusion of the non-surviving animals from tissue analysis. This may have created a survival bias that excluded the samples with the greatest derangement in inflammatory and tissue injury markers. Given the higher mortality rate in the OSMR-deficient group, this effect would dilute the observed differences between WT and OSMR-/- animals.
Survival differences correlated with upregulation of IL-10 responses in the OSMR-/- animals. These findings suggest a role for OSM in mediating inflammatory activation after AIIRI. Oncostatin M has pro-inflammatory and anti-inflammatory activities according to the cellular context [21]. In this study, OSM played a role in mediating the IL-10 response, which plays prominent inflammatory roles in the setting of AIIRI. Interleukin-10 is described classically as an anti-inflammatory cytokine involved in maintenance of intestinal barrier function [42]. However, in AIIRI, augmented concentrations of IL-10 have detrimental effects on local intestinal injury and distant organ dysfunction [43,44]. This may explain how the elevated IL-10 concentrations in the OSMR-deficient mice resulted in a higher mortality rate. Further study into additional signalling intermediates could explain the effects seen in this study. The progression of this model would be to study the impact of directed inhibition of the OSM/OSMR axis using a targeted antibody and to assess the longer-term impact of early targeting of this pathway in an animal model.
In conclusion, OSM is a cytokine induced early by AIIRI. The OSM/OSMR axis is an important mediator of organ injury, and deficiency results in a higher mortality rate after AIIRI. Elucidation of the novel OSM/OSMR pathway may suggest alternative approaches to preventing local and remote organ injury after AIIRI.
Footnotes
Acknowledgment
A special thanks to Mr. Christopher Lukowski for his technical expertise and to Dr. Saad Y. Salim for his assistance with the manuscript.
We thank the Edmonton Civic Employees (ECE) and the University (of Alberta) Hospital Foundation for all of their support in our research.
Author Disclosure Statement
The authors have no sources of funding to disclose.