
Abstract
Background:
Necrotizing enterocolitis (NEC) is the leading gastrointestinal cause of death of premature neonates. We have previously shown that this hyperinflammatory state persists even post-recovery. We hypothesize that recovered patients with NEC will have a decreased hyperinflammatory response when the anti-inflammatory medication mesalamine (5-ASA) is administered even when exposed to in vitro NEC induction.
Methods:
Enteroids were generated and subjected to in vitro NEC induction. One half were subjected to 5-ASA treatment. Tumor necrosis factor-alpha (TNF-α) and interleukin 8 (IL-8) were evaluated via RT-qPCR. Mice underwent in vivo NEC induction, one group was given 5-ASA 50 mg/kg 12 h before the start of NEC induction. The intestine was harvested and assessed for hyperinflammatory markers and histological grading was performed.
Results:
Recovered NEC enteroids treated with 5-ASA during NEC induction show a significant decrease in inflammatory markers compared with control (p = 0.0014 TNF-α, downtrend IL-8). Active NEC enteroids treated with 5-ASA during in vitro NEC induction show a significant decrease in TNF-α RT-qPCR (p = 0.0443) and IL-8 RT-qPCR (p = 0.0265). In mice that received 5-ASA 50 mg/kg before in vivo NEC induction, there is a significant decrease in both TNF-α (p = 0.0114) and IL-8 (p = 0.0051).
Conclusion:
Enteroids and mice exposed to 5-ASA have a significant decrease in inflammatory markers. This decrease despite NEC induction in both enteroids and mice may demonstrate the impact that anti-inflammatory agents could have on treatment for NEC. This could be important given the robust hyperinflammatory response to a second hit after recovery and may impact the trajectory of an illness post-recovery from NEC.
Introduction
Necrotizing enterocolitis (NEC) is a known disease of premature neonates characterized by significant intestinal injury with severe morbidity and mortality.1,2 While the pathogenesis of the disease is not fully understood, there are associations such as prematurity, early enteral feeding, and increased hyperinflammatory response.3–5 The hyperinflammatory response in relation to NEC has been previously described by our laboratory and expands on the two-hit hypothesis.6,7 This hypothesis has been previously described in inflammatory bowel disease (IBD) literature, and we were able to demonstrate that patients with NEC are more hyperinflammatory at baseline and have a heightened response to additional injury, or a second hit as well.6–8 We were able to successfully delineate this hyperinflammatory process further in patients who have recovered from NEC. We sought to expand on this, particularly in the patients who have recovered from NEC, to explore the hyperinflammatory response after a second hit with potential anti-inflammatory agents to ameliorate this response post-recovery.
While IBD and NEC are differing disease processes, the basis of a heightened inflammatory state, altered gastrointestinal flora, and damage to intestinal mucosa and barrier are cornerstones to the pathophysiology of both.3,4,9 Treatment for NEC currently consists of supportive care with antibiotic agents, bowel rest, and in its most severe form, surgery. Mesalamine or 5-ASA is often used as first-line treatment for mild-to-moderate ulcerative colitis.10,11 We postulated that this response could be modulated in NEC. To explore this, we studied hyperinflammatory markers tumor necrosis factor-alpha (TNF-α) and interleukin 8 (IL-8) following the administration of 5-ASA in our human enteroid NEC model and our mouse NEC model. We chose to focus mainly on IL-8 as literature demonstrates that IL-8 serum concentration changes may be a useful marker to monitor disease. 12
We hypothesize that active and recovered patients with NEC will have a decreased hyperinflammatory response following 5-ASA administration even when exposed to in vitro NEC induction and that mice undergoing in vivo NEC induction will have a decreased hyperinflammatory response following 5-ASA administration before NEC induction.
Patients and Methods
Human samples
Following institutional review board approval (IRB #11610-11611) and parental consent, human neonatal intestinal tissue was collected from patients undergoing bowel resection at the Oklahoma Children’s Hospital in Oklahoma City, OK. For this study, all infants were premature infants who were included, with one patient with active NEC, one patient without NEC, and one patient who had fully recovered from NEC. Tissue specimens were divided after collection and subsequently in cold Dulbecco’s phosphate-buffered saline (DPBS, Sigma Life Science, #D8573, St. Louis, MO), and then kept in Roswell Park Memorial Institute media (Gibco, #11875-093, Waltham, MA) for 24 hours or less until processed for enteroid culture. Enteroids were developed from each of the three patients used for this particular set of experiments.
In vitro human neonatal enteroid cultures
Following collection, within 24 hours, intestinal crypts were isolated from the samples, and enteroid cultures were generated as per our previously described protocol used for crypt isolation and enteroid processing. 13 The enteroids were then suspended in basement membrane matrix, Matrigel (Corning, #CB-40230C, Corning, NY) domes and grown in 50% L-WRN-conditioned media, which is supplemented with 50 ng/mL epidermal growth factor (Millipore Sigma, #GF144, Burlington, MA), 1 mM N-acetylcysteine (Millipore Sigma, #A9165-5G), 500 nM A-83-01 (R&D Systems, #2939/10, Minneapolis, MN), 10 mcM SB202190 (Millipore Sigma, #S7067-5 MG), 10 mM nicotinamide (Millipore Sigma, #N0636-100G), and 10 nM [leu] 15-gastrin 1 (Millipore Sigma, #G9145-.1 MG). All enteroids were passaged every 5–7 days and experimented on when at full maturity. All experiments were performed between passages 3 and 10.
Enteroid NEC experimentation
The enteroid cultures were grown in the abovementioned basement membrane matrix, Matrigel, with the supplemented media for 7–21 days. For all experiments, the enteroids were mature at the time of experimentation determined by blebbing and ensuring they were 7–10 days from the last passage. Our previously described protocol for enteroid NEC induction in vitro was followed for 24 hours. 7 For each set of experiments, to induce NEC, enteroids underwent treatment consisting of 100 mcg/mL lipopolysaccharide injection into the supplemented media along with 24 hours of hypoxia. This time point and LPS dose with hypoxia were followed again as per prior published protocols to induce in vitro NEC without causing frank necrosis.13–15
Within each set of enteroids, one half of each plate underwent media supplementation with 5-aminosalicylclic acid mesalamine (5-ASA) (Fisher Scientific, #NC1087013, Waltham, MA) with 5 mM concentration for each. This was chosen based on prior IBD literature and a dose curve performed as described below. 16 Enteroids derived from three different patients were used for all experiments. A dose curve NEC induction experiment was performed on enteroids derived from a patient without NEC with media supplementation with 5-ASA (Fisher Scientific, #NC1087013) in the following concentrations 0 mM, 1 mM, and 10 mM. Concentrations of TNF-α were chosen to be measured as this inflammatory marker is frequently upregulated in NEC. All 5-ASA was dissolved in warm 50% L-WRN-conditioned media described above. For each experiment, enteroids that underwent NEC induction without 5-ASA were compared with those that underwent NEC induction without 5-ASA supplementation. All experiments were performed in triplicate to enhance scientific rigor.
RNA isolation, reverse transcription, and RT-qPCR of enteroids
Total RNA was isolated and extracted using TRIzol (Life Technologies, Carlsbad, CA). RNA was quantified for purity (2.0–2.1 A260/A280 ratio) and quantity using a NanoDrop Lite spectrophotometer (Thermo Scientific, Waltham, MA). RNA was then reverse transcribed to 0.5–2 mcg of complementary DNA (cDNA) using the high-capacity cDNA reverse transcription kit (Applied Biosystems, #4374966, Waltham, MA). Real-time PCR results were obtained using the CFX Opus 96 system, an iQ SYBR Green Supermix (Bio-Rad, Hercules, CA) with 4 ng of cDNA template, and the final primer concentration was 0.5 mcM. Expression levels were quantitated and normalized to GAPDH. The following primers were used for amplification of target cDNA: TNF-α, 5′ TGA GGT ACA GGC CCT CTG AT 3′ and 5′ CTC GAA CCC CGA GTG ACA AG 3′; IL-8, 5′ GGC CGT GGC TCT CTT GGC AG 3′ and 5′ TGT GTT GGC GCA GTG TGG TCC 3′; GAPDH, 5′ TCA AGG CTG AGA ACG GGA AG 3′ and 5′ CGC CCC ACT TGA TTT TGG AG 3′.
Enzyme-linked immunosorbent assay
For experiment comparisons, the enteroid media were isolated and enzyme-linked immunosorbent assay (ELISA) was performed for IL-8 (KHC0081) (Invitrogen, Waltham, MA).
Mouse samples
Institutional Animal Care and Use Committee (IACUC) approval (IACUC Protocol # 20-014) was obtained prior to any experimentation. Wild-type mice C57BL/6J were purchased for breeding for experimentation (#000664, Jackson Laboratory, Bar Harbor, ME). Breeding was started after proper acclimation time and the subsequent mouse pups were subjected to the in vivo NEC experiment on day of life 14.
In vivo NEC induction
In vivo NEC induction was performed on day of life 14 for the mouse pups. 5-ASA (Fisher Scientific, #NC1087013) was gavage fed to the mouse pups at the following doses: 0 mg/kg (PBS, D1408, Sigma Aldrich, St. Louis, MO) and 50 mg/kg 12 h prior to intraperitoneal dithizone (33 mg/kg) (43820, Sigma Aldrich) injection. The mouse pups are then placed in the incubator at 32°C without confounding food or water. Six hours following dithizone injection, each mouse pup was gavage fed Klebsiella (10031, American Type Culture Collection, Manassas, Virginia) in a solution of PBS in dosing of 1 × 108 CFU/g body weight. The mouse pups are then kept in the incubator with physical assessments every 30 minutes for signs of clinical distress. The mouse pups are sacrificed at 10 hours post-Klebsiella administration and intestinal tissue was collected for snap-frozen specimens and histological grading.
Histological grading
Intestine was harvested and graded by hematoxylin and eosin for injury scoring by three independent clinician reviewers blinded to experimental groups on a scale of grade 1 to grade 3.
RNA isolation, reverse transcription, and RT-qPCR of intestinal tissue
Total RNA isolation was performed from snap-frozen tissue beginning with homogenization using the Fisher Brand Pellet Pestle cordless motor (Fisher Scientific, Hampton, NH) and VWR disposable pestles (VWR, Radnor, PA). This was followed by RNA isolation and extraction using TRIzol (Life Technologies). RNA was quantified for purity (2.0–2.1 A260/A280 ratio) and quantity using a NanoDrop Lite spectrophotometer (Thermo Scientific). RNA was then reverse transcribed to 0.5–2 mcg of cDNA using the high-capacity cDNA reverse transcription kit (Applied Biosystems, #4374966). Real-time PCR results were obtained using the CFX Opus 96 system, an iQ SYBR Green Supermix (Bio-Rad) with 4 ng of cDNA template, and the final primer concentration was 0.5 mcM. Expression levels were quantitated and normalized to GAPDH. The following primers were used for amplification of target cDNA: TNF-α, 5′-CGGGCAGGTCTACTTTGGAG-3′ and 5′-ACCCTGAGCCATAATCCCCT-3′; IL-8, ACCTAGGCATCTTCGTCCGT-3′ and 5′-GCCAACAGTAGCCTTCACCC-3′; GAPDH, 5′ TCA ACG GGA AGC CCA TCA 3′ and 5′ CTC GTG GTT CAC ACC CAT CA 3′.
Results
Human specimens
All specimens were obtained from three patients who underwent intestinal surgeries, including for active NEC, ostomy reversal following recovery from NEC, and intestinal atresia repair. For each specimen obtained, the anatomical site was noted along with the specific surgical procedure and patient surgical history. The age at birth, age at resection, and corrected gestational age are also noted for comparison purposes. Corrected gestational age at time of resection ranged from 36 weeks 2 days to 46 weeks and 6 days. For the patient who underwent an operation for active NEC, the specimen was collected at the initial exploratory laparotomy during the active NEC episode. For the patient who underwent the procedure for an ostomy takedown following recovery from NEC, this was determined by the patient’s ability to tolerate enteral feeds, having adequate ostomy output, and vital signs and laboratories within normal limits before reversal. This particular operation occurred almost four months following the active NEC episode that patient was initially taken to the operating room for. For the patient who underwent an operation for intestinal atresia, the specimen was collected during the patient’s ostomy reversal following the intestinal atresia operation almost two months after the initial operation.
Enteroids derived from a patient without NEC demonstrating a dose curve response to 5-ASA administration
A dose curve was performed in increments of 0 mM, 1 mM, and 10 mM. With the dose curve and combination with the literature review, we selected the dose of 5 mM. 16 This demonstrated a significant decrease in the inflammatory marker TNF-α from 0 mM to 10 mM (p = 0.0170) and from 1 mM to 10 mM (0.0308) (Fig. 1).
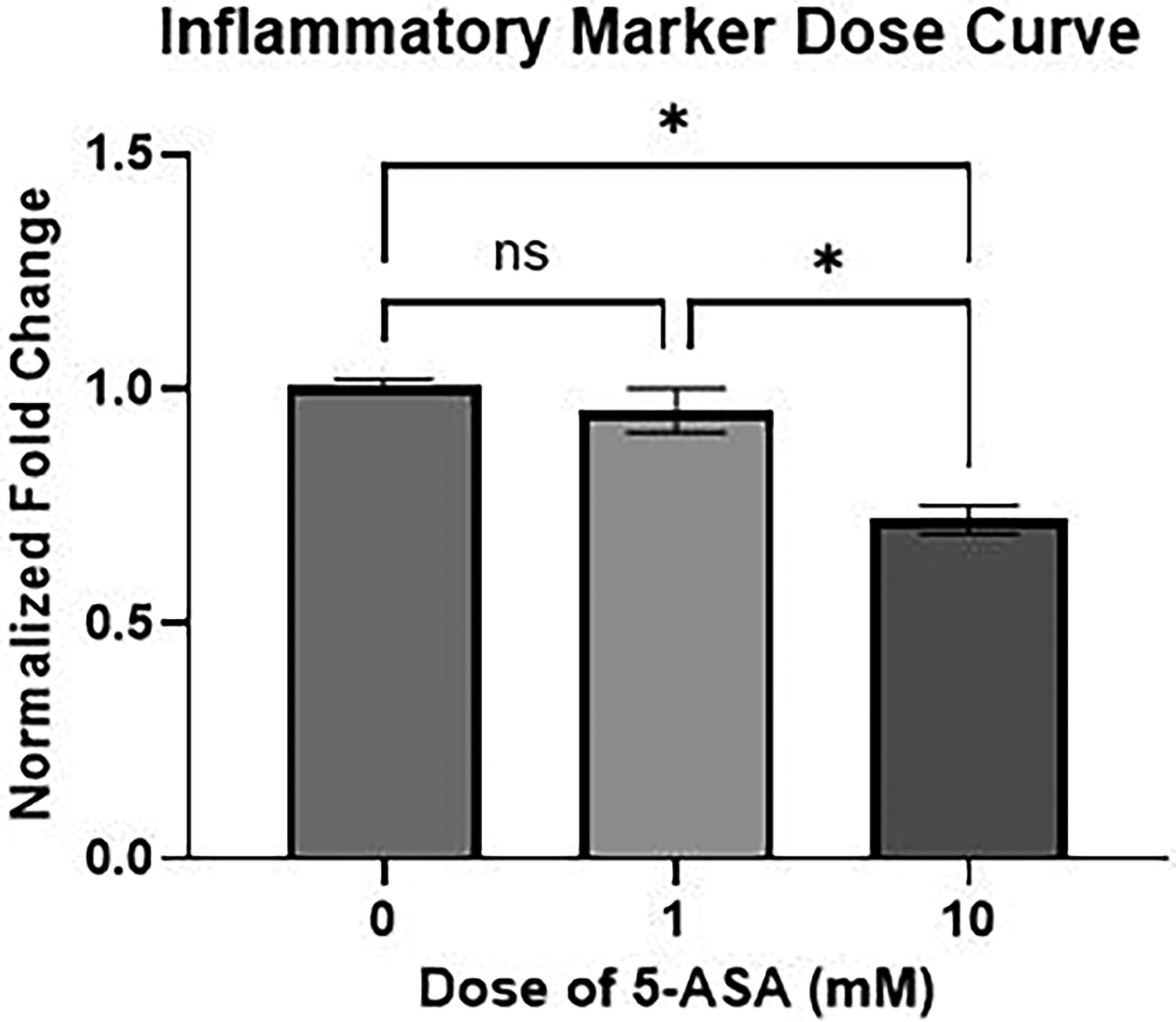
Dose curve performed on control enteroids demonstrating a significant decrease in inflammatory marker tumor necrosis factor-alpha (TNF-α) from 0 mM to 10 mM (p = 0.0170) and from 1 mM to 10 mM (0.0308). *p < 0.05.
Enteroids derived from tissue from a patient fully recovered from NEC compared with enteroids derived from that same patient supplemented with 5-ASA
When comparing enteroids that underwent NEC induction without 5-ASA supplementation with the enteroids that underwent NEC induction with 5-ASA administration, we were able to see a significant decrease in the amount of inflammatory markers present when supplemented 5-ASA in RT-qPCR for TNF-α (p = 0.0014) and a strong downward trend with IL-8. This was further demonstrated for IL-8 via ELISA analysis with a significant decrease seen in the recovered enteroids that underwent induction with 5-ASA supplementation compared with those that underwent induction without 5-ASA supplementation (p = 0.0026) (Fig. 2). These results indicate that with supplementation of 5-ASA, it is possible to decrease the primed hyperinflammatory state that we see in enteroids derived from a patient who has recovered from NEC. 6 This could ameliorate the response in these patients to a second hit.
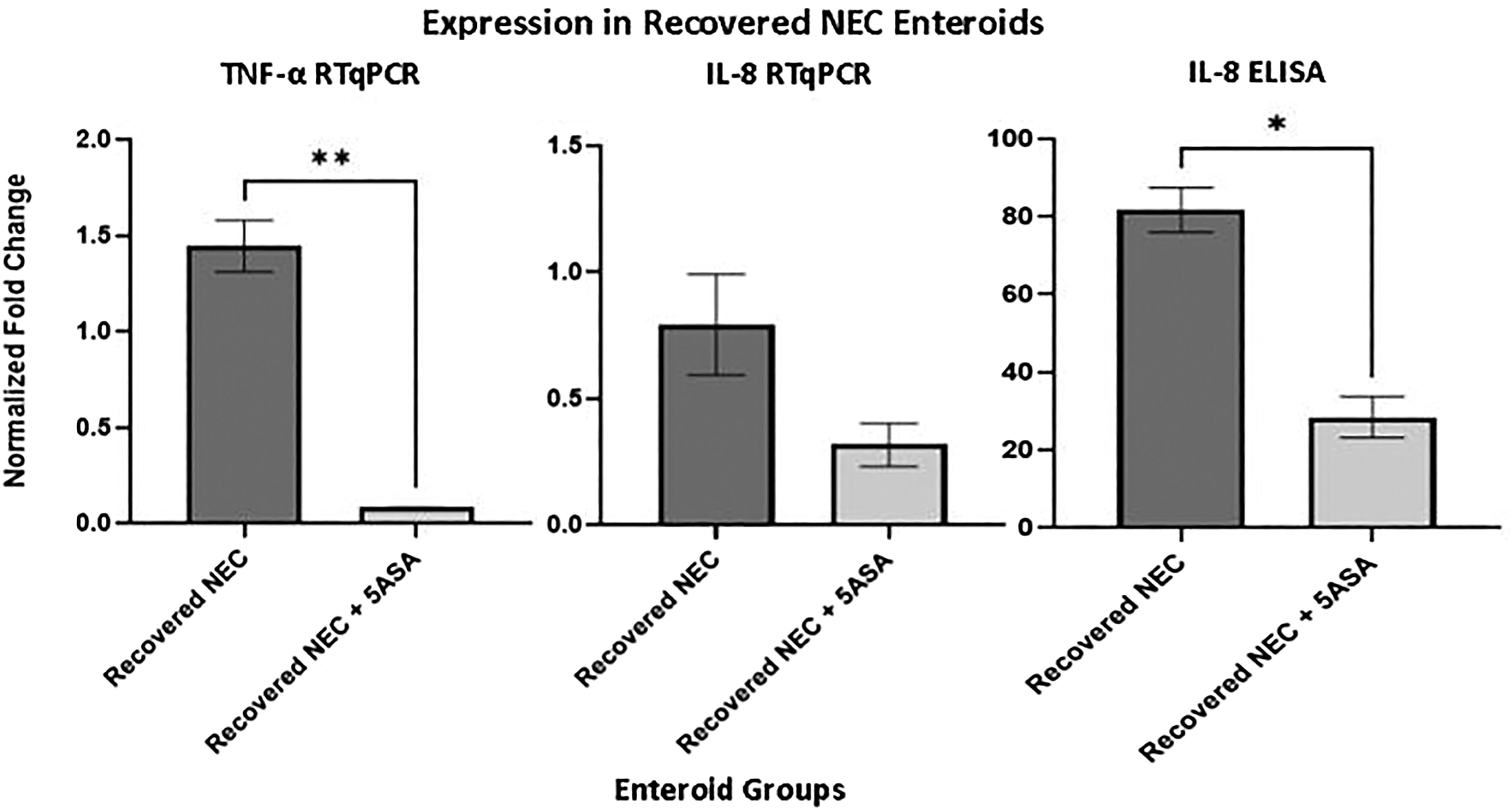
Inflammatory marker expression in recovered necrotizing enterocolitis (NEC) enteroids with 5-ASA supplementation following NEC induction. There is a significant decrease in the amount of inflammatory markers present when supplemented 5-ASA in RT-qPCR for tumor necrosis factor-alpha (TNF-α) (p = 0.0014) and a strong downward trend with interleukin 8 (IL-8). There is also a significant decrease in IL-8 seen on enzyme-linked immunosorbent assay (ELISA) (p = 0.0026). *p < 0.05; **p < 0.01.
Enteroids derived from tissue from a patient with active NEC compared with enteroids derived from that same patient supplemented with 5-ASA
When subjected to 5-ASA supplementation, enteroids that underwent in vitro NEC induction demonstrate a significant decrease in TNFα RT-qPCR (p = 0.0443), IL-8 RT-qPCR (p = 0.0265), and ELISA (p = 0.0055), similar to what is seen in the recovered NEC enteroid group (Fig. 3). This could indicate a potential avenue for additional medical therapy during an active NEC infection to potentially aid in the avoidance of a surgical procedure for patients by dampening the hyperinflammatory response and intestinal injury.

Inflammatory marker expression in active necrotizing enterocolitis (NEC) enteroids with 5-ASA supplementation following NEC induction. When subjected to 5-ASA supplementation, there is a significant decrease in tumor necrosis factor-alpha (TNF-α) RT-qPCR (p = 0.0443), interleukin 8 (IL-8) RT-qPCR (p = 0.0265), and enzyme-linked immunosorbent assay (ELISA) (p = 0.0055). *p < 0.05; **p < 0.01.
Mouse specimens
All specimens were obtained from 36 days of life for 14 mice following the in vivo NEC conditions with 5-ASA administration described above. For each specimen obtained, the anatomical site taken for the snap-frozen tissue and for the histological specimen was noted.
Tissue samples from mice that did not have 5-ASA supplementation compared with mice that had 50 mg/kg of 5-ASA supplementation
Mice were equally and randomly divided into groups for 5-ASA administration during the in vivo NEC induction experiment described above. Tissue from mice that were given 5-ASA 50 mg/kg 12 h prior to the start of NEC induction showcase a significant decrease in the amount of TNF-α (p = 0.0114) and IL-8 (p = 0.0051) on RT-qPCR (Fig. 4). By studying this, we are able to confirm what is seen above in the enteroid model in an in vivo setting, opening up potential avenues to further this study in patients with NEC.
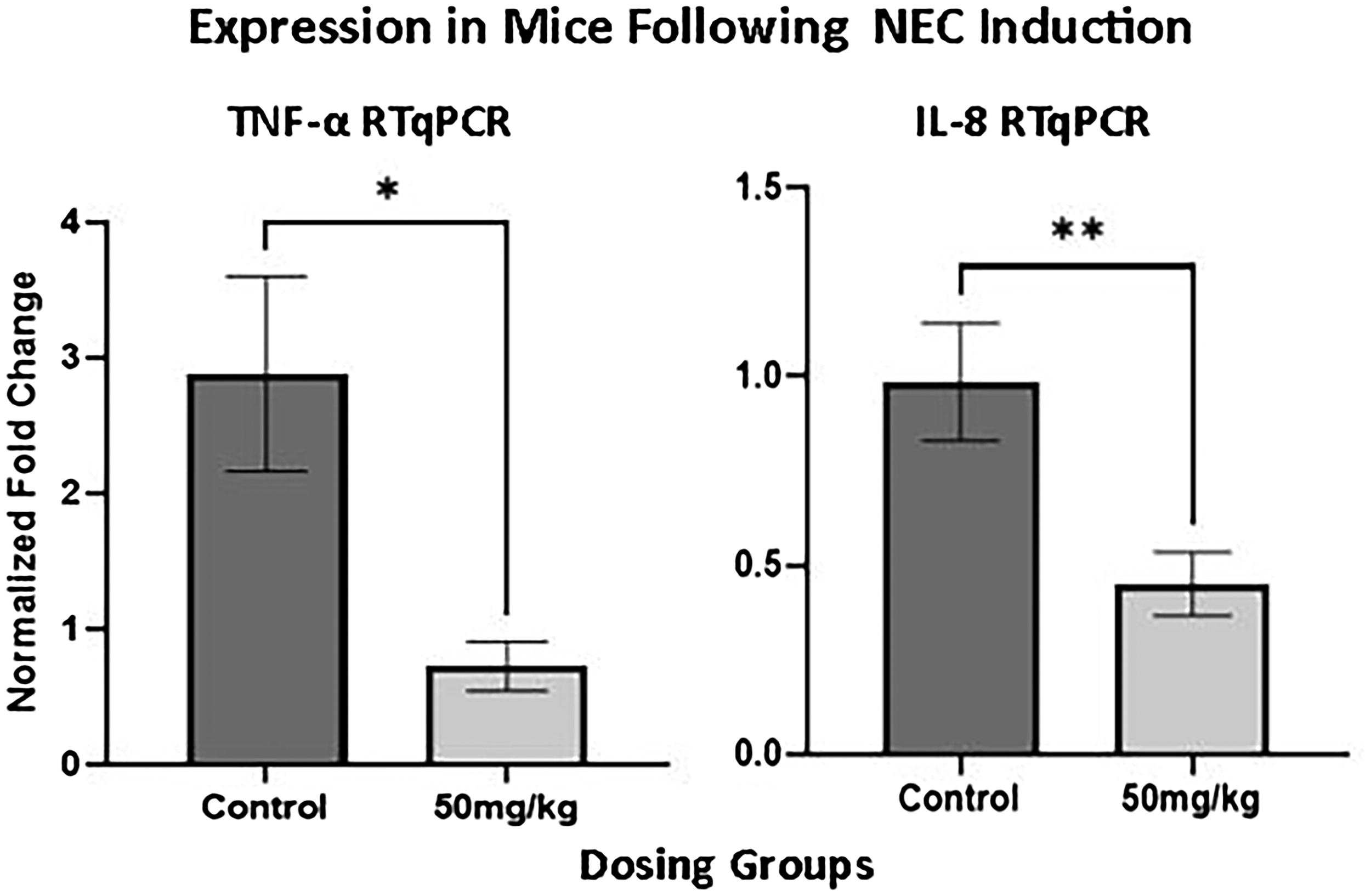
Tissue from mice following in vivo necrotizing enterocolitis (NEC) induction with 5-ASA supplementation. Mice that were given 5-ASA 50 mg/kg 12 h before the start of NEC induction showcase a significant decrease in the amount of tumor necrosis factor-alpha (TNF-α) (p = 0.0114) and interleukin 8 (IL-8) (p = 0.0051) on RT-qPCR. *p < 0.05; **p < 0.01.
Histological grading of NEC from mice that did not have 5-ASA supplementation and mice that did have 50 mg/kg of 5-ASA supplementation
All tissue specimens were graded histologically on an NEC scale from grade 0 (normal) to grade 3 (severe). An injury severity score greater than 1.5 was considered NEC. 17 Histological scoring revealed a significantly greater injury score in the control group of mice not given 5-ASA administration prior to in vivo NEC induction compared with mice that were given 5-ASA at a dose of 50 mg/kg prior to in vivo NEC induction (p = 0.0206) (Fig. 5).
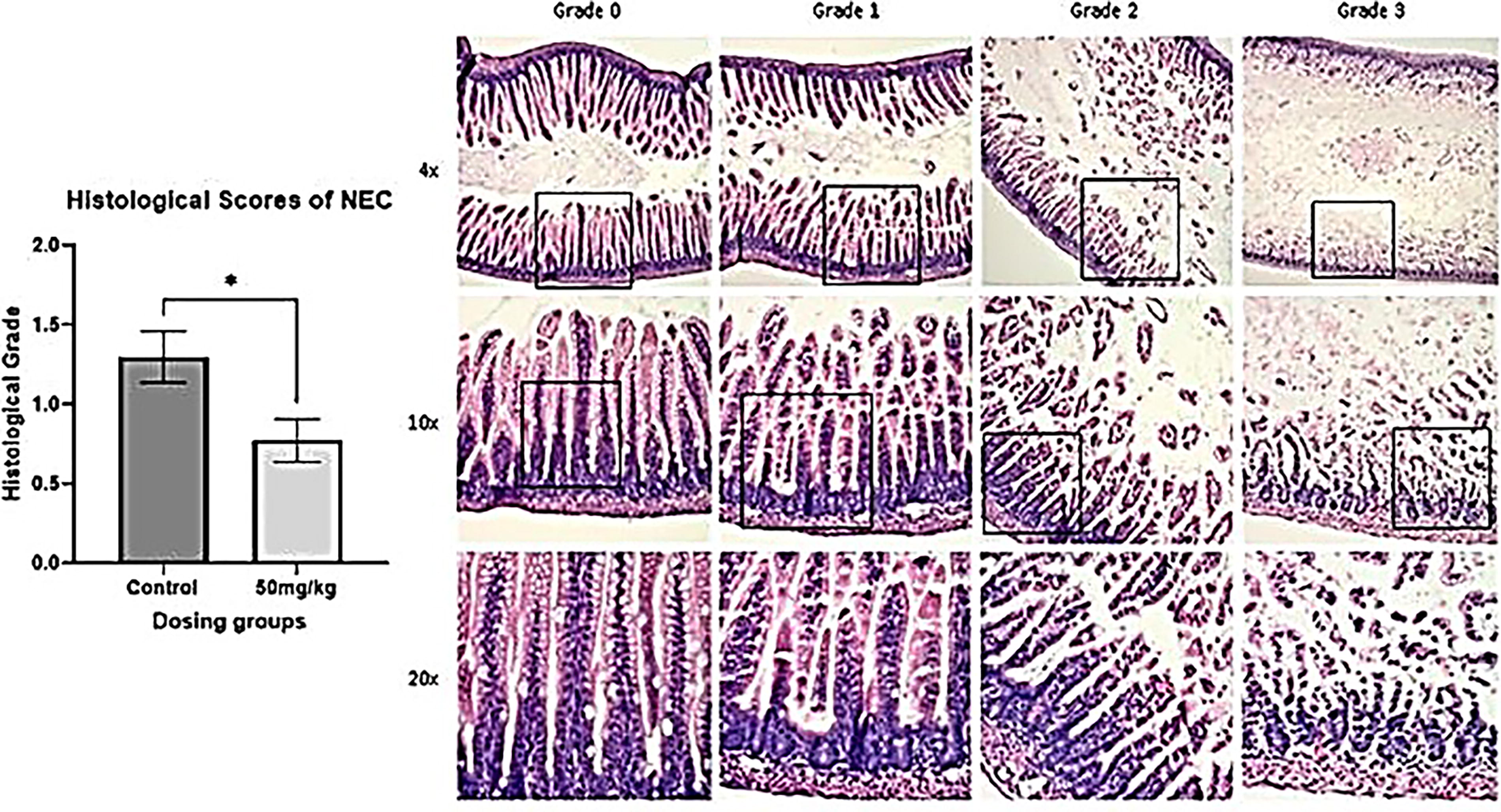
Histological scores of necrotizing enterocolitis (NEC) on a scale of 1–3. There is a significantly greater injury score in the control group compared with mice that were given 5-ASA at a dose of 50 mg/kg before in vivo NEC induction (p = 0.0206). *p < 0.05.
Discussion
The hyperinflammatory cascade is well established to be at play in NEC.6,7,13,15,18,19 Our laboratory previously has demonstrated data indicating a persistent proclivity to a hyperinflammatory state in infants who develop NEC and delineated the impact of a second hit on the patients who do recover from NEC.6,7 We have also previously demonstrated that enteroids derived from these patients provide an excellent model for studying NEC in vitro as the enteroids maintain the characteristics from the host from which they were obtained.6,7,13,14 The retained memory that enteroids demonstrate allowed us to study the impact of a second hit on patients who had recovered NEC in prior studies, and now has allowed us to evaluate the effect of the anti-inflammatory agent 5-ASA on these patients during the in vitro second hit. 6
IBD often is a relapsing and remitting inflammatory disease of the gastrointestinal tract. 20 This can be seen to a degree in NEC as well, as it has been shown that patients who have recovered from NEC retain the tendency to enter into the hyperinflammatory state no different than that of active NEC when subjected to another insult. 6 This is seen clinically through patients who have survived NEC presenting with issues such as failure to thrive, intestinal strictures, and intestinal failure as they age.21–25 We were able to expand on the two-hit hypothesis within the realm of NEC and potential presentations of patients following recovery in our prior studies. 6 This was done by demonstrating that the enteroids derived from a patient who has recovered from NEC experienced a hyperinflammatory response elevation no different than that of enteroids derived from a patient during an active NEC episode. 6
NEC currently does not have a treatment regimen along the same lines as what is seen in the world of IBD. As previously discussed, 5-ASA is often the first-line treatment for the relapsing and remitting ulcerative colitis. The exact mechanism of action of 5-ASA remains to be fully elucidated. However, it is thought to have an effect on the cyclooxygenase and lipoxygenase pathways, reducing the formation of proinflammatory prostaglandins, leukotrienes, and cytokines.11,26,27 The side effect profile of 5-ASA is overall negligible making it a safe and effective medication for both children and adults.11,28 Given the proposed mechanistic action of 5-ASA, we postulated that it may have a beneficial effect in NEC as well, especially given what our previous studies have shown with heightened inflammation post-recovery when subjected to an insult.
Our studies demonstrate that this is the case in not only the enteroids developed from a patient who has fully recovered from NEC, but also in enteroids developed from a patient who had active NEC. Our previous study demonstrated that after being subjected to a second hit, recovered NEC enteroids were in a hyperinflammatory state no different than in active NEC. 6 We are currently able to show that those same enteroids, when subjected to 5-ASA prior to the second hit, have inflammatory markers significantly decreased.
We were able to take this further through the use of an animal model. In our mouse in vivo NEC model, mice given 5-ASA prior to the start of NEC induction did not develop as severe NEC as demonstrated by histology and with significantly decreased inflammatory markers compared with mice that did not receive 5-ASA prior to the start of induction.
To our knowledge, this is the first study of this kind with regard to NEC. By utilizing our well-established NEC protocols and building upon our most recent studies regarding the second hit hypothesis within NEC, we were able to demonstrate the impact of an anti-inflammatory agent after recovery from NEC in enteroids and during active NEC.
This study could have significant implications for treatment algorithms going forward for active NEC by decreasing the hyperinflammatory response and bowel injury in these patients. This could also be significant in patients who have recovered from NEC to prevent potential problems such as strictures by decreasing the primed response to a second hit.
Conclusion
NEC is a devastating disease with potential hyperinflammatory impacts on patients who have recovered age. Through expansion of our studies, we have now been able to demonstrate the impact of the anti-inflammatory agent, 5-ASA, on not only recovered and active NEC enteroids but also in a mouse model. The persistence of this primed hyperinflammatory response following recovery from NEC has provided us an avenue to potentially study the impact of an anti-inflammatory agent such as 5-ASA clinically to help these patients as they grow post-recovery.
Footnotes
Authors’ Contributions
K.B.S.: Conceptualization, methodology, validation, formal analysis, investigation, resources, data curation, writing, review, editing, and visualization. C.D.: Methodology, formal analysis, and review and editing. G.G.: Methodology, formal analysis, and review and editing. C.S.: Methodology, investigation, and formal analysis. A.M.: Methodology, formal analysis, and review and editing. C.L.C.: Methodology and review and editing. J.E.: Methodology and investigation. W.B.: Methodology and investigation. C.J.H.: Conceptualization, methodology, validation, resources, writing, review, editing, visualization, supervision, project administration, and funding acquisition.
Ethical Statement
The authors are accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
Oklahoma Center for Adult Stem Cell Research (CH): Research reported in this article was supported by the National Institutes of Health NIDDK under award number 1R01DK133207-01A1.