
Abstract
Background:
Burn injury continues to be a significant cause of morbidity and death, with infectious complications being the primary cause of death. Patients are susceptible to overwhelming infection secondary to both the physical breakdown of the skin and mucosal barrier and the immune dysfunction that accompanies the inflammatory response to a major burn. With resistance to traditional antibiosis looming as a serious threat to patient outcome, advancement in the treatment of burn infections is imperative.
Methods:
Between February 15 and March 15, 2020, a search of Pubmed and clinicaltrials.gov was performed using search terms such as “burn immunotherapy,” “therapeutic microorganisms in burn,” “burn infection clinical trials,” and applicable variations.
Results:
Topical antimicrobial drugs continue to be standard of care for burn wound injuries, but personalized and molecular treatments that rely on immune manipulation of the host show great promise. We discuss novel therapeutics for the treatment of burn infection: Probiotics and therapeutic microorganisms, immune modulators, tailored monoclonal antibodies, and extracellular vesicles and proteins.
Conclusions:
The treatment strategies discussed employ manipulation of structure and function in host immune cells and pathogen virulence for improved outcomes in burn infection.
Burn trauma is a common cause of morbidity and death worldwide, with nearly 500,000 patients per year receiving treatment [1-3]. Survival rates have improved because of innovation in fluid management, wound care, early nutrition, and infection control. However, the primary cause of death in all age groups remains infection and subsequent sepsis. The burn wound is the most likely portal of entry for pathogens, although greater susceptibility to invasion from the lung, abdomen, and urinary tract have been demonstrated [4].
The skin and native microbiota are the first line of defense against environmental pathogens in the body. The skin provides an obvious physical barrier as well as a competent immune system. Indeed, local skin flora impacts the regulation and balance of the leukocyte types in the body [5]. Thus, disruption of this barrier through large surface area burns leaves the patient at risk for immune dysfunction. Burn patients also suffer from gut mucosal loss and endothelial integrity, which permits bacterial translocation and further bacterial exposure [6]. This breakdown in the traditional immune defense systems is compounded by a state of relative immune compromise that is observed in burn patients who survive. After the initial trauma, major findings include the depletion of blood lymphocytes, cytokine suppression, and decreased ability of cells to proliferate on stimulation. This is thought to leave patients at high risk for infection of the wound, lung, and urinary tract, as well as bacteremia and overwhelming sepsis [7,8]. A secondary infection can exacerbate immune suppression and dysfunction, as summarized in Figure 1.
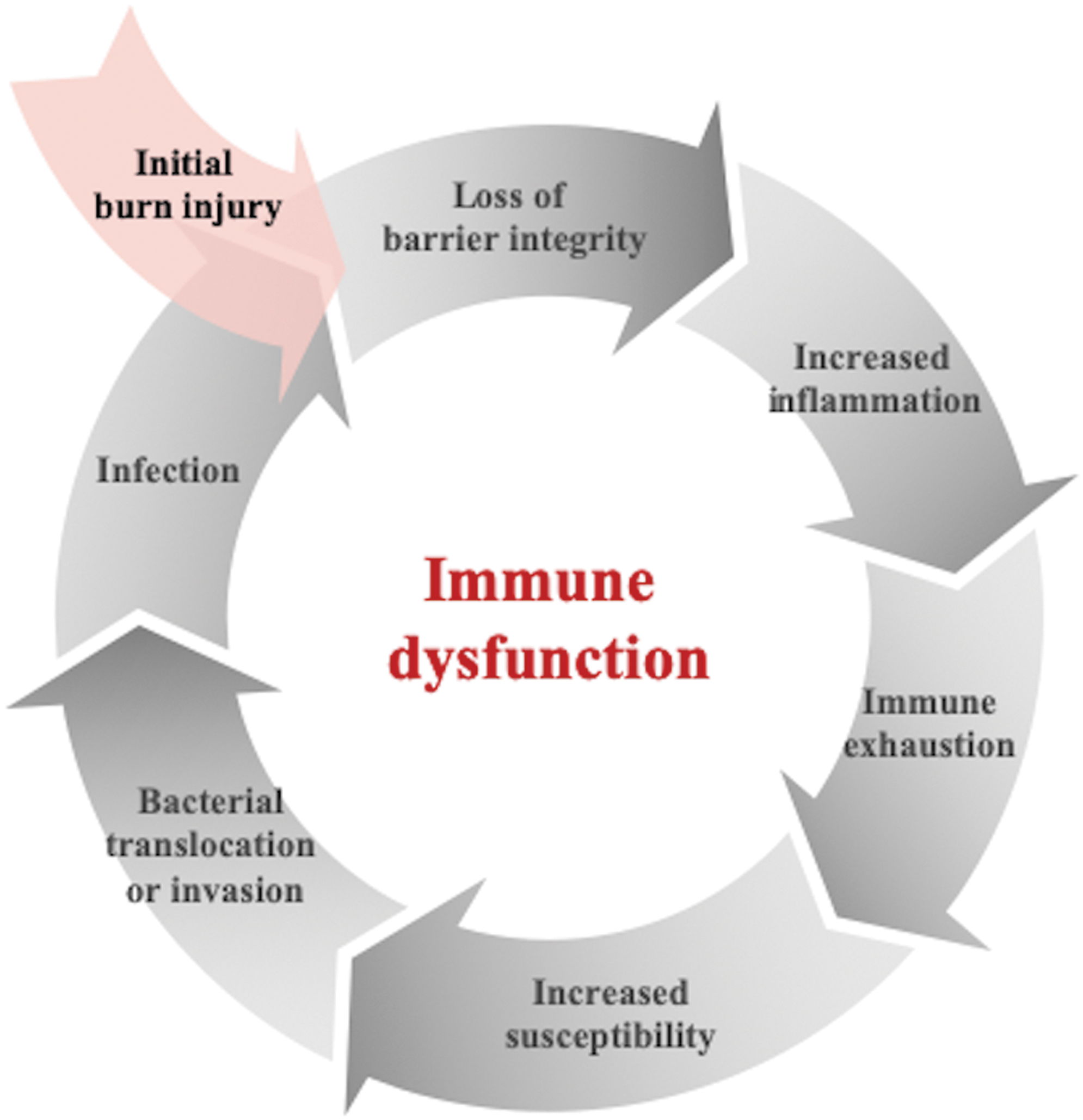
Burn-induced immune dysfunction. Burn injury causes loss of skin, mucosal, and endothelial integrity, as well as up-regulation and eventual dysfunction of the immune system. This leaves patients highly susceptible to infection, which further propagates the immune compromise.
Traditional therapy for burns has revolved around debridement of devitalized tissue, saline irrigation, and application of antimicrobial agents to reduce the bacterial load in the wound [9,10]. Antibiotics have been used both parenterally and locally, although topical use is emphasized, as systemic antibiotics may have poor penetrance to the site of infection in sufficiently active concentrations to eliminate microbial pathogens [1,11]. However, the common usage of antibiotics has led to the rise of antimicrobial drug-resistant pathogens, with an incidence of multi-drug–resistant infection between 10%–17%, which imposes a significant burden on the healthcare system [12]. Alternatives to traditional antibiotics in the treatment of burn infections are continually being sought, as diagnosis and treatment continue to evolve [11]. Here, we review recent improvements in treatment modalities in burn infection.
Therapeutic Micro-Organisms: Probiotics and Bacteriophages
Probiotics
The World Health Organization (WHO) defines probiotics as “live microorganisms, which when administered in adequate amounts, confer a health benefit on the host” [13]. There has been a compelling interest in the medical benefit of probiotics in the scientific literature, with evidence for their value in gastrointestinal, gynecologic, and dermatologic illness, as well as demonstrated effects on lowering blood pressure and cholesterol concentrations [14, 15]. Studies show that probiotics, most commonly Lactobacillus species, have a competitive inhibition effect on pathogenic bacteria while not inhibiting protective native strains, as well as a stimulating effect on the production of immune cells [16]. Multiple animal models have demonstrated the efficacy and safety of probiotic use. For example, application of Lactobacillus plantarum decreases the severity and length of murine burn wound infection with Pseudomonas aeruginosa [17]. Additional studies demonstrated a protective effect of topical Lactobacillus when applied in a rodent burn model prior to wound exposure to P. aeruginosa, Escherichia coli, and Staphylococcus aureus, although a definite therapeutic effect on an established infection was equivocal [18, 19].
Clinical studies have demonstrated that topical probiotic treatment of second-degree burns using L. plantarum was as effective as silver sulphadiazine in decreasing pathogen load [20]. At this point, higher-power clinical studies will be needed to demonstrate statistical significance. Altogether, the evidence for the safety and efficacy of topical probiotics as a valid alternative to traditional antimicrobial therapy is encouraging.
In addition to the topical usage of probiotics, there is an opportunity for the use of enteric organisms. After burn injury, functional gene expression of the microbiome is altered dramatically by probiotics. Specifically, 38 of 331 signaling pathways are changed significantly [21]. Further, butyrate-producing bacteria are reduced to an important degree in the intestine of burn-injured mice compared with healthy controls [22]. This is significant, as butyrate can regulate T-cell function and prevent T-cell apoptosis in an acid sphingomyelinase-dependent manner [23]. T-cell depletion is a key component of burn-induced immunosuppression and enhanced susceptibility to opportunistic infections. Additionally, fecal microbiota transplants from healthy donors can reconstitute butyrate-producing organism populations in burn-injured mice. Interestingly, this re-introduction of normal colonic flora ameliorates burn-induced colon permeability [22], decreasing the likelihood of enteral bacterial translocation and subsequent bacteremia. Given the apparent alterations in the gut microbiota after burn injury, enteral administration of butyrate-producing microorganisms may be useful as a therapeutic adjunct to ameliorate adaptive immunosuppression and intestinal integrity after a burn.
Oral administration of probiotics also has been of significant interest, given its ease of delivery and ready availability. Oral administration of Lactobacillus to human patients resulted in shorter durations of a requirement for antifungal agents after burn injury. However, it did not change the requirement for antibiotic agents [24]. Other studies have shown trends toward a decreased incidence of infection after administration of L. fermentum throughout the hospital stay [25]. In contrasting, a separate study demonstrated that when patients were given L. acidophilus and L. rhamnosus, there were no significant differences in the incidence of Clostridioides difficile infection, sepsis rate, emesis, or gastric residual volume. This absence of therapeutic benefit may in fact be associated with a higher incidence of diarrhea and malabsorption [26].
Thus, both exogenous topical application and oral administration of probiotic species have demonstrated a decreased pathogenic load in the wound. Further studies are needed to recommend routine oral use in burn patients.
Bacteriophage therapy
Bacteriophages are an alternative type of therapeutic microorganism that can be engineered to target pathogenic bacteria [27]. A bacteriophage is a natural bacterial virus able to enter into a host bacterium and replicate until the pathogen is destroyed [28].
Multiple studies have demonstrated phage therapy to be a safe alternative to current antibiosis of drug-resistant bacteria. Phage cocktails have improved survival and decreased bacterial counts in burn-injured, bacteria-inoculated mice compared with control animals [28]. Clinical trials have taken place with varied results. For example, PP1131, a cocktail of 12 natural phages, was compared with standard of care treatment, silver sulfadiazine, in human patients with infected burn wounds. The treatment reduced bacterial burden to the primary endpoint, but at a slower rate than was seen with the standard-of-care topical treatment [29].
Limitations of phage therapy include identification of appropriate delivery vehicles given the instability and rapid clearance of bacteriophages by the reticuloendothelial system in human beings. Additionally, adequate specificity for the host bacterium requires a cocktail of multiple targeted bacteriophages to have even somewhat broad-spectrum capacity, and further investigation into long-term effects after systemic application are needed [30,31].
Both of these applications of micro-organisms have shown promise in burn infection. Oral probiotics are used routinely in burn care, although clinical evidence of efficacy still is lacking. Phage therapy shows promise as an adjunct to current antimicrobial regimens, and specific cocktails are undergoing Phase I trials (clinicaltrials.gov; NCT 04323475).
Immune-Modulating Therapies
Immunomodulation can be an alternative antimicrobial strategy, specifically avoiding resistance to bacterial infections in patients predisposed to infectious complications. The ultimate goals of this approach are first to determine the specific immune dysfunction and then to improve the immune status of the host. This circumvents the increasing incidence of bacterial antibiotic resistance, especially in nosocomial infections.
Toll-like receptor modulation
Gram-negative bacteria express a lipopolysaccharide (LPS or endotoxin) in the cell wall that mediates an immune response in the host through interaction with Toll-like receptor 4 (TLR4), which is expressed on a variety of leukocytes [32]. Activation of TLR4 induces a pathway that leads to cellular production of pro-inflammatory mediators such as cytokines, chemokines, and nitric oxide that shape the immune response, namely leukocyte recruitment and vascular permeability [33]. After exposure to LPS, myeloid cells can develop tolerance to prevent further high degrees of inflammation. In murine models of bacterial infection, LPS tolerance has a protective effect; however, this therapy may be limited in human beings because of the potential toxicity [34].
The TLR4 agonist monophosphoryl lipid A (MPLA) has long been used successfully as an immune adjunct in human vaccinations [35]. This immune activation is theorized to be of clinical use in burn injury and infection. For example, burn-injured mice treated with MPLA demonstrate better survival, attenuated pro-inflammatory cytokine production, and enhanced clearance of bacteria in both the cecal-ligation and puncture models of sepsis and in mice with Pseudomonas wound infection [36]. More recently, in burn-injured sheep with pneumonia, MPLA treatment attenuated acute lung injury, improved pulmonary mechanics, and lowered inflammatory cytokine concentrations with significantly decreased vascular permeability [37]. Altogether, MPLA represents a promising therapeutic product for burn patients at risk for infection.
In contrast to therapies that target TLR4, which is found predominantly on myeloid cells, interleukin (IL-7) is a cytokine responsible for T-cell development, proliferation, and homeostasis. In human burn injury, IL-7 concentrations fluctuate throughout the course of injury and in response to burn severity but trend lower than in control patients. Additionally, in the same study, patients who survived a burn had higher IL-7 concentrations than non-survivors [38]. In murine models of sepsis, IL-7 treatment can decrease sepsis-induced depletion of CD4+ and CD8+ T cells and improve lymphocyte recruitment, preventing a sepsis-induced loss in adaptive immunity. Importantly, the application of IL-7 does not exacerbate the pro-inflammatory response in murine models of sepsis and improves survival [39]. As a test for feasibility in human patients, lymphocyte proliferation and function were improved to a similar extent after ex vivo IL-7 treatment of lymphocytes isolated from septic patients [40]. Finally, IL-7 was utilized in a first trial of an immunoadjuvant therapy targeting defects in adaptive immunity in patients with sepsis. The IL-7 treatment reversed T-cell depletion and was well tolerated [41]. Altogether, the use of recombinant IL-7 is a promising future therapy for burn-induced lymphocyte suppression [42].
Burn injury and wound infection consistently cause a decline in total white blood cell and lymphocyte populations, implying adaptive immune dysfunction [43]. Fms-like tyrosine kinase-3 ligand (Flt3L) is a hematopoietic factor ligand necessary for the expansion and maintenance of early hematopoietic stem cells. Additionally, it regulates expansion of dendritic cell populations and enhances neutrophil antimicrobial functions, rendering it an effective immunomodulatory agent [44]. Treatment with Flt3L in a murine burn injury model augments lymphocyte counts, increases T-cell interferon gamma production, and blunts programmed cell death ligand (PD-L1) expression by antigen-presenting cells, as will be discussed in the next section [45].
Burn injury can result in a profound derangement of the inflammatory response, leaving patients particularly vulnerable to infection. The LPS-induced tolerance can cause down-regulation of pro-inflammatory cytokines, while rhIL-7 and Flt3L both up-regulate other aspects of the immune response, as summarized in Figure 2. Multiple clinical trials are under way evaluating the efficacy of these immune therapies in sepsis, but as yet, they have not been used clinically in the treatment of burns. There are multiple immune targets under investigation currently, but more research is needed on immunologic staging of patients to better time therapeutic intervention and balance pro-inflammatory and anti-inflammatory treatments.
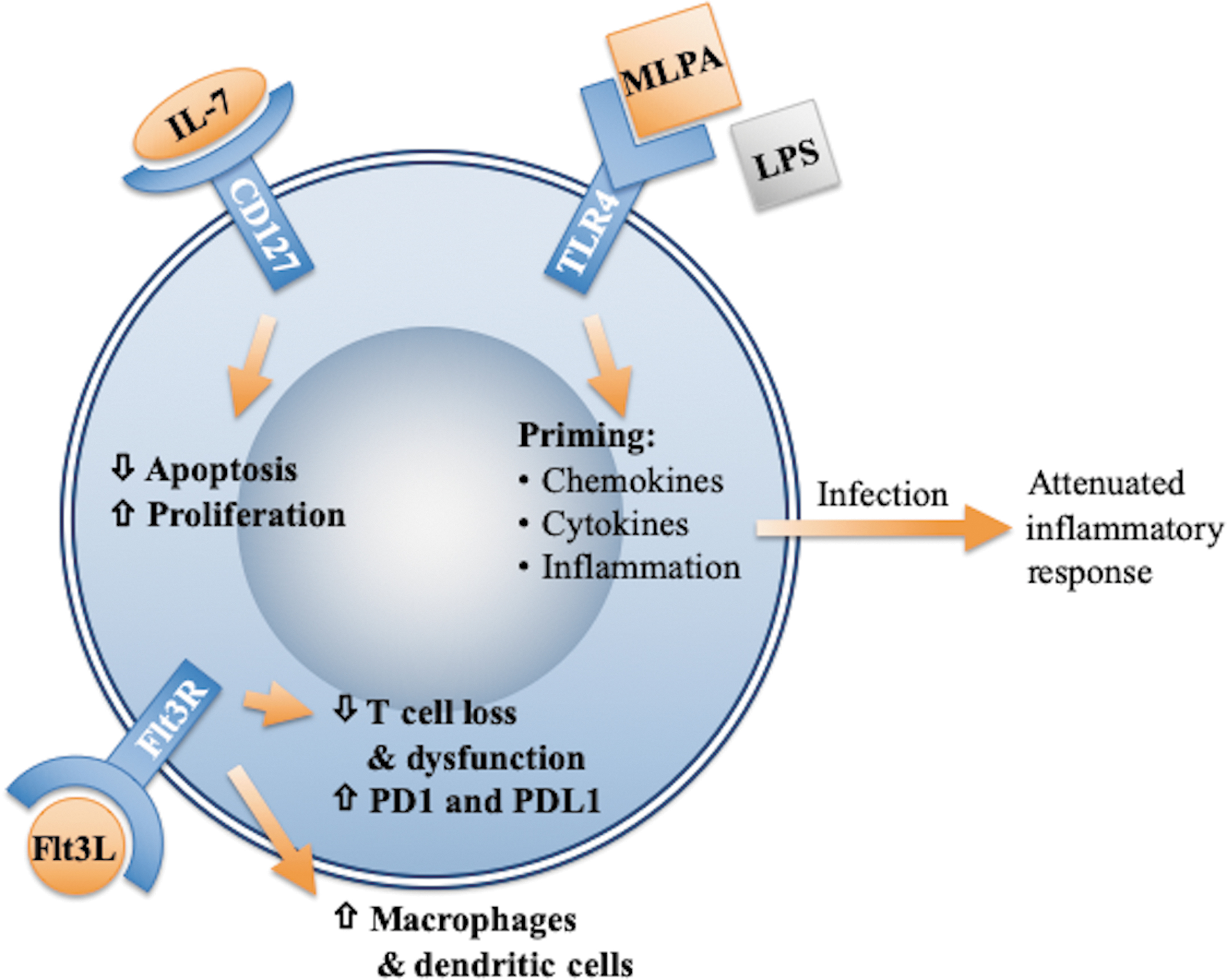
Leukocyte immune modulation. Burn injury can result in profound derangement of inflammatory response. In pre-clinical models, therapy with immune modulators has shown promise in improving outcomes. Modulation of TLR4 receptor pathway by monophosphate lipid A induces down-regulation of innate immune arm, whereas modulation by IL-7 and Flt3L pathways can up-regulate adaptive immunity.
Recombinant antibody therapies
Recombinant antibodies can be used to inhibit both virulence mechanisms in pathogens and dysfunctional immune signaling and have shown promise as therapeutic targets in burn infection. Specific monoclonal antibodies to virulent pathogenic structures and human signaling ligands can be produced using transgenic mice and cultured hybrid cells.
For example, the checkpoint inhibitor receptor programmed death-1 (PD-1) and its ligand on myeloid cells, PD-L1, are up-regulated in murine models of sepsis, as well as in septic human patients, inducing T-cell exhaustion, decreased proliferation, and apoptosis [46]. The efficacy of antibodies tailored to inhibit the PD-1/PD-L1 axis haa been demonstrated in murine models of sepsis [47]. Specific to burn injury, PD-1 and PD-L1 contribute to immune dysfunction, with a decrease in T-cell numbers and function: In murine burn injury, this correlates with significant multi-organ injury and shorter survival. In a murine model of burn injury with P. aeruginosa infection, treatment with anti-PD-L1 antibody reversed T-cell dysfunction, improved bacterial clearance, reduced organ injury, and enhanced survival [48].
Antibodies engineered against virulence factors have shown promise in animal models of burn injury, as shown in Figure 3. In a clinical study, antibodies were engineered against flagellin b and PilA, two virulence factors in P. aeruginosa, responsible for bacterial movement and invasion into tissue. Application as a vaccine in burned mice conferred protection against wound infection through improved opsono-phagocytic killing and inhibition of bacterial invasion into the host [49). As another example, the P. aeruginosa V antigen (PcrV) is an important factor in the type II secretion system (TTSS), which allows direct translocation of cytotoxins into host cells, initiating infection and eventually inducing cell death. Engineered antibodies to PcrV were administered as a vaccine to burned mice, which demonstrated increased survival after lung inoculation with P. aeruginosa and decreased lung inflammation and injury [50]. This antibody has been humanized for clinical use and is undergoing Phase II trials for ventilator-associated Pseudomonas pneumonia [51].
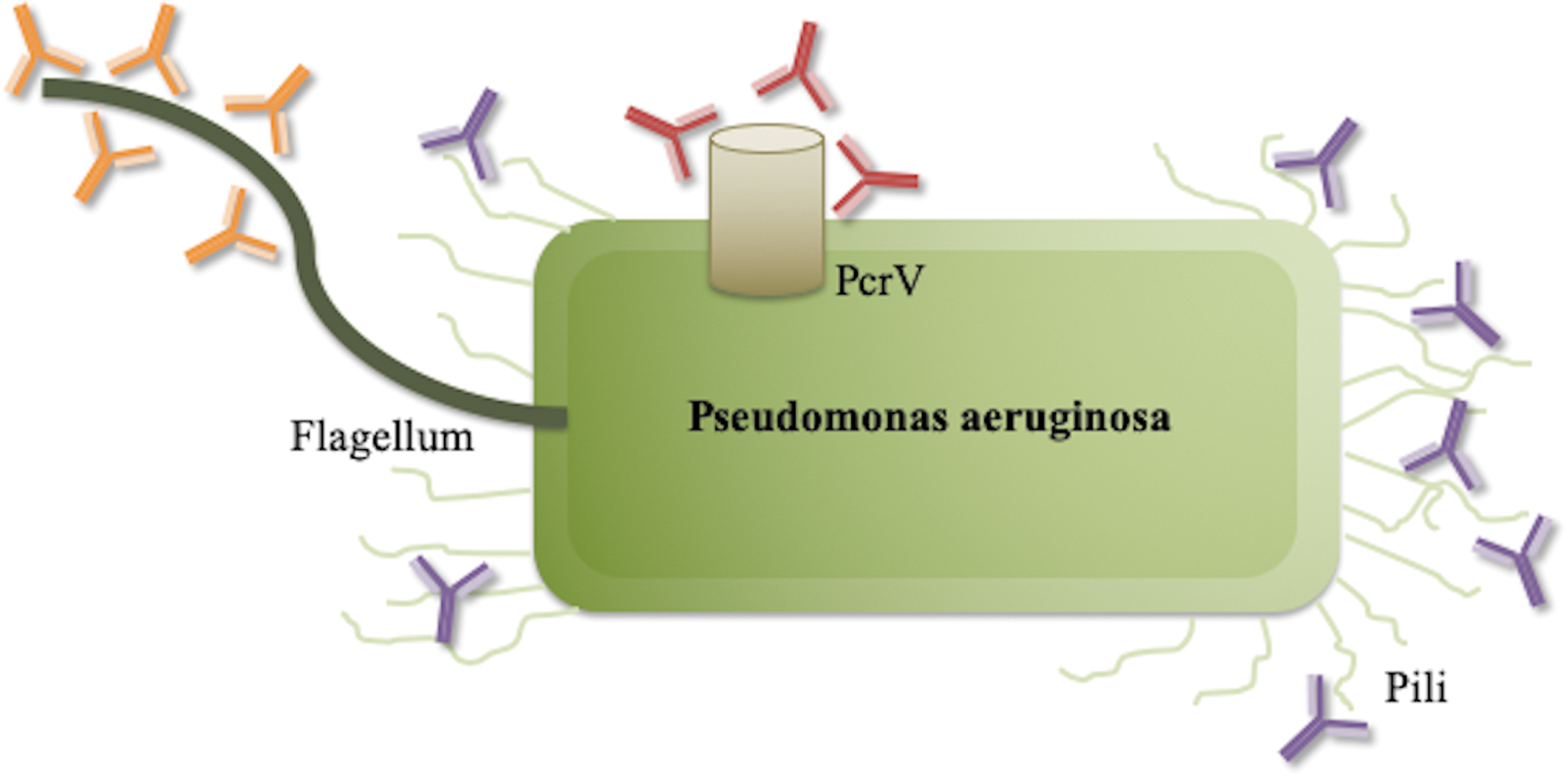
Use of antibodies engineered against virulence factors. Monoclonal antibodies can be created to bind to specific virulent pathogenic structures. A bacterial target especially important after burn injury is Pseudomonas aeruginosa. Antibodies were developed specifically against flagellin b (orange) and PilA (blue), two virulence factors in P. aeruginosa responsible for bacterial movement and invasion into tissue and PcrV (red), a secretion system allowing injection of endotoxin into host cells. In pre-clinical models, antibody-mediated inhibition of these virulence structures has shown promise in conferring protection against infection.
Thus, these studies have demonstrated efficacy, safety, and the encouraging potential of passive monoclonal antibody treatment in preventing infection in burn patients [52].
Extracellular vesicles and proteins
Antimicrobial peptides
Antimicrobial peptides (AMPs) are cationic, amphipathic oligopeptides that attach to and electrostatically disrupt the negatively charged LPS layer of the cell membrane [53]. These are produced naturally by a wide array of organisms and cell types, including skin, mucosa, and immune cells, and have been generated synthetically for therapeutic purposes. These peptides have emerged as a promising therapeutic agent in the management of burns given the broad spectrum of targeted organisms and low propensity for resistance owing to the nonspecific mechanism of action [54]. In addition to the physical disruption of the bacterial membrane, AMPs are capable of adhering to membrane receptors such as TLRs on immune cells, including dendritic cells, macrophages, and monocytes and inducing inflammation. Synthetic AMPS have been engineered for better selectivity and to minimize systemic toxicity. For example, the topical short synthetic β-sheet-folding peptide IK8L was superior to imipenem in a P. aeruginosa-infected mouse burn wound model in both bacterial clearance and overall survival [55].
Recently, two novel synthetic AMPs, WR12 and DIK8, were evaluated for efficacy against infection by multi-drug–resistant Staphylococcus. Both AMPs prevented stationary and intra-cellular growth of methicillin-resistant S. aureus (MRSA) in vitro better than did either vancomycin or linezolid. In mice, the AMPs significantly reduced the bacterial load and regulated the immune system, causing lower concentrations of pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α and IL-6 in MRSA-infected skin lesions [56]. A major potential limitation of AMPs is their toxicity to host tissues, as well as the -proteolytic degradation in vivo [57]. Further research will be needed prior to clinical studies.
Micro-vesicles
Micro-vesicles (MVs) are small, membrane-derived extracellular structures that contain lipids, proteins, or genetic information from parent cells. They are important in cell-to-cell communication, transporting biologically active information to their respective target cells [58]. One reported mechanism of MV generation is that after the externalization of acid sphingomyelinase, local ceramide and sphingosine (SPH) concentrations increase, and this ultimately leads to cellular budding and detachment of the MVs [59].
Pneumonia is a major cause of morbidity and death in burn patients. Sphingosine has been reported to have robust anti-microbial properties [60]. As mentioned previously, SPH is a part of membrane-derived MVs. It has been reported that mice have decreased lung MVs and a reduction in SPH after burn injury. After an intra-nasal application of bronchoalveolar lavage (BAL) MVs from healthy donors, burn-injured mice showed a decreased mortality rate and lung bacterial load while the SPH concentration was restored to that found in sham-treated mice [61]. It was further demonstrated that MVs can kill bacteria directly and even more effectively after supplementation with SPH. Rice et al. found a beneficial effect on the bacterial clearance of intra-peritoneally injected BAL MVs in septic mice, demonstrating that there is a systemic effect. In contrast, neutrophil-derived MVs in cell culture promoted bacterial growth [61]. Additionally, after burn injury, SPH is reduced in bronchial epithelial cells, whereas ceramide concentrations are increased [62]. Of note, SPH concentrations can be restored through direct inhalation or with neutral ceramidase, which converts ceramide to SPH [62]. Thus, SPH alone, or MVs with increased SPH, have the potential to be a potent, broad-spectrum antimicrobial agent. Testing is under way to determine the toxicity of inhaled SPH. A report has shown that repeated SPH treatment in mice over 10 days with doses as high as 1 mM caused no perceptible change in behaviour or histologic signs of toxicity [63]. Further testing needs to be conducted in large-animal models and in healthy human beings to strengthen the case for SPH as an excellent candidate for a novel inhaled antimicrobial drug.
Exosomes
Exosomes are the smallest subset of extra-cellular vesicles, categorized by their endocytotic origin and release after the fusion of a membrane-bound carrier with the lipid membrane [64]. They contain specific proteins, messenger ribonucleic acids (mRNAs), miRNAs, and lipids and have diverse functions in both the normal and inflammatory states [65].
Recent studies have shown that umbilical cord-derived mesenchymal stem cells secrete exosomes, which suppress inflammation in various animal models and disease states [66]. Burn injury significantly increases LPS-induced macrophage inflammation in rodent models, with increased TNF-α and IL-1β amounts and decreased IL-10 concentrations systemically. Administration of exosomes derived from mesenchymal stem cells reversed inflammation as assessed by a decrease in each one of these markers. Further research has shown that the responsible mechanism is down-regulation of the TLR4 pathway [67], as discussed previously in this review.
The use of sub-cellular vesicles and proteins in an immunomodulatory fashion will be dependent on accurate differentiation of the individual patient's immune status at the time of treatment. Much clinical research in the field of both burn and sepsis has focused on early therapeutic interventions aimed at suppression of the hyper-inflammatory response after the initial insult [68]. However, given concurrent immune deficiency and dysfunction, the degree of both inflammation and immunosuppression needs to be determined in order to administer an effective regimen [69]. Much of this is dependent on development of an immune staging system to identify the immune state of the individual patient better, in order to treat with the specific small particles that would alter the immune response beneficially.
Conclusion
With the growing threat of multi-drug–resistant pathogens in burn injury, novel therapeutic innovation remains imperative. Manipulation of structure and function of both host immune cells and pathogen virulence are at the forefront. There is promising evidence in pre-clinical models, with improvements of outcomes with targeted immune modulation, as well as encouraging human clinical trials under way. Understanding the pathophysiology of burn immune dysfunction, discovering new biomarker targets for therapy, and immune-stratifying patients according to their individual immune deficits will allow appropriate and better therapy in the burned patient population.
Footnotes
Authors' Contributions
CES, CB, NB, and CCC were responsible for the literature review, drafting the article, and preparation of figures. CES, CB, CP, and CCC performed critical revision of the article and final approval of the version to be published.
Funding Information
This project was supported by Shriners Hospitals for Children award number 85000-CIN-17 (CCC) and T32 GM08478 (CES) from the National Institute of General Medical Sciences (NIGMS) and by the Deutsche Forschungsgemeinschaft (DFG) (BE 7016/1-1). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIGMS, the National Institutes of Health, or the DFG.
Author Disclosure Statement
The authors declare no conflict of interest.