
Abstract
Abstract
A mesenchymal stem cell (MSC)-seeded collagen gel under static or dynamic tension is a well-established model to study the potential of MSCs in regenerating a tendon- or ligament-like tissue. Using this model, upregulation of fibrillar collagen mRNA expression and protein production has been demonstrated in response to cyclic tensile mechanical stimulation. However, the mechanisms driving MSC tenogenesis (differentiation into tendon or ligament fibroblasts) have not been elucidated. This study investigated the mechanisms of tenogenesis of human bone marrow–derived MSCs in a dynamic, three-dimensional (3D) tissue-engineering model by investigating the effects of cyclic stretching on matrix production and gene expression of candidate tendon and ligament markers. The 3D MSC tenogenesis culture system upregulated scleraxis, but cyclic stretching was required to maintain expression of this putative tendon marker over time. Enhanced tendinous neo-tissue development demonstrated with extracellular matrix staining was largely due to changes in matrix deposition and remodeling activity under dynamic loading conditions, as evidenced by differential regulation of matrix metalloproteinases at a transcriptional level with minimal changes in collagen mRNA levels. Regulation of Wnt gene expression with cyclic stimulation suggested a similar role for Wnt4 versus Wnt5a in tenogenesis as in cartilage development. This first report of the potential involvement of matrix remodeling and Wnt signaling during tenogenesis of human MSCs in a dynamic, 3D tissue-engineering model provides insights into the mechanisms of tenogenesis in a mechanoactive environment and supports the therapeutic potential of adult stem cells.
Introduction
MSCs have the potential to differentiate readily along various mesenchymal lineages, including bone, cartilage, muscle, adipose, and stroma.6,7 The most common method to direct MSCs towards different lineages in vitro is through the use of supplements in the culture medium. In contrast, attempts to differentiate MSCs into tendon or ligament fibroblasts (tenogenesis) have relied largely upon uniaxial tension in three-dimensional (3D) constructs. MSC-seeded collagen gels under uniaxial tension have been used to create engineered replacements for tendons and ligaments. In this system, MSCs remodel the surrounding collagen fibrils and orient themselves and the matrix along the axis of tension. Variations of these models have been reported, but the underlying constant is the creation of a mechanoactive environment due to endogenous cell traction or external stimulation. For instance, Awad et al. contracted an MSC–collagen gel on a pre-tensioned suture, characterized the contraction rate and degree of cell alignment and elongation under static conditions, and hypothesized that the tensioned suture provided crucial initial alignment cues to the MSCs. 8 They speculated that resistance to traction provided by the collagen fibers and scaffold suture resulted in transmission of forces to the cells that led to changes in cellular and nuclear morphology necessary to switch cells from proliferation to differentiation. MSC–collagen gel constructs have been tested in vivo in patellar tendon defects in a number of studies and consistently demonstrated to repair tissues with better mechanical properties than contralateral controls.9–11 Other studies have shown that dynamic mechanical forces can promote tendon and ligament differentiation. Altman and colleagues showed that dynamic loading could promote differentiation of MSCs cultured in a collagen gel between bone anchors to create bone–ligament–bone constructs. 12 Mechanical stimulation over a period of 21 days upregulated collagen types I and III and tenascin-C and induced significant cell and collagen fiber alignment. Recently, Noth et al. published a similar study in which an MSC–collagen gel construct was cast between bone anchors and cyclically loaded for 14 days to increase mRNA levels of collagen types I and III, elastin, and fibronectin. 13 Although these studies have advanced the development of tendon and ligament constructs using MSCs, they have not specifically focused on investigating the tenogenic process itself, particularly whether the differentiation of MSCs in an engineered tissue model replicates embryonic tendon development. Understanding tenogenesis in vitro is critical for developing adequate strategies for engineering functional tendon and ligament constructs.
The major challenge in the engineering of tendons and ligaments has been the lack of unique biological markers available to characterize MSC differentiation. This is due mainly to insufficient understanding of the developmental process of tendons and ligaments. A key regulatory transcription factor to characterize tenogenic differentiation has yet to be found, challenging the ability to truly characterize whether MSCs are undergoing differentiation down a tendon or ligament pathway. Recently, scleraxis, a basic helix–loop–helix transcription factor, was shown to be specifically associated with embryonic tendon and ligament development.14,15 Its expression was shown in presumptive tendon progenitor cells during embryogenesis and was continuous through tendon development. Although scleraxis function has yet to be elucidated, development of force-transmitting and intermuscular tendons is disrupted in scleraxis-null mice, suggesting that scleraxis function is required for normal tendon formation. 16
Expression of growth factors, signaling molecules, and matrix components may also serve as markers of differentiation. Most studies have used the latter in characterizing tenogenesis, including collagen types I and III, elastin, and fibronectin. Few have used cellular signaling molecules as in vitro markers. Wnts and matrix metalloproteinases (MMPs) have been shown to be critical to developmental programs in embryonic cells and MSCs. Wnts are secreted glycoproteins that bind to frizzled receptors to activate specific signaling pathways. A number of groups have directed osteogenic differentiation of MSCs by manipulating Wnt signaling, both canonical and non-canonical.17–21 The involvement of Wnts in tenogenic differentiation of human MSCs has not been investigated. However, during embryogenesis, Wnt4, Wnt14, and Wnt16 have been identified in non-cartilaginous mesenchymal cells that will form tendons and ligaments.22,23 Also involved in skeletal development are MMPs, a family of zinc metallo-endopeptidases that cleave various components of the extracellular matrix (ECM). MMPs are critical in tissue embryogenesis, including skeletal development, 24 and it is likely that they play a role in adult tissue maintenance of tendon and ligaments. However, the role of MMPs in embryonic tendon and ligament development remains to be elucidated, although it is likely that MMPs are involved in active matrix remodeling during tendon and ligament development.
A cell-seeded collagen gel under tension is an established model that has been used, with the primary aim of creating a functional tendon or ligament replacement, but the cellular and molecular mechanisms of the tenogenic differentiation process of MSCs in this 3D environment have not been elucidated. Current assays for tenogenesis are functional and based mainly on collagen expression. The purpose of this work was to investigate whether tenogenesis of MSCs in vitro follows a similar cellular and molecular program as embryonic development in vivo, and if mechanical loading is necessary to maintain this program. Therefore, we investigated the role of the 3D environment in establishing a gene expression program (profile) similar to embryonic development and examined the mechanical effects on this program. We used human MSCs cultured in collagen-coated 2D substrates or 3D collagen gels under static and dynamic tension and analyzed the expression of putative markers, including scleraxis, and signaling and ECM molecules implicated in embryonic tenogenesis, such as collagens, Wnts, and MMPs.
Materials and Methods
MSC isolation
Isolation of MSCs is typically based on substrate adherence. 25 Bone marrow-derived MSCs have been used in many of our previous studies, as well as in studies from other laboratories.26–28 Bone marrow was obtained from the femoral heads of adult patients undergoing total joint arthroplasty with the approval of the Institutional Review Board of George Washington University Medical School. The bone marrow was flushed repeatedly with Dulbecco's modified Eagle medium (DMEM), strained through a 40-μm mesh filter, and then centrifuged at 1,000 rpm for 5 min to pellet cells. The cell pellet was resuspended in expansion medium containing DMEM, 100 units/mL penicillin, 100 μg/mL streptomycin, 25 ng/mL Fungizone, and 10% preselected fetal bovine serum (FBS) and plated. Expansion medium was carefully added to the culture after 3 days and completely changed after 6 days. The remaining adherent cells were fibroblastic in morphology and were used as human MSCs (hMSCs). Medium was changed every 3 to 4 days. Cells were subcultured at 85% confluence and frozen at passage 1. After thawing, cells were used at passage 2 or 3. Cell culture reagents were purchased from Invitrogen (Carlsbad, CA).
Static 2D and 3D cultures
To compare the effects of 3D culture with that of 2D culture under static conditions, cells were cultured on flat substrates or in collagen type I gels under uniaxial static tension between two anchors resulting from cell traction exerted on the gel.29,30 For 2D culture, cells were plated at 2 × 104 cells/cm2 on tissue culture plastic (TCP) or collagen type I–coated plates (BD Biosciences, San Jose, CA). For 3D cultures, 8 mL of concentrated (3 mg/mL) collagen type I solution (Vitrogen; Cohesion Technologies, Palo Alto, CA) was mixed with 1 mL of 10× phosphate buffered saline (PBS) and 1 mL of 0.1 M sodium hydroxide. The diluted collagen solution was then neutralized to pH 7.4 with hydrochloric acid. Cells were suspended in the neutralized collagen solution at 2 × 106 cells/mL and cast in wells that had been silicone-coated and outfitted with two fixed-point mounted (1 cm distance) dissecting pins each. Gels were allowed to set for 1 to 2 h at 37°C before being fed with medium.
Mechanical stimulation
A commercial loading system was used to conduct experiments comparing effects of static and cyclic 3D loading. Cells were suspended at 2×106 cells/mL in neutralized collagen type I solution. Linear 3D constructs were fabricated by placing a Tissue Train culture plate (Flexcell International, Hillsborough, NC) into the Flexcell Loading Station with Trough Loaders under vacuum and pipetting 200 μL of the cell suspension into each well of the Tissue Train culture plate. The constructs were allowed to set at 37°C for 1 to 2 h before feeding with medium. Two loading conditions were used. Constructs were cultured under uniaxial static tension, created as a result of MSC-generated cell traction exerted on the anchored collagen gel. Cyclically loaded samples were subjected to a 7-day loading regime of cyclic uniaxial strain at 1 Hz for 30 min/day at 1% elongation beginning 1 day after construct fabrication. Cyclic loading was superimposed on the cell-generated static tension. Statically loaded samples were not subjected to cyclic loading. Loading parameters were chosen based on previous studies that loaded tendon fibroblasts in 3D culture with this system. 31 Constructs were harvested 24 h after 1, 3, 5, and 7 days of cyclic loading for RNA isolation or histological staining.
Construct contraction measurements
Constructs were imaged with a stereoscope before fixation at days 1, 3, 5, and 7. Digital images were acquired using a Polaroid DMC2 digital camera (Polaroid Corporation, Concord, MD) and imported into NIH ImageJ (public domain software available through the National Institutes of Health (Bethesda, MD; available at http://rsb.info.nih.gov/ij/), and the diameter was measured at the narrowest point based on pixel density. Pixel measurements for each day were normalized to day 1 of static loading.
Histology
Constructs were washed with PBS, fixed for 30 min in 10% phosphate buffered formalin (Electron Microscopy Sciences, Hatfield, PA), and washed again with PBS. After fixation, samples were serially dehydrated, cleared with xylene, and paraffin-embedded. Samples were sectioned at 10 μm thickness and stained with picrosirius red for polarized microscopy or at 5 μm thickness for general histological staining. Sections were de-paraffinized in xylene, rehydrated, and stained with picrosirius red, hematoxylin and eosin, or Mallory's trichrome. Slides from different donors were stained together for the same staining time to allow for quantitation of staining intensity.
Light and polarized microscopy
Picrosirius red–stained samples were imaged under polarized light on a Leica DMRB microscope (Leica Microsystems Inc., Bannockburn, IL) equipped with a Q-Color3 digital camera (Olympus, Melville, NY). Histologically stained samples were visualized on a Zeiss Axiovert 135 inverted microscope (Carl Zeiss Microimaging, Inc., Thornwood, NY) equipped with a Polaroid DMC2 digital microscope camera.
Histomorphometry
Histomorphometry was performed on images of trichrome-stained samples. The average blue staining of a sample (indicating collagen content) was quantified using ImageJ. One section from the center of each construct (2 constructs for each of 3 donors) was selected, and microscopic images obtained with a 10× objective at 8 bit depth using a Zeiss Axiovert 135 inverted microscope equipped with a Polaroid DMC2 digital microscope camera. For each image, the total sum of intensity of all blue pixels in the histological section was divided by the total number of pixels in each section. The total number of pixels in a section was determined by drawing a bounding box around the section and counting the number of pixels within. This average measure of trichrome blue staining per section area was then normalized to static conditions of the same day.
Total RNA isolation and reverse transcription
Each construct was homogenized in TRIzol reagent (Invitrogen) and total RNA extracted according to manufacturer's protocol. Extracts were purified with the TURBO DNA-free kit (Ambion, Austin, TX) or Qiagen spin-columns (Qiagen, Valencia, CA). RNA quality was assessed spectrophotometrically on the basis of A260/280. Reverse transcription was performed using Superscript III First-Strand Synthesis System (Invitrogen), with 1.5 μg total RNA reverse transcribed with oligo(dT) in a 20-μL final reaction volume.
Quantitative polymerase chain reaction
Quantitative polymerase chain reaction (qPCR) was performed with iQ SYBR Green Supermix (Bio-Rad, Hercules, CA) on a Bio-Rad iCycler. Primer pairs (Table 1) were designed and optimized for qPCR analysis of expression levels of scleraxis; elastin; biglycan; decorin; tenascin-C; collagen types I, 32 III, XII, and XIV; Wnt4; Wnt5a; Wnt14; Wnt16; Frizzled-5; ß-catenin; and MMP-1, -2, -3, -9, and -13, with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as control. Expression levels relative to GAPDH were calculated using the delta-delta CT method.
Experimental design
Unless otherwise noted, all experiments were performed with cells from three donors (see Table 2).
Effects of 3D culture under static tension
Static culture experiments performed in 2D and 3D were harvested at 11 days of culture for gene expression (n = 3 for each donor, each sample treated individually and subsequently averaged for each donor) and picrosirius red staining (n = 3 for each donor). Cultures to compare substrate material (TCP versus collagen type I–coated plates) were harvested at 3 days of culture for qPCR.
Effects of dynamic loading on 3D cultures
Dynamically loaded samples were loaded for 30 min each day and harvested 24 h after loading for 1, 3, 5, and 7 days. Additional loads were not imposed on statically loaded samples. Constructs were harvested for histology and histomorphometry (n = 2 for each donor) and qPCR (n ≥ 3 for each donor, samples were pooled). Measurements for contraction were taken for samples harvested for histology before sample processing. For all studies, variability between samples (n for each donor) was minimal in comparison with that seen between donors.
Statistical analysis
Means and standard deviations were reported for three donors for all studies unless otherwise noted. Paired Student t-tests were performed as appropriate (when multiple comparisons were not made). For change over time with more than two groups (2 loading conditions × 4 time points), a Fisher least significant difference was performed: an analysis of variance to find effects, followed by multiple t-tests to find the significant changes. All statistical analyses were performed with SigmaStat 3.5 (Systat Software, Inc., Point Richmond, CA).
Results
Effects of 3D culture under static tension
Collagen fibril alignment in 3D
Human MSC–collagen gel constructs were cultured for 7 days between fixed-point anchors under static tension created by endogenous cell traction exerted on surrounding collagen fibrils (Fig. 1A). Polarized microscopy of Picrosirius Red–stained sections revealed birefringence, indicative of matrix alignment along the axis of tension (Fig. 1B). Collagen gels without cells did not contract, and fibrils did not align (results not shown).
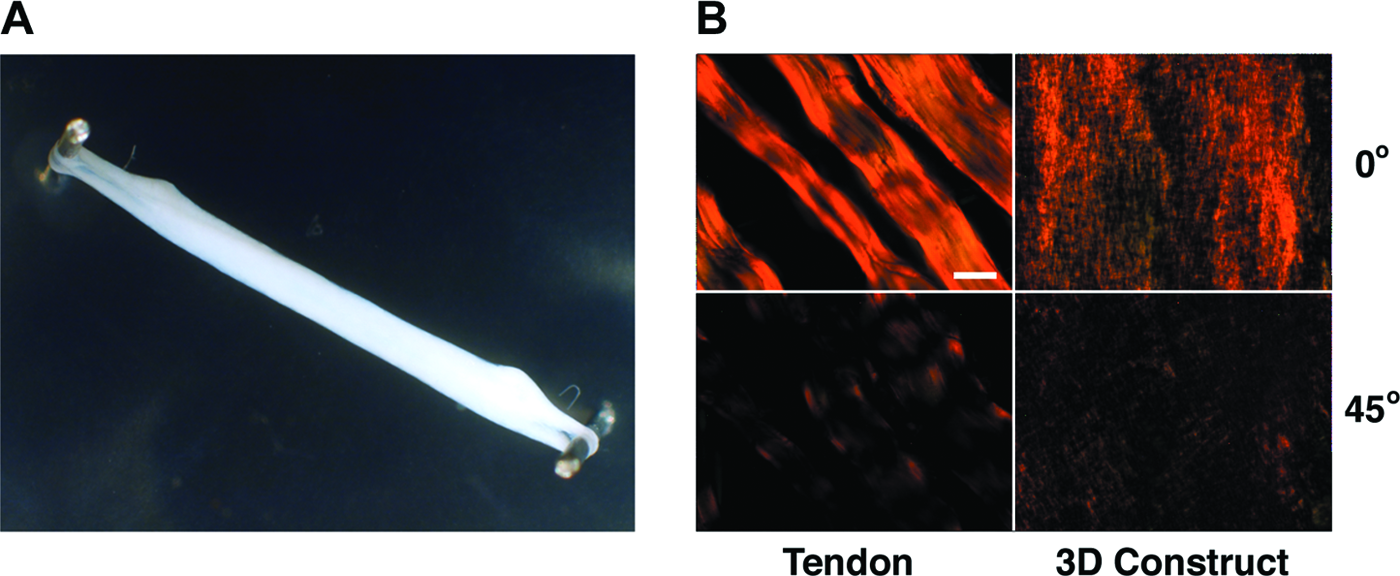
Human mesenchymal stem cells seeded in type I collagen gel and cultured under static tension. (
Gene expression
Gene expression levels were quantified for scleraxis; collagen types I, III, XII, and XIV; biglycan; decorin; elastin; and tenascin-C for hMSCs grown in 2D (TCP) or 3D (collagen gel). Three-dimensional culture in a collagen gel under static tension upregulated scleraxis consistently with all three replicates (Fig. 2), whereas 2D culture did not. Collagen type III was upregulated as well (Fig. 2). Although statistical power was insufficient to determine significance at p = 0.05 because of the limited number of patient samples, a consistent trend of upregulation was observed across all patients. In contrast, collagen types XII and XIV and elastin were downregulated in 3D culture (p < 0.05) (Fig. 2). Other ECM genes, including tenascin-C, decorin, and biglycan, were not regulated in a consistent manner across patients, resulting in no statistically significant differences (Fig. 2). Furthermore, the effect of collagen gel concentration on gene expression showed no consistent trends when hMSCs were cultured in 3D gels of varying collagen type I concentration (1.2 and 2.4 mg/mL) under tension (results not shown).
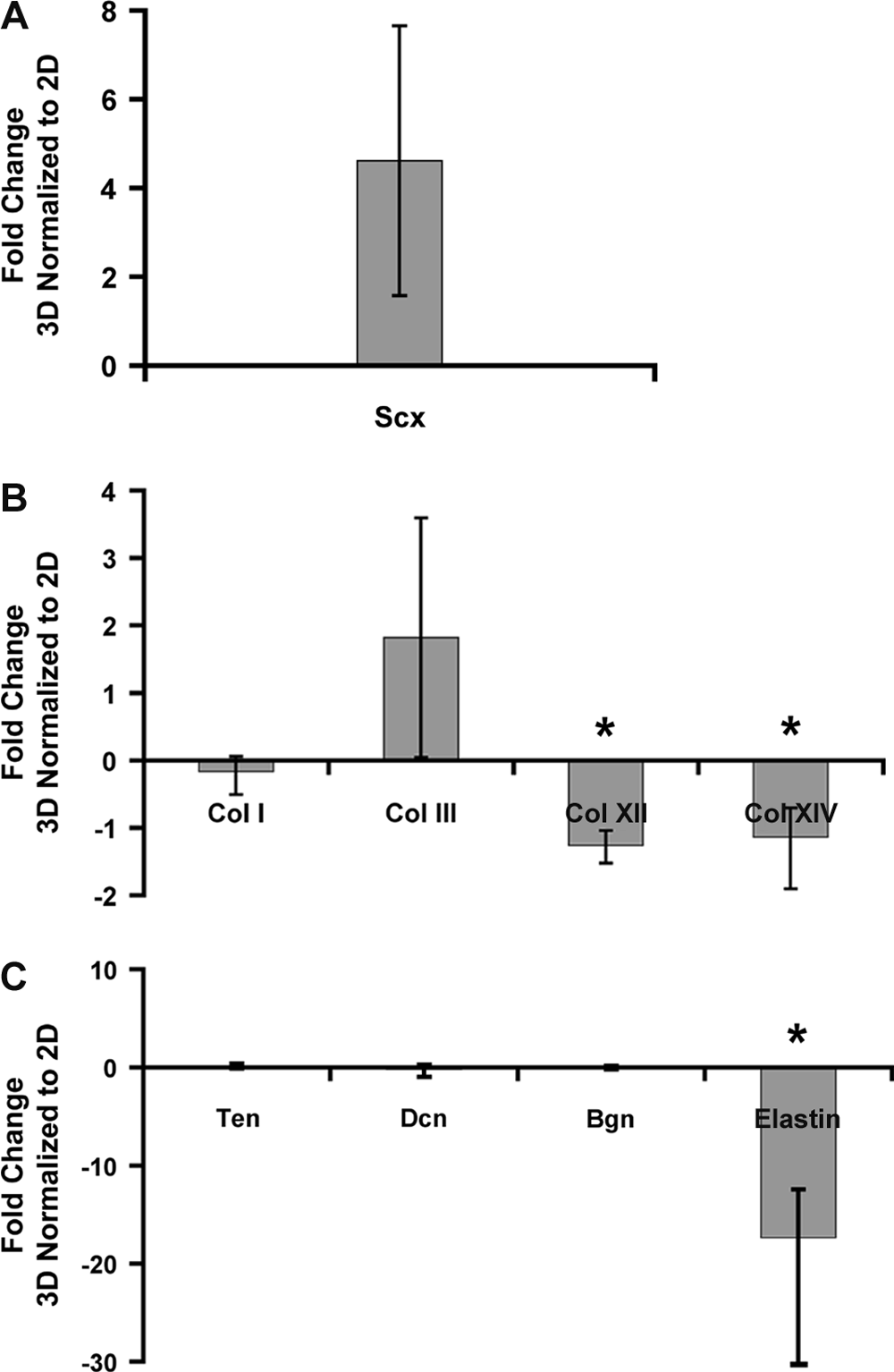
Effects of three-dimensional (3D) and 2D culture conditions on gene expression levels. Quantitative polymerase chain reaction was used to measure transcript levels of (
To examine whether it was the collagen material or the 3D aspect of gel culture that primarily regulated the differences in gene expression between 2D and 3D, hMSCs were cultured in 2D on collagen type I film or TCP. Collagen type I film slightly but significantly upregulated collagen type I gene expression (113 ± 1.6% of TCP control, p < 0.005). In contrast, no other genes examined were consistently regulated in all three patients as a function of substrate material (results not shown). Taken together, these results suggest that the dominant factor in upregulating scleraxis and modulating elastin and collagen types III, XII, and XIV was the 3D culture environment.
Effects of dynamic loading on 3D cultures
Construct contraction
Human MSCs were suspended in collagen type I gels and cast in Tissue Train plates for dynamic mechanical stimulation studies. Human MSC–collagen gel constructs formed cylindrically shaped 3D constructs, attaching to the anchor ends on each side of the well. Under static and cyclic loading conditions, hMSC–collagen gel constructs contracted to form thin constructs, presumably because of cell traction and matrix remodeling (Fig. 3A). To determine contraction rate, measurements of the construct diameter were taken at the mid-section of the construct. Statically and cyclically stretched constructs both contracted at a similar rate (Fig. 3B). No significant differences were found between dynamic and static conditions at day 7 (29 ± 23% and 20 ± 8.0% of the original size, respectively, p = 0.40) (Fig. 3C).
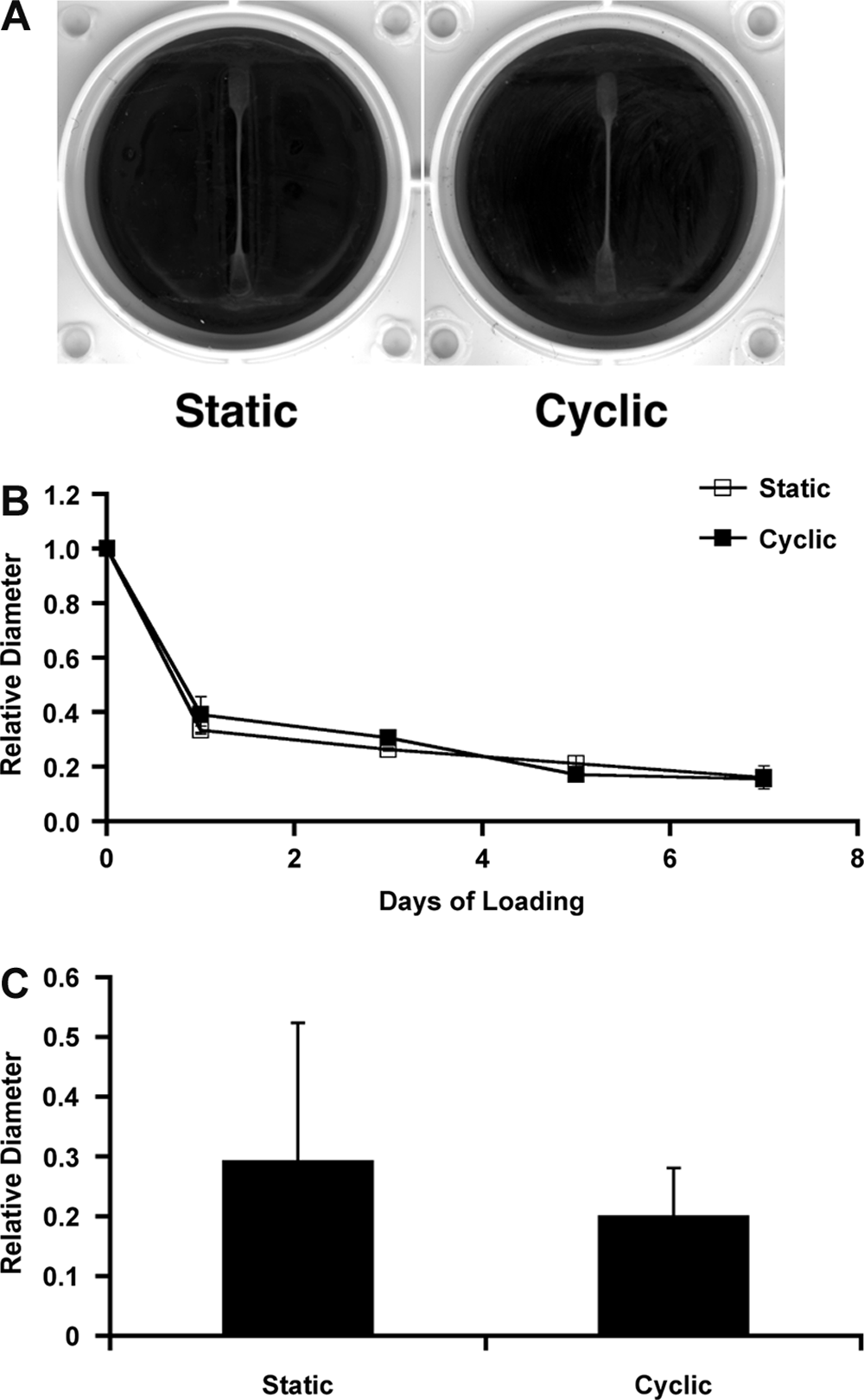
Mesenchymal stem cell–collagen gel constructs loaded for 7 days under static and cyclic conditions. (
Matrix production
Hematoxylin and eosin and trichrome staining indicated neo-tissue formation in statically and cyclically stretched cultures over the 7-day loading period in all three donors. More-intensely eosin-stained matrix was consistently seen in cyclically stretched samples as early as 24 h after only 1 day of loading (Fig. 4A). Trichrome staining indicated greater collagen content (blue) in statically and dynamically loaded constructs with time (Fig. 4B). Correspondingly, cyclically loaded samples displayed greater collagen staining than static controls; histomorphometric measurements indicated 50% more intense blue staining in cyclically loaded samples by day 7 (2 samples per donor of 3 donors, p ≤ 0.05).

Histology of constructs harvested after 1 and 7 days of static and cyclic loading. Longitudinal 5-μm-thick sections histologically stained with (
Gene expression
Gene expression levels were examined as a function of time (all conditions normalized to static conditions on day 1) or of loading (cyclically loaded normalized to statically loaded for each day). To assess the effects of time, day 1 was chosen as the baseline, providing cells time to adjust to culture in the collagen gel.
Scleraxis expression regulated by loading
Gene expression levels of scleraxis were analyzed using qPCR and normalized to day 1 static conditions. Scleraxis expression levels in statically cultured samples decreased with time (68.4 ± 7.7% by day 7, p < 0.05) (Table 3A). In contrast, cyclic loading maintained scleraxis expression over time, with an initial decrease followed by a subsequent increase. Gene expression levels initially decreased (85.6 ± 5.5%, day 1, p < 0.05) but then returned to day 1 levels (95.2 ± 5.5% by day 3, p > 0.10) (Table 3B).
All transcript levels were normalized to GAPDH. Transcript levels expressed relative to static control at day 1. (A) Static Loading, (B) Cyclic Loading. n = 3; * = significant change relative to static control at day 1 with p≤0.05
Gene expression of Wnts and Wnt-related molecules as a function of time
To examine whether dynamic tensile loading regulates tenogenic Wnts, we probed for gene expression of Wnt and Wnt-related molecules (Table 3A, B). Wnt5a was downregulated with time under static and dynamic conditions (p < 0.05 as early as day 3). Cyclic loading downregulated Wnt14 mRNA levels with time (p < 0.05 by day 5) while maintained under static conditions (p > 0.10). In contrast, Wnt16 mRNA levels increased significantly after 3 days of cyclic loading (p < 0.05), followed by a return to day 1 levels. Frizzled-5 also varied with time in response to cyclic loading, with significantly lower mRNA levels after 1 day (p < 0.05), then higher levels at 3, 5, and 7 days (p < 0.01, p < 0.001, p < 0.05, respectively). Expression levels for Frizzled-5 under static conditions had also increased significantly at 3 days (p = 0.005). Transcript levels for ß-catenin were consistently upregulated after 1 day of loading under dynamic conditions for days 3, 5, and 7 (p < 0.05, p = 0.002, p = 0.005, respectively).
Gene expression of Wnts and Wnt-related molecules as a function of loading
Wnt4 and Frizzled-5 were both significantly downregulated (29% and 44% decrease, respectively, p < 0.05) after the first day of cyclic loading, while Wnt5a demonstrated a 0.2-fold increase in expression (p < 0.05) after loading for 3 days (Fig. 5A, B).
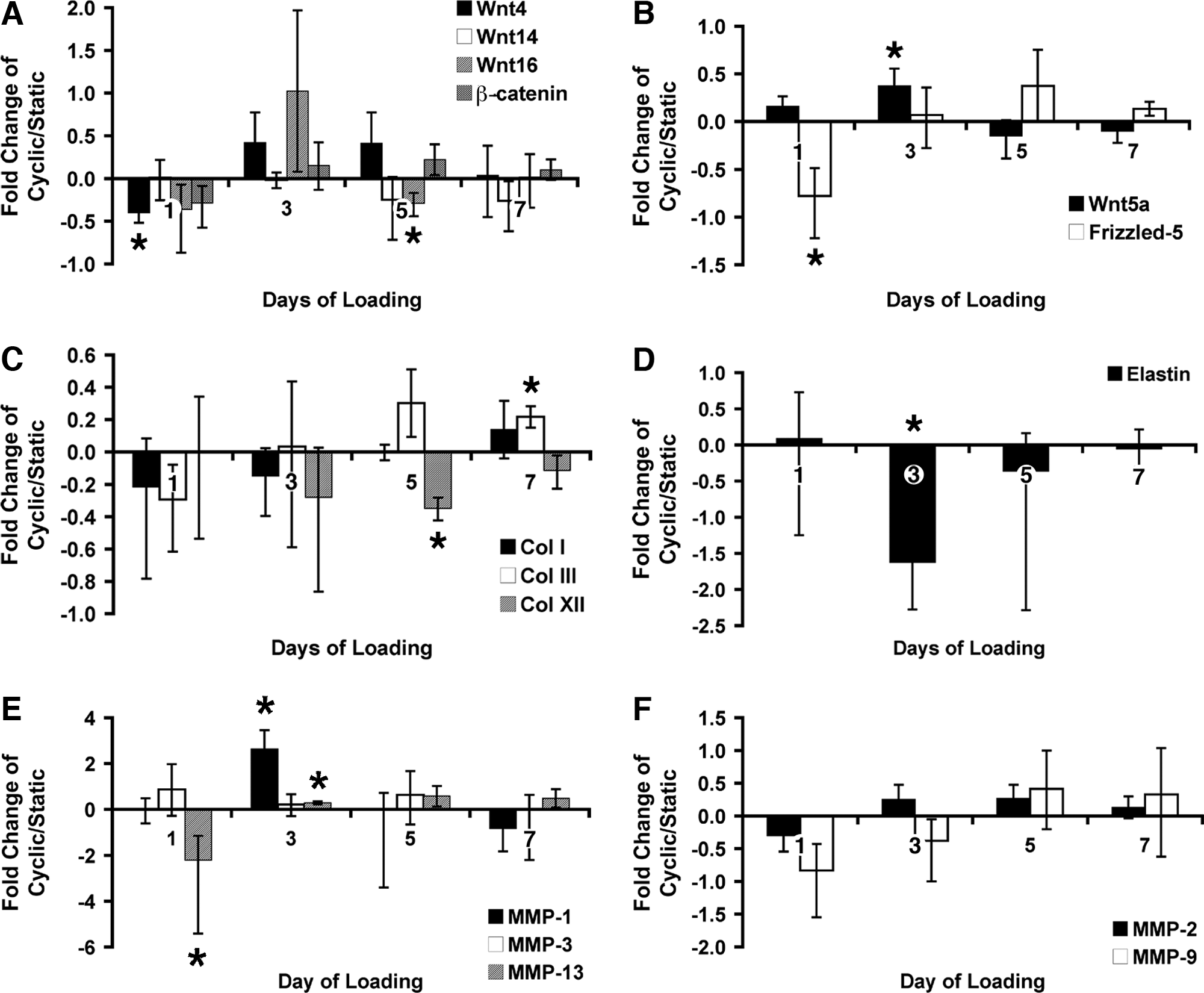
Gene expression as a function of static or cyclic load. All transcript levels were normalized to glyceraldehyde 3-phosphate dehydrogenase. Proportional fold change in transcript levels in cyclically loaded constructs is expressed relative to static control according to culture day. Increases are plotted as [Fold Change – 1], and decreases are plotted as [1/(Fold Change) – 1]. (
Gene expression of matrix molecules and MMPs as a function of time
To determine whether the differences seen in matrix accumulation as detected using histological staining might be due to matrix synthesis, active remodeling of the matrix, or both, we examined gene expression of matrix molecules and MMPs (Table 3A, B). Under static and dynamic conditions, transcript levels for collagen types I, III, and XII increased with time (p ≤ 0.06 by day 7). Gene expression levels of elastin increased with time under both loading conditions as well but at a slightly greater rate under static conditions. Under static and dynamic conditions, MMP-1 and -3 mRNA levels decreased with time (p ≤ 0.05 as early as day 3 and p ≤ 0.02 by day 7), whereas gene expression levels for MMP-13 increased with cyclic loading after 3 days of loading (p < 0.001) before decreasing for the remaining time-points. MMP-2 mRNA levels increased after 3 days of cyclic loading and maintained higher levels for the remaining time points (p < 0.05). MMP-9 mRNA levels remained constant over the 7 days of loading.
Gene expression of matrix molecules and MMPs as a function of loading
Minimal but statistically significant differences were detected between dynamic loading and static conditions on day 5 for collagen type XII (26% decrease, p ≤ 0.01) and on day 7 for collagen type III (0.22-fold increase, p < 0.05) (Fig. 5C). Elastin mRNA levels were significantly higher for constructs cultured under static conditions than under dynamic conditions at day 3 (1.62-fold increase, p < 0.05) (Fig. 5D). These results suggest that cyclic loading regulated matrix synthesis. However, MMPs were also significantly regulated under dynamic loading conditions. Under dynamic conditions, MMP-1 and -13 were upregulated significantly more (2.6-fold increase, p = 0.01, and 0.3-fold increase, p < 0.05, respectively) than under static conditions after 3 days of loading, whereas MMP-9 and MMP-13 were downregulated 44% (p < 0.07) and 69% (p < 0.05), respectively, after just 1 day of loading (Fig. 5E, F). MMP-1 mRNA levels decreased (p = 0.08) after 7 days of cyclic loading. Taken together, these results suggest that active matrix remodeling activity accompanied increased matrix accumulation as detected using histochemical staining.
Discussion
This work investigated the biological influence of mechanical load and spatial culture environment on MSC tenogenesis with respect to known embryonic differentiation marker expression and histological outcomes. Studies using MSC-seeded collagen gels to regenerate tendon and ligament replacements have focused on functional measures (e.g., matrix stiffness, cell and matrix alignment, and matrix composition—primarily collagen types I and III). In contrast, we studied expression of markers correlated with embryonic tenogenesis (e.g., scleraxis and Wnt signaling molecules) and functional markers (histological measures) in MSCs subjected to a tenogenic environment (3D gel under tension).
We chose to use a collagen type I gel as the scaffold for 3D cultures for several reasons. First, this model is reasonably well characterized, and thus experiments performed using this scaffold would allow us to compare our data with those from previously reported results. Second, fabrication of cell-seeded scaffolds is easily repeatable to achieve a large number of replicate constructs. Third, there are readily available methods of applying mechanical stimulation to collagen gel constructs. The Flexercell Strain Unit used with Tissue Train culture plates has been demonstrated to maintain the proper phenotype of tendon fibroblasts when seeded in collagen gels and cyclically loaded under uniaxial tension. 31 Finally, we chose collagen type I gel for our scaffold because collagen type I is the primary matrix component of mature tendon and ligament. Other matrix molecules found in tendon and ligament include smaller, but significant, amounts of collagen types III, XII, and XIV; elastin; and proteoglycans.1–3,33
Our results support a role for scleraxis in tenogenesis of MSCs. Scleraxis has been shown to be a unique developmental marker for tendon and ligament during avian and murine embryogenesis,14,15 because it is expressed in the earliest tendon progenitor cells and is continuously expressed throughout development. Disruption of normal tendon development in scleraxis-null mice suggests that scleraxis function is required for normal tendon formation. 16 In our studies, scleraxis was consistently upregulated more in 3D culture than in 2D culture, and its expression was maintained with dynamic stimulation, supporting the hypothesis that tensile forces will induce tenogenesis of MSCs and that dynamic stimulation is necessary to maintain the tenogenic differentiation process. Based on these results, together with matrix gene expression, we focused on 3D cultures for the loading studies.
Wnt4, Wnt14, and Wnt16 have been implicated in tendon development during embryogenesis. During joint formation in the chick embryo, Wnt14 mRNA expression is seen in the developing tendons and soft tissue–derived structures, suggesting a distinct role for this molecule in tendon morphogenesis. 23 In the developing joint of the mouse embryo, Wnt4, Wnt14, and Wnt16 are expressed in non-cartilaginous mesenchymal cells that will form fibrous tissues including ligaments and tendons 22 and have been demonstrated to signal through the canonical Wnt pathway involving ß-catenin. 22 Wnt14 and ß-catenin have been shown to inhibit chondrogenesis.22,23 Although several studies have implicated Wnts in bone and cartilage formation,34–41 there are no functional studies of Wnts in tenogenesis of MSCs. We hypothesized that Wnt4, Wnt 14, Wnt16, and ß-catenin are involved in tenogenesis of MSCs and therefore would be candidate marker genes for tenogenesis of MSCs. We examined the regulation of Wnt4, Wnt14, Wnt16, and ß-catenin transcripts in response to dynamic loading. Wnt5a and its commonly paired receptor Frizzled-5 were included to check for the potential involvement of non-canonical Wnt signaling in tenogenesis. Wnt4 and Frizzled-5 mRNA levels decreased more after 1 day in response to cyclic loading than in static controls. In contrast, Wnt5a increased after 3 days in response to cyclic loading. These opposing trends in gene expression of Wnt4 and Wnt5a were interesting, because others have hypothesized that Wnt4 and Wnt5a may have disparate effects on differentiation of chondrocytes. Wnt4 inhibits initiation of chondrogenesis and in chondrogenic cultures signals through the canonical Wnt pathway to accelerate terminal differentiation.36,38 In contrast, Wnt5a is necessary for chondrogenesis41,42 and in chondrogenic cultures signals through the non-canonical Wnt/Ca2+ pathway to delay chondrocyte maturation.36,38,41,43 In our studies, the downregulation of Wnt4 coupled with the upregulation of Wnt5a detected under dynamic conditions suggested a parallel Wnt4/Wnt5a relationship during tenogenesis of MSCs.
Altman and colleagues demonstrated the potential for 3D MSC–collagen constructs to produce ligament replacements in response to mechanical stimulation. The MSCs expressed significantly higher mRNA levels of collagen types I and III after 14 days of cyclic loading and stained positively for collagen types I and III after 21 days. 12 There was no significant difference in collagen type I and III expression levels before 14 days. Similarly, in our study, there was minimal differential regulation of gene expression levels for collagen types I, III, and XII with cyclic loading for up to 7 days. We observed a significant increase in matrix staining in cyclically loaded constructs for all time points, even at the earliest time point. The possibility of greater staining intensity due to an increase in compaction of gel-derived collagen fibrils was ruled out because dynamic and static conditions resulted in similar contraction rates. The gene expression analysis indicated that the greater matrix staining was not due to greater expression of matrix markers. We hypothesized that mechanically regulated post-translational mechanisms, such as matrix remodeling, are involved in greater matrix deposition or assembly. To explore this possibility, we looked for differential regulation of mRNA levels of MMPs.
MMPs are enzymes that are involved in a variety of normal and pathological conditions that involve matrix degradation and remodeling, including embryonic development, morphogenesis, reproduction, and tissue remodeling. Collagenases, which include MMP-1 and -13, have the ability to cleave native collagen fibrils to produce denatured collagen. In turn, the breakdown of denatured collagen occurs through gelatinases, such as MMP-2 and -9. MMP-3, commonly referred to as stromelysin, has the capacity to degrade a range of substrates, including non-collagenous matrix molecules. In mature tendons and ligaments, our knowledge of the involvement of MMPs in normal tissue maintenance is gleaned largely from in vitro mechanical stimulation studies. MMP-1 and -3 expression levels increased in rabbit and human tendon fibroblasts subjected to cyclic strains and fluid-induced shear stress in vitro44–47 without corresponding changes in collagen type I expression. 44 In contrast, uniaxial cyclic loading downregulated MMP-1 in rat-tail tendon explants, 48 whereas rat tenocytes subjected to shear stress demonstrated upregulation of MMPs accompanied by downregulated expression of collagen types I and III. 49 In humans, pro-MMP levels were measured in vivo in Achilles tendons after 1 h of physical exercise and found to vary throughout recovery time. 50 Although not conclusive, these studies suggest that tissue maintenance via active matrix remodeling activity occurs in response to movement or exercise within physiological strains. The role of MMPs in embryonic tendon development or tenogenesis of adult MSCs has yet to be elucidated. In our studies, cyclic loading differentially regulated MMP mRNA levels by decreasing MMP-9 and MMP-13 expression after 1 day of loading, increasing MMP-1 and -13 after 3 days, and decreasing MMP-1 after 7 days. Based on these results, we propose that dynamic loading enhances tenogenesis in part by promoting active remodeling of the collagen matrix.
Our results demonstrate that scleraxis, Wnts, and MMPs are regulated during tenogenic differentiation of hMSCs in vitro. We speculate that these putative tendon markers may be regulating neo-tissue formation, based on changes in gene expression levels and correlating histological staining. Our results provide important information regarding the ability of mechanically defined 3D cultures to control transcriptional changes. However, changes in mRNA levels may not correlate with protein levels, because post-transcriptional or post-translational modification of mRNA or protein, respectively, can occur. Future studies should analyze the presence of proteins. For instance, immunohistochemical staining of the construct will define the specific collagen types in the newly deposited matrix and can provide further information regarding which Wnts or MMPs are acting upon the cells or matrix within the construct. An additional layer of complexity exists in determining the total amount of MMP protein present (e.g., via immunological methods or zymography), because these enzymes may be present in the inactive pro-form or in complexes with inhibitors such as tissue inhibitors of MMPs. Verification of net enzyme activity would provide more definitive evidence of remodeling activity due to MMPs.
In summary, a 3D environment and mechanical loading potentiate in vitro expression of embryonic tenogenic markers in MSCs. Short-term periods of stimulation (7 days) for only 30 min/day were sufficient to induce significant changes at the mRNA and the post-translational level in 3D MSC cultures. Based on our results, we propose that scleraxis can be used as an early marker of tenogenesis of MSCs in vitro. Three-dimensional culture under static tension upregulated scleraxis in human MSCs, but cyclic loading was required to maintain scleraxis expression with time, demonstrating the need for dynamic mechanical stimulation for tenogenic differentiation of MSCs. Greater staining for matrix coupled with differential regulation of MMPs at a transcriptional level and minimal changes in collagen mRNA levels suggested that enhanced tendinous neo-tissue development was largely due to changes in matrix deposition and remodeling activity regulated by mechanical loading conditions. This is the first report to demonstrate the potential involvement of matrix remodeling during tenogenesis of human MSCs in a dynamic, 3D tissue-engineering model. Finally, we showed a possible Wnt4/Wnt5a relationship in tenogenesis parallel to the one that has been reported in the cartilage system. To our knowledge, this is the first report to investigate the possible involvement of Wnt regulation in tenogenic differentiation of MSCs.
Footnotes
Acknowledgments
The authors are grateful to Dr. Juan M. Taboas (National Institutes of Health) for his invaluable consultation with statistical analysis and to Dr. Paul Manner (George Washington University) for providing the human bone marrow samples. The authors would also like to acknowledge the National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS) Light Imaging Section for use of the microscopy facilities. This research was supported by the Intramural Research Program of the NIH, NIAMS (Z01 AR41131).