
Abstract
Adipose tissue–derived mesenchymal stem cells (AT-MSCs) in combination with bone morphogenetic protein-2 (BMP-2) or transforming growth factor-β1 (TGF-β1) are under evaluation for bone tissue engineering. Posttranslational modification of type I collagen is essential for functional bone tissue with adequate physical and mechanical properties. We investigated whether BMP-2 (10–100 ng/mL) and/or TGF-β1 (1–10 ng/mL) affect gene expression of α2(I) procollagen and collagen-modifying enzymes, that is, lysyl oxidase and lysyl hydroxylases 1, 2, and 3 (encoded by PLOD1, 2, and 3), by human AT-MSCs. BMP-2, but not TGF-β1, increased alkaline phosphatase activity after 28 days, indicating osteogenic differentiation of AT-MSCs. At day 4, both BMP-2 and TGF-β1 upregulated α2(I) procollagen and PLOD1, which was downregulated at day 28. TGF-β1, but not BMP-2, downregulated PLOD3 at day 28. Lysyl oxidase was upregulated by TGF-β1 at day 4 and by BMP-2 at day 7. Neither BMP-2 nor TGF-β1 affected PLOD2. In conclusion, these results suggest that AT-MSCs differentially respond to BMP-2 and TGF-β1 with changes in gene expression of collagen-modifying enzymes. AT-MSCs may thus be able to appropriately modify type I collagen to form a functional bone extracellular matrix for tissue engineering, dependent on the growth factor added.
Introduction
Bone tissue engineering requires cells that are able to generate bone extracellular matrix to provide the mechanical properties allowing proper bone functioning. The principal components of bone extracellular matrix are anorganic hydroxyapatite and organic collagen, mainly type I collagen. 11 Fibrillar type I collagen is the major type of collagen present in bone extracellular matrix, and responsible for the elasticity of bone tissue. 11 Type I collagen is synthesized by the bone-forming osteoblasts, and is composed of a triple helix of two α1(I) procollagen chains (COL1A1) and one α2(I) procollagen chain (COL1A2). 12 The procollagen chains are posttranslationally modified to promote the fibrillar supramolecular structure of collagen that is responsible for the mechanical resilience of the tissue. 12 Hydroxylation of the proline and lysine residues is catalyzed by prolyl hydroxylases and lysyl hydroxylases, respectively. 12 Hydroxyproline (Hyp) is essential for intramolecular hydrogen bonds within the triple helical domains. 12 Specific hydroxylysine (Hyl) residues serve as attachment sites for galactose or glucosylgalactose, and are involved in the intermolecular cross-linking process, providing collagen with its tensile strength. 13 Stable, mature cross-links of hydroxylysyl pyridinoline (HP) and lysyl pyridinoline (LP) can be formed in between separate collagen triple helices in the extracellular matrix after lysyl oxidase (LOX) has catalyzed the formation of aldehydes of the lysine or Hyl residues in the telopeptides. 12 These cross-links are a prerequisite for the physical and mechanical properties of collagen fibrils and for the formation of a stable collagen network (reviewed by Gelse et al. 12 ).
The lysyl hydroxylase family of enzymes consists of lysyl hydroxylase 1, 2, and 3.14–17 Lysyl hydroxylases 1 and 3 hydroxylate lysine residues within the triple helical region of the collagen molecule. 18 In addition, lysyl hydroxylase 3 has been shown to exhibit collagen glucosyltransferase and galactosyltransferase activity.16,19,20 Lysyl hydroxylase 2 is held responsible for hydroxylation of telopeptidal lysine residues situated at the terminal ends of the collagen triple helix. 21 Lysyl hydroxylases 1, 2, and 3 are encoded by procollagen-lysine,2-oxoglutatarate 5-dioxygenase 1 (PLOD1), PLOD2, and PLOD3 genes, respectively.14–17,22 Mutations in these genes are responsible for defective collagen assembly and result in human pathologies; mutations in PLOD1 or PLOD2 result in Ehlers-Danlos syndrome type VI or Bruck syndrome.21,23–25
Members of the transforming growth factor-β (TGF-β) superfamily are currently under investigation for bone tissue engineering applications.8,26–28 These growth factors play a key role in skeletogenesis, that is, during endochondral and intramembranous bone formation.29,30 Especially, BMP-2 and TGF-β1 are under evaluation for use in bone tissue engineering.8,27 Both BMP-2 and TGF-β1 are present in the bone extracellular matrix, where TGF-β1 is the most abundant. Unlike BMP-2, TGF-β1 will not produce bone when injected into ectopic sites.31–33 Both growth factors induce differentiation of stem cells along the osteogenic lineage, and stimulate type I collagen production by the cells.8,26,34 BMP-2 has been shown to affect gene expression of collagen-modifying enzymes in the murine promyoblastic cell line C2C12 and the Hyl content in the matrix produced by C2C12 cells. 35
Posttranslational modification of collagen is a prerequisite for the development of functional bone tissue, as desired in bone tissue engineering. This has not yet been studied in human AT-MSCs in response to BMP-2 or TGF-β1 treatment. We hypothesize that AT-MSCs are able to posttranslationally modify type I collagen and thereby promote formation of an appropriate functional extracellular matrix. To address this issue, we have assessed gene expression of COL1A2, and collagen-modifying enzymes LOX and lysyl hydroxylases 1, 2, and 3 (PLOD1, PLOD2, and PLOD3) by AT-MSCs. The Hyl content, the amount of collagen produced by human AT-MSCs, and the amount of mature trifunctional collagen cross-links HP and LP in the extracellular matrix was determined as well. Alkaline phosphatase (ALP) activity and gene expression of runx2 and OPN were used as indicators of osteogenic differentiation of AT-MSCs. Gene expression of aggrecan (AGG), the predominant large chondroitin sulfate proteoglycan in cartilage, and SOX9 and SOX5, transcription factors crucial for chondrogenic differentiation, were used to study whether AT-MSCs undergo chondrogenic differentiation after treatment with BMP-2 or TGF-β1.36,37 Peroxisome proliferator–activated receptor γ (pparG) is crucially involved in adipogenesis and was therefore studied as indicator for adipogenesis in AT-MSCs in response to BMP-2 or TGF-β1. 36
Materials and Methods
Donors
Human subcutaneous adipose tissue samples were obtained as waste material after elective surgery and donated upon informed consent of the patients by surgery departments of clinics in The Netherlands. Adipose tissue was taken from three female patients (age: 16, 36, and 37 years). The Ethics Review Board of the Vrije Universiteit approved the protocol.
AT-MSC isolation and storage
Human adipose tissue was harvested by tumescent liposuction or resection. AT-MSCs were isolated from the lipoaspirate or the resection material as described earlier with minor modifications.4–6,8,38 Briefly, the harvested tissue was enzymatically digested for 45 min at 37°C with 0.1% collagenase A (Roche Diagnostics, Mannheim, Germany) in phosphate-buffered saline (PBS) containing 1% bovine serum albumin (BSA; Sigma-Aldrich, St. Louis, MO) under intermittent shaking, washed with Dulbecco's modified Eagle's Medium–glucose (DMEM-glucose; Bio Whittaker, Cambrex, Verviers, Belgium) containing 10% fetal bovine serum (FBS; Gibco, Paisley, UK), and centrifuged for 10 min at 600 g. The resulting cell pellet containing the AT-MSCs was resuspended in PBS, filtered through a 200 μm mesh (Brau/Beldico s.a-n.v, Marche-en-Famenne, Belgium) to remove debris, and subjected to Ficoll density centrifugation (lymphoprep, ρ, 1.077 g/mL; osmolarity, 280 ± 15 mOsm; Axis-Shield, Oslo, Norway) to remove remaining erythrocytes. The interface containing the AT-MSCs was collected and washed with DMEM containing 10% FBS. Three to 10 × 106 cells were resuspended in a mixture (1:1) of DMEM with 10% FBS and cyroprotective medium (Bio Whittaker), frozen under controlled rate conditions in a Kryosave (HCI Cryogenics BV, Hedel, The Netherlands), and stored in the vapor phase of liquid nitrogen.
AT-MSC culture
Cryopreserved AT-MSCs containing cell suspensions were thawed and seeded at 4–10 × 104 cells/cm2 in culture medium, which was composed of DMEM containing 10% FBS (Gibco) supplemented with 500 μg/mL streptomycin sulfate (Sigma-Aldrich), 600 μg/mL penicillin (Sigma-Aldrich), and 2.5 μg/mL amphotericin B (Fungizone; Gibco). AT-MSCs were incubated at 37°C under 5% CO2 in a humidified atmosphere, and medium was exchanged every 3–4 days. The culture period right after isolation is defined as passage 0. Upon confluence, AT-MSCs were harvested using 0.25% trypsin and 0.1% EDTA in PBS, seeded in 75 cm2 culture flasks (Greiner Bio-One, Kremsmuenster, Austria), and cultured until the cell layer reached confluence again (passage 1). AT-MSCs used in this study were used at passage 2 or 3.
AT-MSC characterization
Human AT-MSCs at passage 1 were analyzed for the expression of the mesenchymal stem cell surface markers CD105/endoglin and CD166/ALCAM by fluorescence-activated cell sorting (FACS) as described earlier.6,8,39,40 Briefly, cells were incubated with either phycoerythrin-labeled anti-CD166/ALCAM (BD Biosciences–Pharmingen, San Diego, CA) or with anti-CD105/endoglin monoclonal antibodies (Abcam, Cambridge, UK) and with a secondary fluoroisothiocyanine-conjugated antibody against CD105/Endoglin (Biotrend, Cologne, Germany), and expression of CD105/endoglin and CD166/ALCAM was assessed with a FACSScan (BD Biosciences–Pharmingen). Approximately 58% of human AT-MSCs at passage 1 were positive for the mesenchymal stem cell marker CD105/endoglin, and 59% were positive for the mesenchymal stem cell marker CD166/ALCAM. 39 More than 60% of the AT-MSC population showed the marker profile CD31− CD34+ CD45− CD90+ CD105+ CD166+ as described earlier. 38
BMP-2 and TGF-β1
Recombinant human BMP-2 (Peprotech EC, London, UK) was reconstituted at 100 μg/mL in PBS containing 0.1% BSA. Recombinant human TGF-β1 (Sigma-Aldrich) was reconstituted at 2 μg/mL in 4 mM HCl containing 0.1% BSA (Sigma-Aldrich). AT-MSCs were treated with either BMP-2 at 10 and 100 ng/mL or 1 and 10 ng/mL TGF-β1 for 4, 7, and 28 days. AT-MSCs that were harvested after 4 and 7 days of treatment with BMP-2 or TGF-β1 were seeded at 104 cells/cm2, and AT-MSCs harvested after 28 days of growth factor treatment at 5 × 103 cells/cm2. Cells were cultured in DMEM supplemented with 10% FBS, antibiotics, amphotericin B, 10 mM β-glycerophosphate (Sigma-Aldrich), 50 μg/mL ascorbic acid (Merck, Darmstadt, Germany), and either 10 or 100 ng/mL BMP-2, or 1 or 10 ng/mL TGF-β1 for 4, 7, and 28 days. Control cultures were cultured in medium without BMP-2 or TGF-β1.
Alkaline phosphatase
ALP activity was measured to assess the osteoblastic phenotype of AT-MSCs after 28 days of treatment with BMP-2 and TGF-β1. ALP activity and protein content were determined in the cell layer. ALP activity was measured in the cell lysate using p-nitrophenyl phosphate (Merck) as a substrate at pH 10.3, according to the method described by Lowry. 41 The absorbance was read at 410 nm with a microplate reader (BioRad Laboratories, Hercules, CA). The amount of protein at days 4, 7, and 28 in the AT-MSC layer was measured using a BCA protein assay reagent kit (Pierce, Rockford, IL), and the absorbance was read at 570 nm with a microplate reader. ALP activity values were expressed per amount of protein in the AT-MSC layer.
Gene expression
Total RNA was isolated from AT-MSCs after 4, 7, and 28 days of treatment with or without BMP-2 or TGF-β1 using Trizol® (Invitrogen, Carlsbad, CA), according to the manufacturer's instructions. cDNA synthesis was performed using 0.5–1 μg total RNA in a 20 μL reaction mix consisting of 5 units Transcriptor Reverse Transcriptase (Roche Diagnostics), 0.08 A260 units random primers (Roche Diagnostics), 1 mM of each dNTP (Invitrogen), and 1× concentrated Transcriptor RT reaction buffer (Roche Diagnostics).
For the determination of relative gene expression of osteogenic markers, cDNA was amplified using specific primers for 18S as housekeeping gene, and the target genes RUNX2, OPN, osteocalcin (OC), AGG, SOX9, SOX5, and pparg. The primer sequences are listed in Table 1. Real-time PCR reactions were performed using the LightCycler 480 SYBR green I Master reaction mix according to the manufacturer's instructions (Roche Diagnostics) in a LightCycler 480 (Roche Diagnostics). Values of target gene expression were normalized for 18S gene expression.
FAM, 6-carboxyfluorescein; DABCYL, 4-(4-dimethylaminophenyl) benzoic acid.
For the determination of gene expression of COL1A2 and collagen-modifying enzymes, cDNA was amplified using specific primers and specific molecular beacon probes in a total reaction volume of 12.5 μL, containing 1× PCR buffer (Applied Biosystems), 400 μM of each dNTP (Roche Diagnostics), 3.5 mM Mg2+ (Applied Biosystems), 500 nM of each specific primer, 200 nM of specific molecular beacon probe, and 0.5 unit Amplitaq Gold polymerase (Applied Biosystems). Real-time PCR for determination of COL1A2, PLOD1, PLOD2, and PLOD3 gene expression was performed in an ABI PRISM® 7700 Sequence Detection System (Applied Biosystems) and consisted of a 5 min interval at 95°C followed by 40 cycles of 95°C for 30 s, 56°C for 40 s, and 72°C for 30 s. Primer sequences and the corresponding molecular beacon probe sequences for COL1A2, PLOD1, PLOD2, and PLOD3 are listed in Table 1. TaqMan® gene expression assays were used according to the manufacturer's protocol for the determination of 18S (Hs99999901_s1; Applied Biosystems) and LOX (Hs00184700_m1; Applied Biosystems) gene expression in an ABI PRISM® 7700 Sequence Detection System (Applied Biosystems). Data were analyzed using sequence detector V1.7 software (Applied Biosystems) and normalized for 18S or COL1A2 gene expression.
Cross-link and amino acid analysis
Measurements were performed on extracellular matrix produced by AT-MSCs after 28 of treatment with or without the growth factors BMP-2 or TGF-β1. At day 28, the deposited matrix was washed with PBS, and the cells present in the matrix were killed with two freeze–thaw cycles. The remaining matrix was incubated in PBS at 37°C for another week. During the latter incubation, the difunctional cross-links did further mature into the trifunctional pyridinoline cross-links. The level of pyridinoline cross-links per collagen molecule after the postincubation period reflects more closely the in vivo situation than without the postincubation period. 42 Hyp and Hyl levels were determined in acid hydrolysates of matrix samples after derivatization with 9-flourenylmethyl chloroformate by reverse-phase high-performance liquid chromatography. 43 HP and LP cross-links were measured in the same acid hydrolysates as described previously.44,45 Collagen content of the extracellular matrix produced by human AT-MSCs was calculated assuming 300 Hyp residues per collagen triple helix. Pyridinolines and Hyl levels were expressed as total amount of residues per collagen molecule (triple helix).
Statistics
Data were obtained using AT-MSCs derived from three donors (n = 3). Each data set was treated either with BMP-2 (10 or 100 ng/mL) or TGF-β1 (1 or 10 ng/mL), or not treated with growth factors (control). Cells from each donor were harvested after 4, 7, and 28 days of treatment with growth factors, or left untreated and harvested at the same time points. Treatment modalities were compared to within patient control modalities, or treatment-over-control (T/C) ratios were calculated. To obtain normally distributed data, values were log-transformed. ALP activity (normalized for protein content) and COL1A2 gene expression (normalized for 18S) data are expressed as mean ± SEM. Target gene expression (runx2, OPN, sox9, AGG, PLOD1–3, and LOX) was normalized for 18S, and genes encoding collagen-modifying enzymes were also normalized for COL1A2 gene expression and expressed as mean ± SEM of growth factor T/C ratios. Because every patient represents one experimental unit, we used a paired t-test to evaluate statistically significant differences between control and treatment modalities for ALP activity and COL1A2 gene expression. T/C ratios were tested with a t-test for single group mean and compared to 0 (log T/C = 0, no effect) to observe significant increases or decreases in gene expression. To explore differences between growth factor treated and controls at each time point separately, we chose for a planned comparison as analytic strategy. Therefore, we have used a one-tailed t-test, and differences were considered significant if p < 0.05.
Results
The protein content of AT-MSC cultures increased during the 28-day culture period. Neither BMP-2 nor TGF-β1 affected the total amount of protein compared to untreated controls at all time points (Fig. 1).

Effect of BMP-2 (10 and 100 ng/mL) or TGF-β1 (1 and 10 ng/mL) on protein content of AT-MSCs. (
We studied gene expression of RUNX2 and OPN as indicators of osteogenic differentiation, and SOX5, SOX9, and AGG gene expression as indicator of chondrogenic differentiation of human AT-MSCs in response to BMP-2 or TGF-β1. A trend in upregulation of RUNX2 gene expression by BMP-2 and TGF-β1 was observed at day 4 (Fig. 2A). TGF-β1 (10 ng/mL) significantly increased RUNX2 gene expression at days 7 and 28, whereas BMP-2 (100 ng/mL) decreased RUNX2 gene expression at day 28 (Fig. 2A). TGF-β1 (10 ng/mL) increased OPN gene expression at day 4; BMP-2 (10 ng/mL) seemed to increase (not significant) OPN gene expression at day 4 (Fig. 2B). OC gene expression was detectable, but not quantifiable. SOX9 gene expression was not effected by BMP-2 (Fig. 2C). TGF-β1 (10 ng/mL) increased SOX9 gene expression by 4.4-fold at day 4 (Fig. 2C), which is not accompanied by an increase in AGG gene expression at later time points (Fig. 2D). TGF-β1 (10 ng/mL) decreased AGG gene expression by 4.5-fold at day 4, whereas BMP-2 had no effect. BMP-2 and TGF-β1 had no effect on AGG gene expression at days 7 and 28 (Fig. 2D). SOX5 gene expression was detectable, but not quantifiable. Gene expression of PPARG was not affected (data not shown).
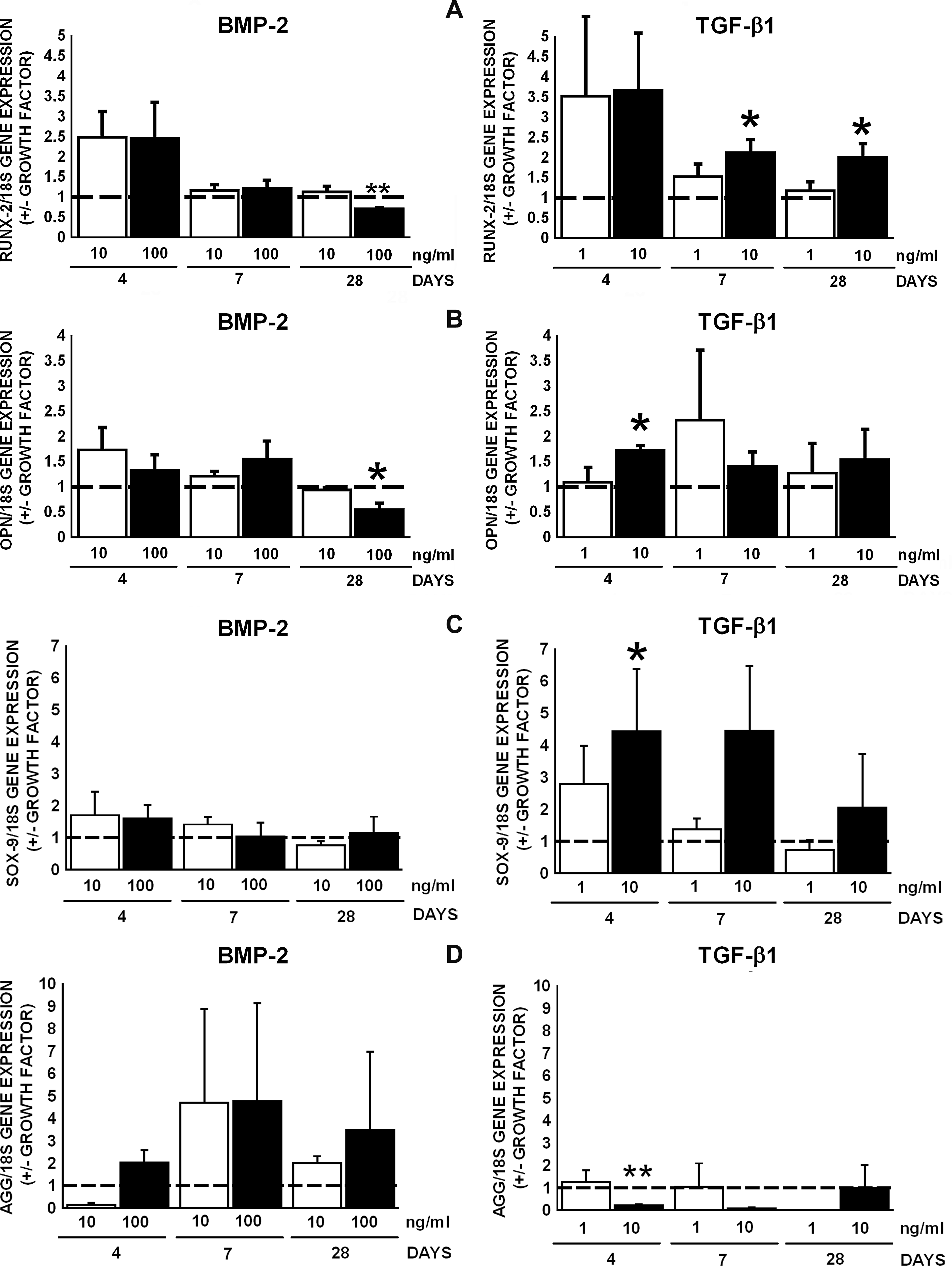
Effect of BMP-2 and TGF-β1 on gene expression of RUNX2 and OPN of AT-MSCs. (
BMP-2 (100 ng/mL) increased ALP activity, which indicates osteogenic differentiation of AT-MSCs, by 1.2-fold at day 28, whereas TGF-β1 had no effect (Fig. 3). BMP-2 and TGF-β1 did not affect ALP activity in AT-MSCs at days 4 and 7 (data not shown).

Effect of BMP-2 (10 and 100 ng/mL) or TGF-β1 (1 and 10 ng/mL) on ALP activity of AT-MSCs. BMP-2 at 100 ng/mL increased ALP activity in AT-MSCs by 1.2-fold at day 28, but TGF-β1 had no effect. Values are mean ± SEM. BMP-2, bone morphogenetic protein-2; TGF-β1, transforming growth factor-β1; AT-MSCs, adipose tissue–derived mesenchymal stem cells; ALP, alkaline phosphatase. Significant effect of growth factor, *p < 0.05.
BMP-2 (10 ng/mL) and TGF-β1 (10 ng/mL) upregulated gene expression of COL1A2, the substrate for collagen-modifying enzymes, by 1.8-fold at day 4 (Fig. 4). Changes in gene expression of COL1A2 in response to BMP-2 or TGF-β1 were not observed at day 7 or 28 (Fig. 4). All AT-MSC cultures contained collagen protein after 28 days of treatment with or without BMP-2 or TGF-β1 (Table 2). The amount of collagen protein in AT-MSC cultures ranged from 119 ± 48 μg to 153 ± 50 μg (Table 2).
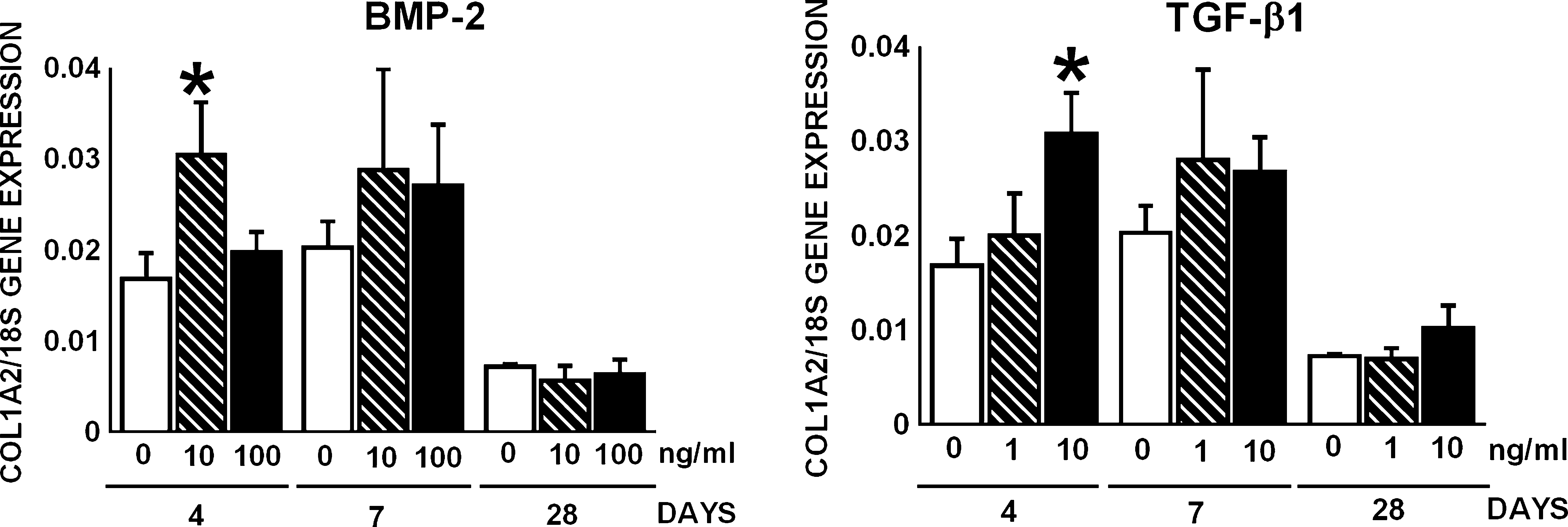
Effect of BMP-2 (10 and 100 ng/mL) or TGF-β1 (1 and 10 ng/mL) on COL1A2 gene expression by AT-MSCs. BMP-2 at 10 ng/mL and TGF-β1 at 10 ng/mL upregulated COL1A2 gene expression by 1.9-fold at day 4. Values are mean ± SEM. BMP-2, bone morphogenetic protein-2; TGF-β1, transforming growth factor-β1; AT-MSCs, adipose tissue–derived mesenchymal stem cells; COL1A2, α2(I) procollagen. Significant effect of growth factor, *p < 0.05.
Data are mean ± SEM per triple helix of collagen, except data on the amount of collagen present per AT-MSC culture in μg (n = 3).
Hyl, hydroxylysine; TH, triple helix; LP, lysyl pyridinoline; HP, hydroxylysyl pyridinoline.
PLOD2 encodes lysyl hydroxylase 2, the lysyl hydroxylase responsible for hydroxylation of the telopeptidal lysine residues. BMP-2 at 10 and 100 ng/mL or TGF-β1 at 1 and 10 ng/mL did not affect PLOD2 gene expression by AT-MSCs at days 4, 7, and 28, when normalized for 18S (Fig. 5A). PLOD2 gene expression relative to COL1A2 was decreased after treatment with BMP-2 (10 ng/mL) by 1.5-fold at day 4 (Fig. 5B). BMP-2 (10 ng/mL) increased PLOD2 gene expression relative to COL1A2 by 1.7 and 1.8-fold at days 7 and 28, and BMP-2 (100 ng/mL) by 1.6-fold at day 28, whereas TGF-β1 had no effect (Fig. 5B).
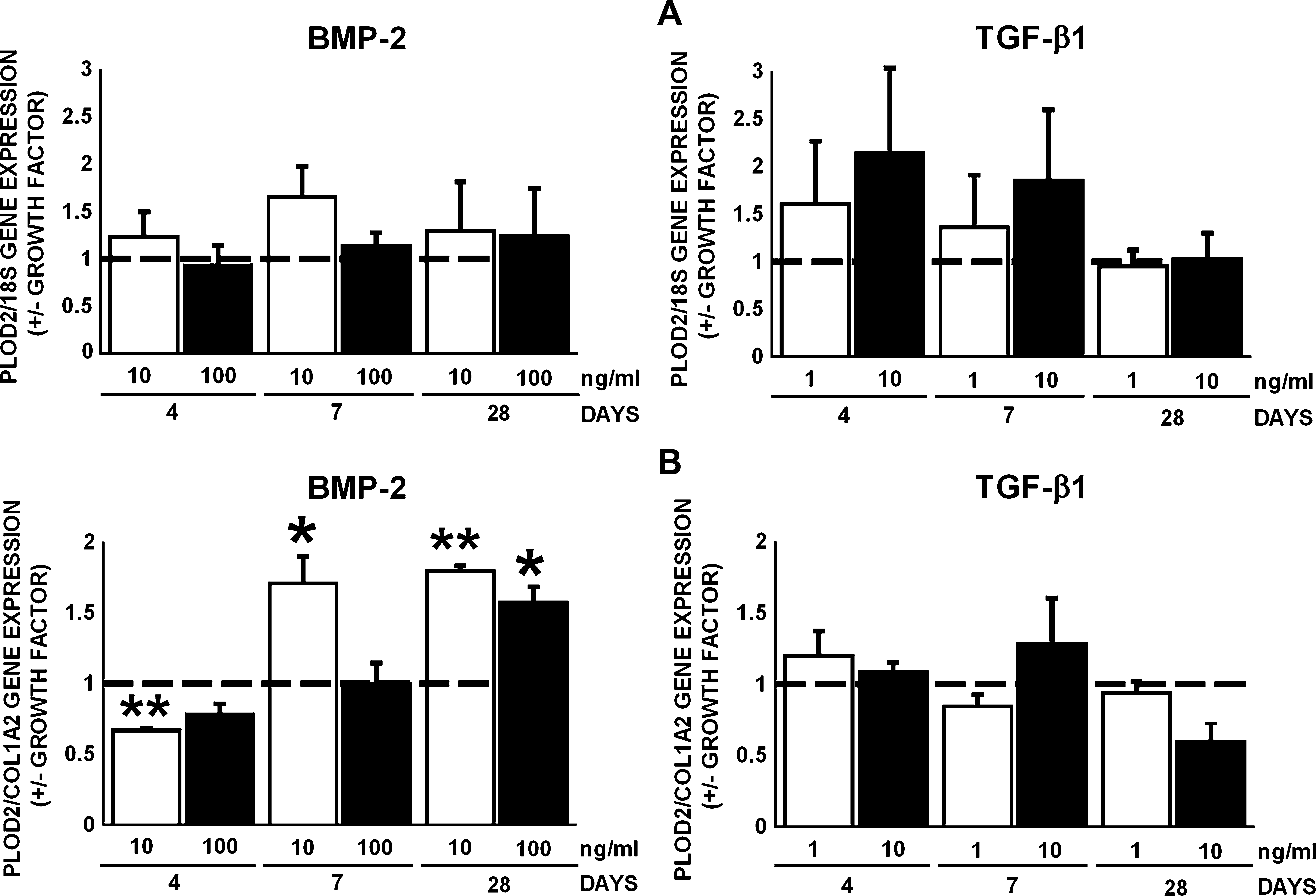
Effect of BMP-2 (10 and 100 ng/mL) or TGF-β1 (1 and 10 ng/mL) on gene expression of PLOD2, encoding lysyl hydroxylase 2, in AT-MSCs. (
PLOD1 and PLOD3 encode the lysyl hydroxylases hydroxylating triple helical lysine residues, lysyl hydroxylase 1 and 3. Both BMP-2 (100 ng/mL) and TGF-β1 (10 ng/mL) upregulated PLOD1 gene expression by 1.5-fold at day 4 (Fig. 6A). However, TGF-β1 (1 ng/mL) downregulated PLOD1 gene expression by 2.1-fold at day 28 (Fig. 6A). BMP-2 (100 ng/mL) increased PLOD1 gene expression relative to COL1A2 by 1.4-fold at day 4. TGF-β1 (1 ng/mL) decreased PLOD1 gene expression relative to COL1A2 by 1.3-fold at day 7, and TGF-β1 (1 and 10 ng/mL) decreased PLOD1 gene expression relative to COL1A2 by 1.7–2-fold at days 4, 7, and 28 (Fig. 6B). TGF-β1 (10 ng/mL) decreased PLOD3 gene expression by 1.9-fold at day 28, but BMP-2 had no effect (Fig. 6C). BMP-2 and TGF-β1 did not affect PLOD3 gene expression relative to COL1A2 (Fig. 6D).
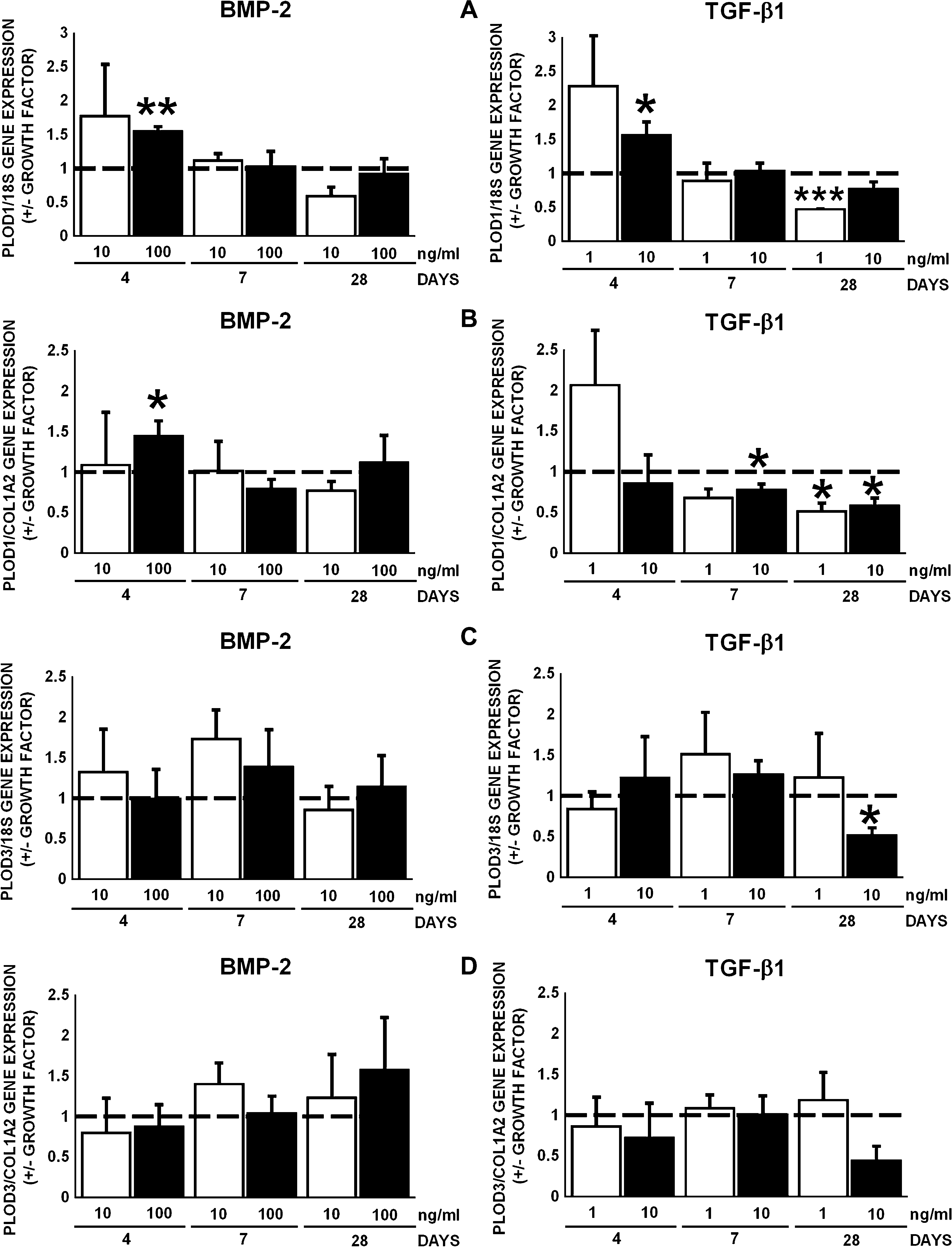
Effect of BMP-2 (10 and 100 ng/mL) or TGF-β1 (1 and 10 ng/mL) on gene expression of PLOD1 and PLOD3, encoding lysyl hydroxylase 1 and 3, in AT-MSCs (
The enzyme LOX catalyzes the oxidative deamination of telopeptidyl lysyl and hydroxylysyl residues of collagen in the extracellular space. BMP-2 (10 and 100 ng/mL) upregulated LOX gene expression normalized for 18S by 1.7–1.8-fold at day 7 (Fig. 7A). TGF-β1 (10 ng/mL) upregulated LOX gene expression normalized for 18S by 1.9-fold at day 4 (Fig. 7A). BMP-2 and TGF-β1 did not affect LOX gene expression relative to COL1A2 (Fig. 7B).
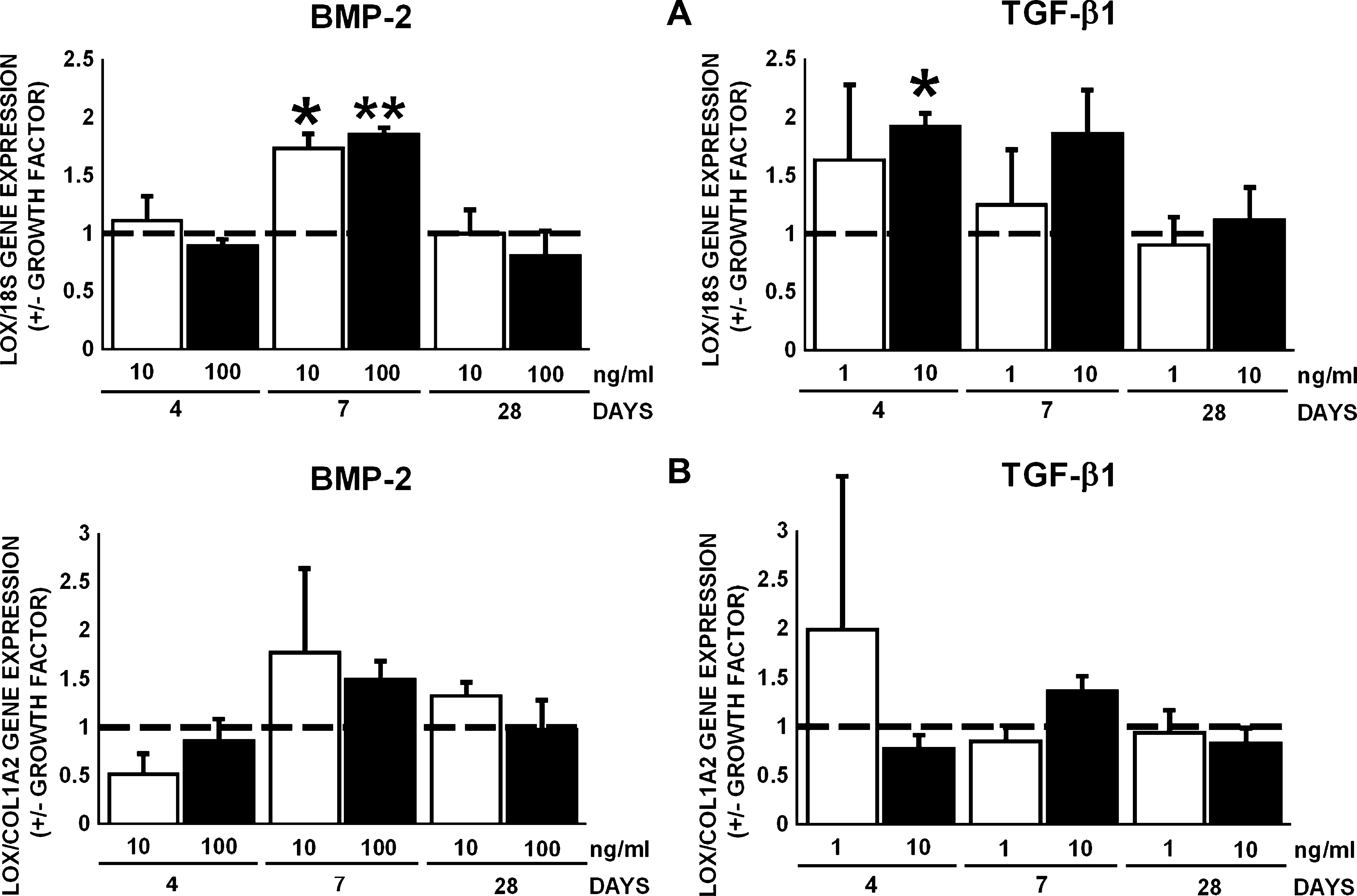
Effect of BMP-2 (10 and 100 ng/mL) or TGF-β1 (1 and 10 ng/mL) on gene expression of LOX in AT-MSCs. (
Lysyl hydroxylases and LOX are needed to posttranslationally modify collagen as preparation for its subsequent cross-linking. We have studied the amount of Hyl and mature trifunctional cross-links HP and LP in the extracellular matrix produced by AT-MSCs in the presence or absence of BMP-2 or TGF-β1 (Table 2). BMP-2 and TGF-β1 did not affect the amount of Hyl, which ranged from 34.6 ± 2.3 to 41.7 ± 4.9 (mean ± SEM) residues per triple helix, the amount of LP, which ranged from 0.02 ± 0.01 to 0.06 ± 0.05 (mean ± SEM) cross-links per triple helix, and the amount of HP, which ranged from 0.32 ± 0.15 to 0.97 ± 0.79 (mean ± SEM) cross-links per triple helix of collagen. The HP/LP ratio ranged from 17.1 ± 2.3 to 20.2 ± 4 (mean ± SEM). The presence of Hyl and of mature trifunctional cross-links HP and LP in AT-MSC cultures suggests the presence of functional lysyl hydroxylases and LOX.
Discussion
For appropriate functioning of engineered tissue, an extracellular matrix that is either provided in form of scaffolds or produced by the cells themselves is of utmost importance. 46 Organic bone extracellular matrix, which is mainly composed of covalently cross-linked type I collagen, is a prerequisite for proper bone function. 47 BMP-2 and TGF-β1 are important growth factors present in bone extracellular matrix, and are involved in bone formation and bone morphogenesis.29,30 Unlike BMP-2, TGF-β1 will not produce bone when injected into ectopic sites.31–33 The present study compares for the first time the effects of BMP-2 and TGF-β1 on gene expression of COL1A2 and collagen-modifying enzymes, lysyl hydroxylase 1, 2, and 3, which are encoded by the genes PLOD1, 2, and 3, and LOX in AT-MSCs. This study provides insight into the effects of these two different growth factors on posttranslational modification of type I collagen before the formation of extracellular matrix by AT-MSCs in vitro. We found that BMP-2, but not TGF-β1, increased ALP activity, indicating that BMP-2 induced osteogenic differentiation of AT-MSCs. Both growth factors differentially affected RUNX2 and OPN gene expression, suggesting that AT-MSCs undergo osteogenic differentiation in response to BMP-2 and TGF-β1. AT-MSCs increase their SOX9 gene expression, which is expressed during chondrogenic differentiation in response to TGF-β1. However, this is not accompanied by an increase in AGG gene expression. AGG gene expression in AT-MSCs decreased in response to TGF-β1. BMP-2 did not affect SOX9 or AGG gene expression, and SOX5 gene expression was detectable, but not quantifiable. These findings suggest that AT-MSCs do not undergo chondrogenic differentiation in response to BMP-2 or TGF-β1. AT-MSCs might need 3-D culture conditions for chondrogenic differentiation. 36 In addition, neither BMP-2 nor TGF-β1 affected pparg gene expression, suggesting that AT-MSCs do not undergo adipogenic differentiation. BMP-2 and TGF-β1 differentially affected COL1A2, PLOD1, and LOX gene expression. PLOD3 was decreased by TGF-β1. When gene expression was normalized for COL1A2 gene expression, we observed that BMP-2 and TGF-β1 differentially affected the expression of PLOD1 and PLOD2. The presence of Hyl and HP and LP cross-links in extracellular matrix produced by human AT-MSCs provides evidence for the presence of functional lysyl hydroxylases and LOX in the AT-MSC cultures. This suggests that the type I collagen matrix that is synthesized by AT-MSCs is posttranslationally modified and subsequently cross-linked, and that expression of genes involved in posttranslational modification is differentially affected dependent on the growth factor added.
PLOD2 gene expression increases in murine bone marrow–derived mesenchymal stem cells at the onset of mineralization, and lysyl hydroxylase 2 has been suggested to be responsible for telopeptidal lysyl hydroxylation in bone. 48 However, substrate specificity of any of the known lysine hydroxylases, for example, for type I collagen, cannot be assigned. 49 Patients suffering from Bruck syndrome show skeletal defects such as short stature or osteoporosis, and underhydroxylation of type I collagen and aberrant cross-linking, which leads to the observed phenotype.21,23–25 A mutation in PLOD2 is responsible for this Bruck syndrome in a subset of patients, and lysyl hydroxylase 2 has been identified as telopeptidal lysyl hydroxylase.21,23,24 Lysyl hydroxylase 2b, a splice variant of lysyl hydroxylase 2, has been associated with collagen cross-linking in MC3T3-E1 osteoblasts, and lysyl hydroxylase 2b overexpression results in defective matrix mineralization by MC3T3-E1 cells.50,51 Therefore, the increase in PLOD2 relative to COL1A2 gene expression caused by BMP-2 treatment of AT-MSCs suggests that telopeptidal Hyl may be present. Thus, trivalent mature cross-links, HP and LP, may be formed in between type I collagen triple helices, which may result in a properly cross-linked collagen extracellular matrix. Whether lysyl hydroxylase 2b is expressed in differentiating AT-MSCs and whether lysyl hydroxylase 2b expression levels change during the course of osteogenic differentiation are subject of further study.
Lysyl hydroxylase 1 and lysyl hydroxylase 2 have been proposed to have telopeptidal lysine hydroxylation activity.52,53 BMP-2 and TGF-β1 induced PLOD1 gene expression, which is encoding lysyl hydroxylase 1. Lysyl hydroxylase 1 might be responsible for hydroxylation of triplehelical lysine residues in type I collagen or other helical collagens, produced by differentiating AT-MSCs. This might compensate for a lack of or for low telopeptidal hydroxylation activity of lysyl hydroxylase 2, because PLOD2 gene expression was not directly affected by BMP-2, but was increased relative to its substrate COL1A2. It is thus likely that telopeptidal hydroxylysyl residues are present and that AT-MSCs are able to form an appropriate extracellular matrix. This, however, does not explain the downregulation of PLOD2 by BMP-2 and TGF-β1 at a late time point, which needs further investigation.
Lysyl hydroxylase 3, the product of the PLOD3 gene, is a multifunctional enzyme that not only hydroxylates triple helical lysine residues, but also possesses collagen glucosyltransferase and galactosyltransferase activity.16,19,20 Lysyl hydroxylase 3 has been proposed to affect matrix remodeling resulting from modification of proteins in the extracellular matrix, due to its localization in the extracellular space. 54 PLOD3 expression levels are not changed during MC3T3-E1 osteoblast differentiation,54,55 which might explain our finding that BMP-2 did not affect PLOD3 gene expression in AT-MSCs. Thus, PLOD3 might be constitutively expressed due to its extracellular matrix housekeeping function. Glucosyltransferase and galactosyltransferase activity of lysyl hydroxylase 3 has been shown in, for example, spleen, brain, and lung. 54 However, this has been studied neither in bone cells or bone tissue nor in stem cells. Further investigation could provide insight into a possible extracellular matrix housekeeping function of PLOD3 (lysyl hydroxylase 3) in bone or during osteogenic differentiation of stem cells and thus the formation of a functional collagen matrix.
LOX catalyses the oxidative deamination of telopeptidal lysyl or hydroxylysyl residue, forming aldehyde moieties, which is the final posttranslational change of collagen before the formation of covalent cross-links.47,56 The enzyme is located in the extracellular space and interacts with the secreted collagen triple helices.57,58 LOX plays a key role in collagen deposition during the course of MC3T3-E1 osteoblast differentiation and chicken calvarial osteoblasts.56,59–61 In the current study, upregulation of LOX gene expression in AT-MSCs by BMP-2 occurs after the upregulation of COL1A2 and PLOD1 (lysyl hydroxylase 1) gene expression. It has been shown in MC3T3-E1 osteoblasts that LOX gene expression follows collagen gene expression, and that LOX activity precedes accumulation of insoluble, cross-linked collagen. 56 TGF-β1 has been shown to directly affect LOX gene expression and LOX activity in MC3T3-E1 osteoblasts, and LOX upregulation in MC3T3-E1 cells coincides with type I collagen gene expression. 60 In addition, processing of the LOX proenzyme appears to occur at all stages of osteoblast differentiation in MC3T3-E1. 56 This suggests that, in our study, the osteogenically differentiating AT-MSCs might posttranslationally modify LOX. Thus, active LOX might be present, which in turn modifies type I collagen synthesized by AT-MSCs. This might lead to appropriately cross-linked collagen in the extracellular matrix produced by AT-MSCs after treatment with BMP-2 or TGF-β1.
Biochemical analysis of the extracellular matrix produced by AT-MSCs was not affected by BMP-2 or TGF-β1. Interestingly, the HP/LP ratios were comparable with HP/LP ratios found during fracture repair 3–5 weeks after fracture in a canine model. 62 At later stages during fracture repair, the HP/LP ratio normalized, resembling a bone-like matrix. 62 The amount of lysyl hydroxylation of collagen produced by AT-MSCs resembled lysyl hydroxylation in canine callus 7 weeks after fracture. Thus, the matrix produced by AT-MSCs might resemble an immature collagen matrix that needs to be remodeled to become a bone-like matrix. However, the changes in gene expression of collagen-modifying enzymes in response to BMP-2 and TGF-β1 did not affect the collagen matrix produced by AT-MSCs. Because BMP-2 affects gene expression of collagen-modifying enzymes and the Hyl content of the matrix produced by C2C12 cells early during differentiation, further experimentation is needed to assess the effect of BMP-2 and TGF-β1 on younger extracellular matrix produced by AT-MSCs, and compare this to a more mature matrix with trifunctional cross-links. 35
In conclusion, our findings indicate that treatment with BMP-2 or TGF-β1 changes the gene expression levels of collagen-modifying enzymes PLOD1, 2, and 3 and LOX in AT-MSCs, and that BMP-2 and TGF-β1 have differential effects on the expression of collagen-modifying enzymes. Therefore, AT-MSCs may be able to appropriately modify type I collagen to form an organic bone extracellular matrix for tissue engineering purposes depending on the growth factor added.
Footnotes
Acknowledgments
The authors thank F.J. van Milligen for providing human AT-MSCs, J. Snabel and J.M.A. de Blieck-Hogervorst for excellent technical support, and The Technology Foundation STW for supporting the work of M. Knippenberg (STW Grant # VPG. 5935). The Dutch Program for Tissue Engineering (DPTE) supported the work of J. Klein-Nulend (DPTE Grant # BGT. 6744). These studies were performed under the auspices of the Skeletal Tissue Engineering Group Amsterdam (STEGA).
Disclosure Statement
No competing financial interests exist.