
Abstract
To elucidate the characterizations of chondrocytes originating from auricular cartilage (donors: 10–15 years) and nasoseptal one (20–23 years), we evaluated proliferation or matrix synthesis of both cells cultured under monolayer and collagen type I (COL1) three-dimensional (3D) conditions. Three passages were needed until cell numbers of auricular chondrocytes in the 3D culture increased 1000-fold, although those in monolayer culture or nasoseptal monolayer and 3D cells reached a 1000-fold increase at four passages. When we cultured the tissue-engineered cartilage pellets made of the chondrocytes proliferated at 1000-fold increase, the pellets of monolayer cells maintained their sizes during the culture period. However, those of nasoseptal 3D cells began to shrink at day 1 and became approximately one-tenth in size at day 21. The downsizing of pellets may result from the upregulation of tumor necrosis factor (TNF)-α or the related proteinases, including matrix metalloproteinases (MMPs)-1, -2, and -3, and cathepsin B, suggesting that the nasoseptal chondrocytes, which are physiologically separated from COL1, may be hardly adapted for the COL1 3D proliferation condition. Ideally, these characteristics would have been compared between the chondrocytes from donors that are completely matched in ages. However, according to our data using closely matched ones, the auricular chondrocytes seemed to more rapidly proliferate and produce less proteinases during this 3D culture than the nasoseptal ones.
Introduction
Based on the matrices contained, cartilage is classified into three categories: hyaline cartilage, elastic cartilage, and fibrocartilage. Articular and nasoseptal cartilages belong to hyaline cartilage that exclusively consists of collagen type II (COL2) and proteoglycan,5–7 while typical elastic cartilage that is rich in flexibility and contains many elastic fibers exists in the auricles. 8 Some parts of costal cartilage and annulus fibrosus in vertebral discs include collagen type I (COL1), termed fibrocartilage. All these kinds of cartilaginous tissues can be candidates for cell sources. For the selection of chondrocyte sources suitable for certain therapies, we should cautiously consider the volume limit of donor tissues, the invasiveness of biopsy, or the differences in cytological properties of chondrocytes from various donor sites.
Some previous papers compared the cytological properties among human chondrocytes originating from articular cartilage, auricular cartilage, costal cone, and nasal septum.9–11 Auricular and nasoseptal chondrocytes showed better proliferation rates than those from costal one, while the tissue-engineered cartilage pellets made of auricular and nasoseptal chondrocytes accumulated cartilaginous matrices more abundantly than those of costal ones. 9 It was also reported that the adult nasoseptal chondrocytes proliferated approximately four times faster than the articular type, while the nasoseptal chondrocytes generated a matrix with significantly higher fractions of COL2 and glycosaminoglycans (GAGs) than the articular chondrocytes. 10 Between auricular chondrocytes and nasoseptal ones, engineered cartilage made of both cells had been compared, showing that the auricular chondrocytes generated significantly less GAG in engineered cartilage than the nasoseptal ones, although COL2 was found accumulated in both cartilages by immunohistological examinations. 11
All of those experiments had applied monolayer culture for chondrocyte preparation with sufficient cell numbers. However, when the chondrocytes are expanded in the monolayer culture, they inevitably lose chondrocyte phenotypes and stop producing the characteristic molecules including COL2 and proteoglycans, termed “dedifferentiation.” 12 To prevent the dedifferentiation of chondrocytes, several researchers had attempted to apply the collagen gel embedding for expansion culture in human chondrocytes.13–16 We also succeeded in the collagen gel three-dimensional (3D) culture of human auricular chondrocytes providing a 1000-fold increase in cell numbers at passage 3 and reducing chondrocyte dedifferentiation more effectively than conventional monolayer culture. 17 Therefore, when we discuss the cytological characterizations of cultured chondrocytes, we should culture the cells under not only a monolayer condition but also 3D condition.
From the standpoints of invasiveness during biopsy or tissue volume obtained, auricular cartilage or nasal septum may be feasible, because both of them are located just under the skin tissue and can be sufficiently obtained from subcutaneous biopsy. Considering the further widening of the indication range for cartilage tissue engineering to maxillofacial areas, these two tissues should be focused on as favorable chondrocyte sources. In the present study, we attempted monolayer culture as well as collagen gel 3D culture for chondrocyte expansion derived from human auricular and nasoseptal cartilages, and examined their cytological characterizations.
Materials and Methods
Chondrocyte isolation and proliferation
All procedures of the present experiments were approved by the ethics committee of the University of Tokyo Hospital (ethics permission #622). Auricular cartilage materials were obtained from five children (age range, 10–15 years), while nasoseptal ones from three adults (age range, 20–23 years). They underwent surgery at the University of Tokyo Hospital, with informed consent. Both cartilage materials were minced into 1-mm3 pieces and digested with 0.15% collagenase in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO) containing penicillin and streptomycin at 37°C for 24 h. The digested suspension was filtered using a sterile 100-μm nylon cell strainer (BD Falcon, Bedford, MA), centrifuged at 430 g for 5 min, and washed twice with DMEM containing the antibiotics. The number of cells was calculated using a hemocytometer, and the viability of the cells was determined using trypan blue vital dye.
In monolayer culture, the isolated chondrocytes were seeded in a 100-mm plastic tissue culture dish at a density of 2500 cells/cm2 and were cultured in a 37°C/5% CO2 incubator with the bullet kit chondrocyte growth medium (CGM; Cambrex Bioscience Walkersville, Walkersville, MD). The CGM contained 5% fetal bovine serum with an undisclosed concentration of fibroblast growth factor-2, insulin-like growth factor-I, and insulin. Passages were performed using a treatment with trypsin–EDTA solution (Sigma-Aldrich) when the cells were approaching confluence at 7 days. For the 3D culture, the chondrocytes were suspended in 0.24% atelocollagen solution (pH 7). The mixture was placed in each well of a 100-mm plastic culture dish at 10 mL at a density of 2 × 104 cells/mL. The atelocollagen formed a gel during 1-h incubation at 37°C, embedding the cells under 3D condition. The CGM was gently poured on the gel at a volume of 10 mL in a 37°C/5% CO2 incubator. To release the cells, the gel was incubated in 0.3% collagenase at 37°C for 30 min, and passages were performed at intervals of 7–10 days. In both cultures, the medium was changed three times/week. The chondrocytes from eight different patients (five for auricular chondrocytes and three for nasoseptal ones) were individually used for cell proliferation experiments. The cell proliferation data were obtained from separate experiments using chondrocytes from each patient.
Generation of tissue-engineered cartilage
The chondrocytes proliferated at a 1000-fold increase in numbers were suspended in 0.8% of atelocollagen solution at a density of 107 cells/mL. In the atelocollagen, 20 μL of the cell/material suspension (total 2 × 105 cells) was placed into the bottom of a 15-mL polypropylene conical tube (BD Falcon) to form a gel in 1-h incubation at 37°C. The DMEM/F-12 medium (Sigma-Aldrich) was used at a volume of 2 mL for each gel and cultured in a 37°C/5% CO2 incubator. To induce redifferentiation of the chondrocytes, 5 μg/mL insulin (MP Biomedicals, Irvine, CA), 200 ng/mL bone morphogenetic protein-2 (BMP-2; kindly provided by Astellas Pharma, Tokyo, Japan), and 100 nM L-3,3′,5′-triiodothyronine (rhT3; EMD Bioscience, San Diego, CA) were added to the medium, according to the previous report. 18 Throughout the experiment, the medium was changed three times/week. The chondrocytes from eight different patients (five for auricular chondrocytes and three for nasoseptal ones) were individually used for the experiments of tissue-engineered cartilage formation.
Total RNA extraction and real-time RT-PCR analysis
Total RNA was isolated from chondrocytes with ISOGEN (Wako Pure Chemical Industries, Osaka, Japan) following the supplier's protocol. Complementary DNA (cDNA) was synthesized from 1 μg of total RNA with the Superscript II reverse transcriptase kit (Invitrogen, Carlsbad, CA). The full-length or partial-length cDNA of the target genes, including PCR amplicon sequences, was amplified by PCR, cloned into pCR-TOPO Zero II or pCR-TOPO II vectors (Invitrogen), and used as standard templates after linearization. With QuantiTect SYBR Green PCR Master Mix (Invitrogen), real-time fluorescence detection was performed with an ABI 7700 Sequence Detection system with the following protocol: initial denaturation for 10 min at 94°C followed by 40 cycles consisting of 15 s at 94°C and 1 min at 60°C. All reactions were run in quadruplicate. Sequences of the primers were 5′-CTCCTCGCTTTCCTTCCTCT-3′ and 5′-GTGCTAAAGGTGC CAATGGT-3′ for type I collagen αI chain (COL1A1); 5′-GAGTCAAGGGTGATCGTGGT-3′ and 5′-CACCTTGGTCT CCAGAAGGA-3′ for type II collagen αI chain (COL2A1); 5′-GGGAGATCATGGGGACAACTC-3′ and 5′-GGGCCTGGTTGAAAAGCAT-3′ for MMP-1; 5′-TCAAGTTCCCCGGCG-3′ and 5′- TGTTCAGGTATTGCA-3′ for MMP-2; 5′-TTTTGGCCATCTCTT-3′ and 5′-TGTGGATGCCTCTTG-3′ for MMP-3; 5′- TCCTACAAATCTCGCGGGAAT-3′ and 5′-GCATTTCTCGGAGCCTCTCA-3′ for MMP-13; 5′-TGTGTATTCGGACTTCCTGCT-3′ and 5′-GTGTGCCATTCTCCACTCC-3′ for cathepsin B; 5′-GAAGGTGAAGGTCGGAGTCA-3′ and 5′- GAAGATGGTGATGGGATTTC-3′ for GAPDH.
GAG measurement, ELISA for COL1 and COL2, and histology
The sulfated GAG content was measured using Alcian blue binding assay (Wieslab AB, Lund, Sweden). After digestion of the regenerated cartilage in 0.3% collagenase for 1 h at 37°C, the cell debris and insoluble material were removed by centrifugation at 6000 g for 30 min. GAG in the supernatant was precipitated with Alcian blue solution, and the sediments obtained by centrifugation at 6000 g for 15 min were dissolved again in 4 M GuHCl–33% propanol solution. The spectrophotometrical absorbance of the mixture was measured at a wavelength of 600 nm.
The collagen proteins of the regenerated cartilage were quantified by ELISA using a human Type 1, 2 Collagen Detection Kit (Chondrex, Redmond, WA). The regenerated cartilage was dissolved in 10 mg/mL pepsin/0.05 M acetic acid at 4°C for 48 h and then in 1 mg/mL pancreatic elastase/0.1 M Tris–0.2 M NaCl–5 mM CaCl2 at 4°C overnight. The samples were centrifuged at 9100 g for 5 min to remove the residue. The collagen proteins were captured by polyclonal anti-human COL1 or COL2 antibodies and detected by biotinylated counterparts and Streptavidin Peroxidase. Both of o-phenylenediamine and H2O2 were added to the mixture, and the spectrophotometrical absorbance of the mixture was measured at a wavelength of 490 nm.
The regenerated cartilage was fixed with 4% paraformaldehyde, embedded in Tissue-Tek® optimal cutting temperature compound (Sakura Seiki, Tokyo, Japan), and cryosectioned into 10-μm slices. The sections were stained with toluidine blue-O.
Measurement of inflammatory cytokines
The chondrocytes, which had increased 1000-fold in cell numbers (passages 4 and 3 under a monolayer or 3D culture, respectively), were cultured without any medium changes. The culture media were harvested for 7 days and were stored at −80°C until further use. The levels of IL-1β and tumor necrosis factor (TNF)-α were determined in the culture medium using a QuantiGlo chemiluminescent ELISA kit (R&D systems, Minneapolis, MN) following the manufacturer's instruction.
Results
Chondrocyte culture and growth
Human auricular and nasoseptal chondrocytes were isolated from surgical waste or debris. The mean weight was approximately 0.4 g (n = 5) and 0.6 g (n = 3), while the cell numbers of isolated chondrocytes were 2.6 × 106 (1.0–6.5 × 106, n = 5) and 2.3 × 106 (1.0–4.0 × 106, n = 3), respectively. The chondrocytes of both origins similarly became spindles during monolayer culture or 3D embedding culture (Fig. 1A). We examined the cell growth of chondrocytes from the different origins under both monolayer and 3D embedding culture. It took three passages until the cell numbers of auricular chondrocytes in the 3D culture (auricular 3D cells) increased 1000-fold, although those in monolayer culture (auricular monolayer cells) reached a 1000-fold increase at four passages. The growth rate of nasal septum chondrocytes in both monolayer and 3D cultures (nasoseptal monolayer and 3D cells) was almost similar to that of auricular monolayer cells (Fig. 1B).
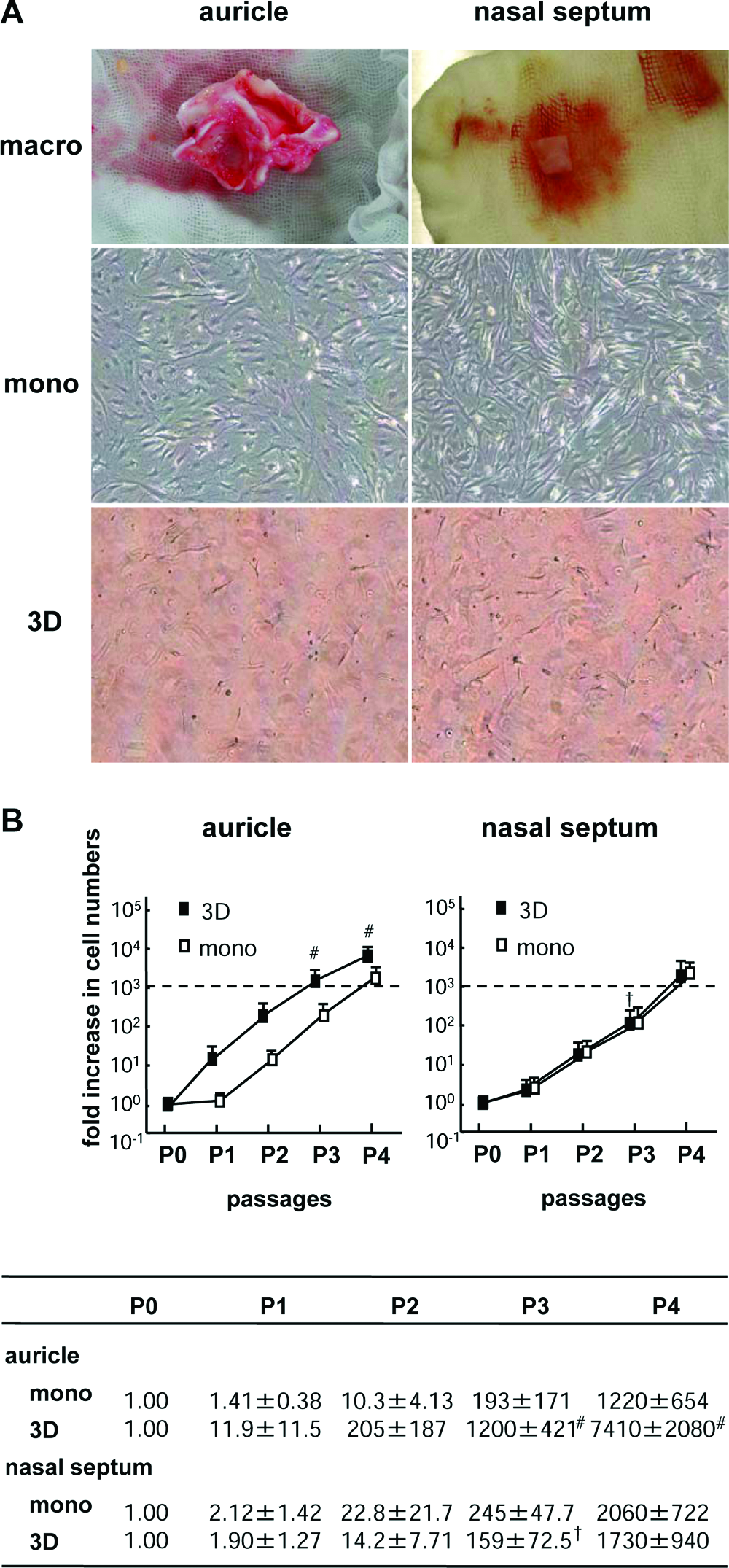
Morphological findings and growth curves of expanded human auricular and nasoseptal chondrocytes. (
Morphological and biochemical changes in tissue-engineered cartilage
Next, we compared the sizes of tissue-engineered cartilage derived from the different origins cultured under a monolayer or 3D culture. When the monolayer or 3D cells proliferated at 1000-fold, the cells were harvested and embedded in atelocollagen gel at a density of 107 cells/mL to make tissue-engineered cartilage pellets. To enhance the maturation of tissue-engineered cartilage, we added BMP-2, insulin, and T3. 18 During the culture, the pellets gradually became round on the bottom of the tubes. The sizes of the pellets made of both auricular monolayer and 3D cells were almost the same over 3 weeks. Although the pellets of nasoseptal monolayer cells tended to be of the same size as those of auricular cells, those of nasoseptal 3D cells began to shrink at day 1 with or without stimulation of growth factors and became approximately one-tenth in size at day 21 (Fig. 2).
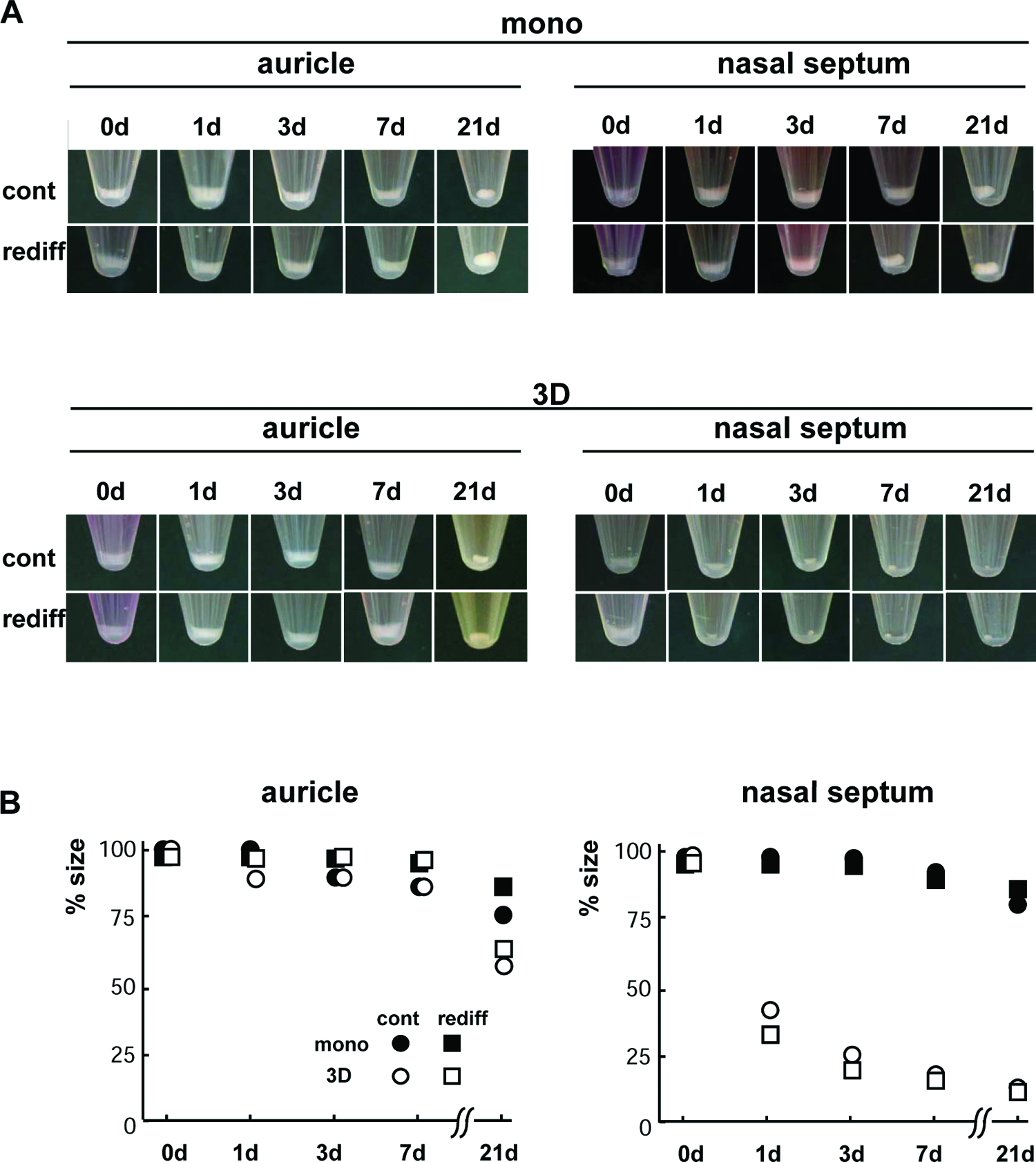
Macroscopic findings of tissue-engineered cartilage. (
The COL1A1 expression in the pellets of either monolayer or 3D cells from both auricular and nasoseptal cartilages was downregulated, according to the addition of BMP-2 and insulin (Fig. 3). Contrarily, the expression of COL2A1 in the auricular cells was prominently enhanced due to the stimulation of growth factors (Fig. 3). The COL2 upregulation with the response to growth factors was more pronounced in the pellets of auricular 3D cells, compared with those of monolayer counterparts. However, in nasoseptal cells, a similar tendency was observed as in the auricular cells, although a statistical significance could hardly be seen (Fig. 3).
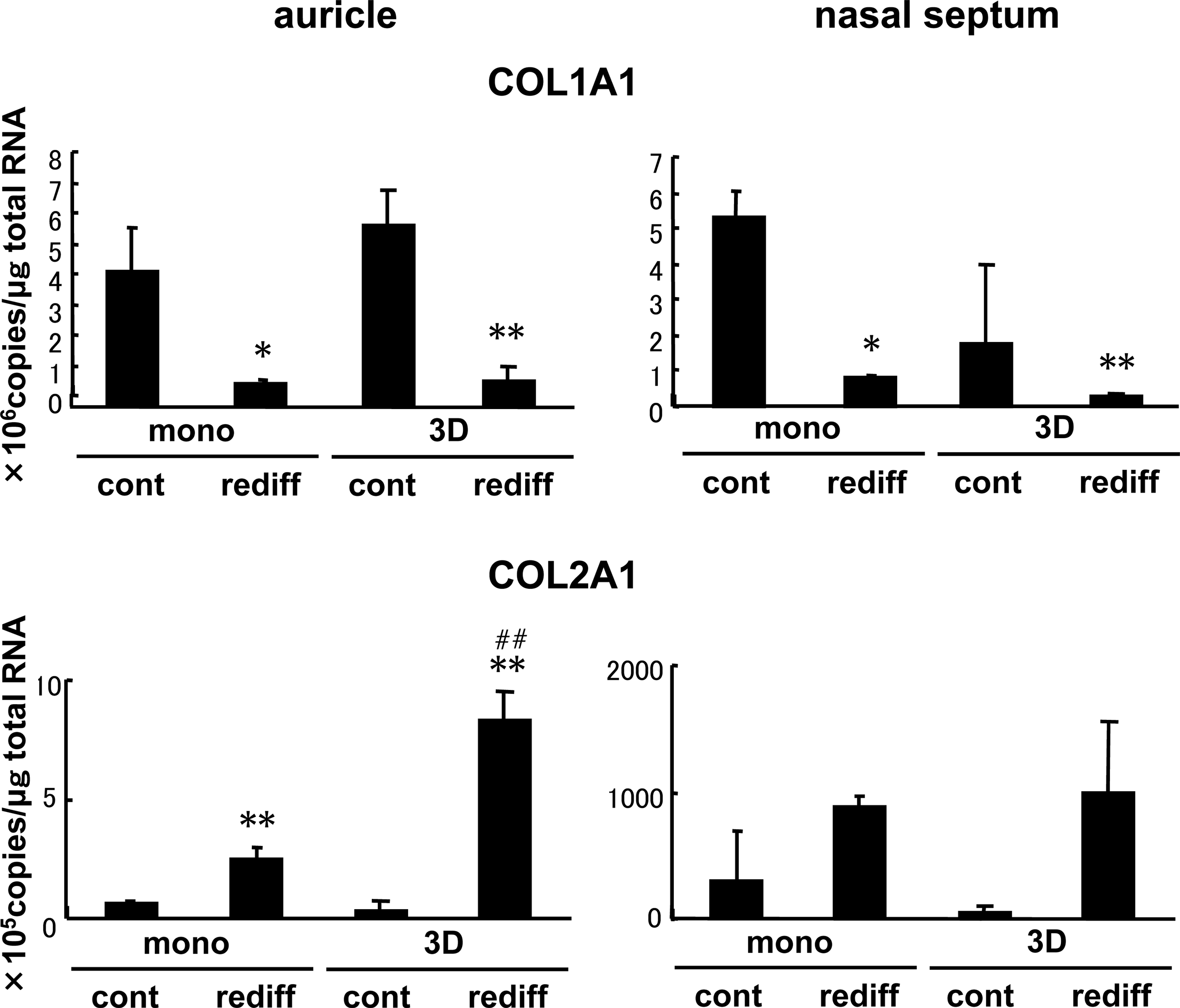
Gene expression of tissue-engineered cartilage pellets made of auricular and nasoseptal chondrocytes. The expressions of COL1A1 and COL2A1 were examined by real-time RT-PCR in the pellets of both monolayer and 3D cells, which were cultured without (cont) or with (rediff ) the stimulation of BMP-2 (200 ng/mL), insulin (5 μg/mL), and T3 (100 nM) for 1 week. All values are presented as mean + standard deviation of three samples from different patients per group. The age ranges of the patients who provided the auricular chondrocytes and the nasoseptal ones were 10–15 years and 20–23 years, respectively. Statistics were assessed using the Student's t-test (*p < 0.05 and **p < 0.005 vs. cont; ##p < 0.005 vs. mono).
Regarding the protein level of tissue-engineered cartilage, COL1 was almost diminished in the pellets of both auricular and nasoseptal 3D cells with or without the simulation of growth factors, while some accumulation was found in those of monolayer cells of both origins (Fig. 4). The pellets of auricular 3D cells more abundantly contained COL2 protein than those of monolayer counterparts when they were stimulated by growth factors (Fig. 4). However, in the nasoseptal chondrocytes, although some response of the COL2 level to the stimulation of growth factors was noted in the pellets of monolayer cells, the COL2 accumulation was hardly observed in 3D cells even with growth factors (Fig. 4). The GAG amount also rose with the stimulation of growth factors compared with that in the control, but it did not respond to the stimulation only in the pellets of nasoseptal 3D cells (Fig. 4).
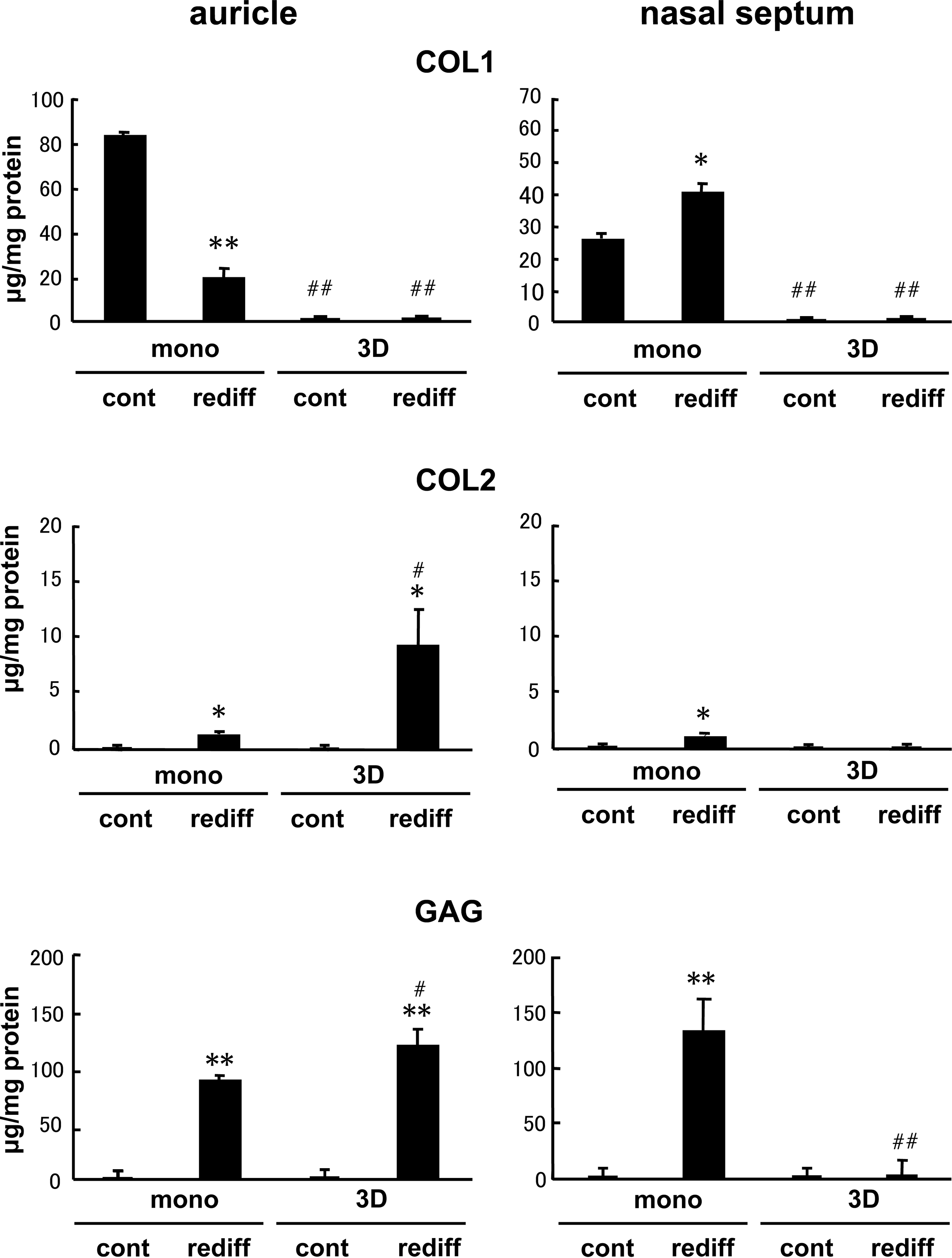
Matrix accumulation of tissue-engineered cartilage pellets made of auricular and nasoseptal chondrocytes. The protein contents of COL1 and COL2 as well as GAG accumulation were examined by ELISA or Alcian blue binding assay in the pellets of both monolayer and 3D cells, which were cultured without (cont) or with (rediff ) the stimulation of BMP-2 (200 ng/mL), insulin (5 μg/mL), and T3 (100 nM) for 3 weeks. All values are presented as mean + standard deviation of three samples from different patients per group. The age ranges of the patients who provided the auricular chondrocytes and the nasoseptal ones were 10–15 years and 20–23 years, respectively. Statistics were assessed using the Student's t-test (*p < 0.05 and **p < 0.005 vs. cont; #p < 0.05 and ##p < 0.005 vs. mono).
The histological findings indicated that the stimulation of growth factors enhanced the metachromagia, implying the accumulation of proteoglycan in tissue-engineered cartilage. The sizes of the pellets of 3D cells tended to be reduced, compared with those of monolayer cells. Particularly, the size reduction was more evident in the pellets of nasoseptal 3D cells, even with the stimulation of growth factors (Fig. 5).

Histological findings of tissue-engineered cartilage. The tissue-engineered cartilage pellets were cultured without (cont) or with (rediff ) the stimulation of BMP-2 (200 ng/mL), insulin (5 μg/mL), and T3 (100 nM) for 3 weeks. Although all sections of the pellets with the stimulation of redifferentiation exhibited metachromagia in toluidine blue suggesting the accumulation of proteoglycan, the pellets of nasoseptal 3D cells were shrunk remarkably. Bar = 1 mm.
Mechanisms of changes in sizes of tissue-engineered cartilage
Considering that the tissue-engineered cartilage made from the nasoseptal 3D cells decreased in sizes and the contents of COL2 or GAG in spite of detectable expression of COL2A1 mRNA, those matrix proteins may be degraded immediately by the secretion of proteinases from the chondrocytes. To confirm this, the expressions of major proteinases were examined by real-time RT-PCR. MMP-1, -2, -3, and -13, and cathepsin B were more highly expressed in the nasoseptal chondrocytes than in the auricular ones. Especially, MMP-1 and -2 were exaggeratedly detected in nasoseptal 3D cells and were upregulated at approximately 1000-fold, compared with auricular cells or nasoseptal monolayer cells (Fig. 6A).
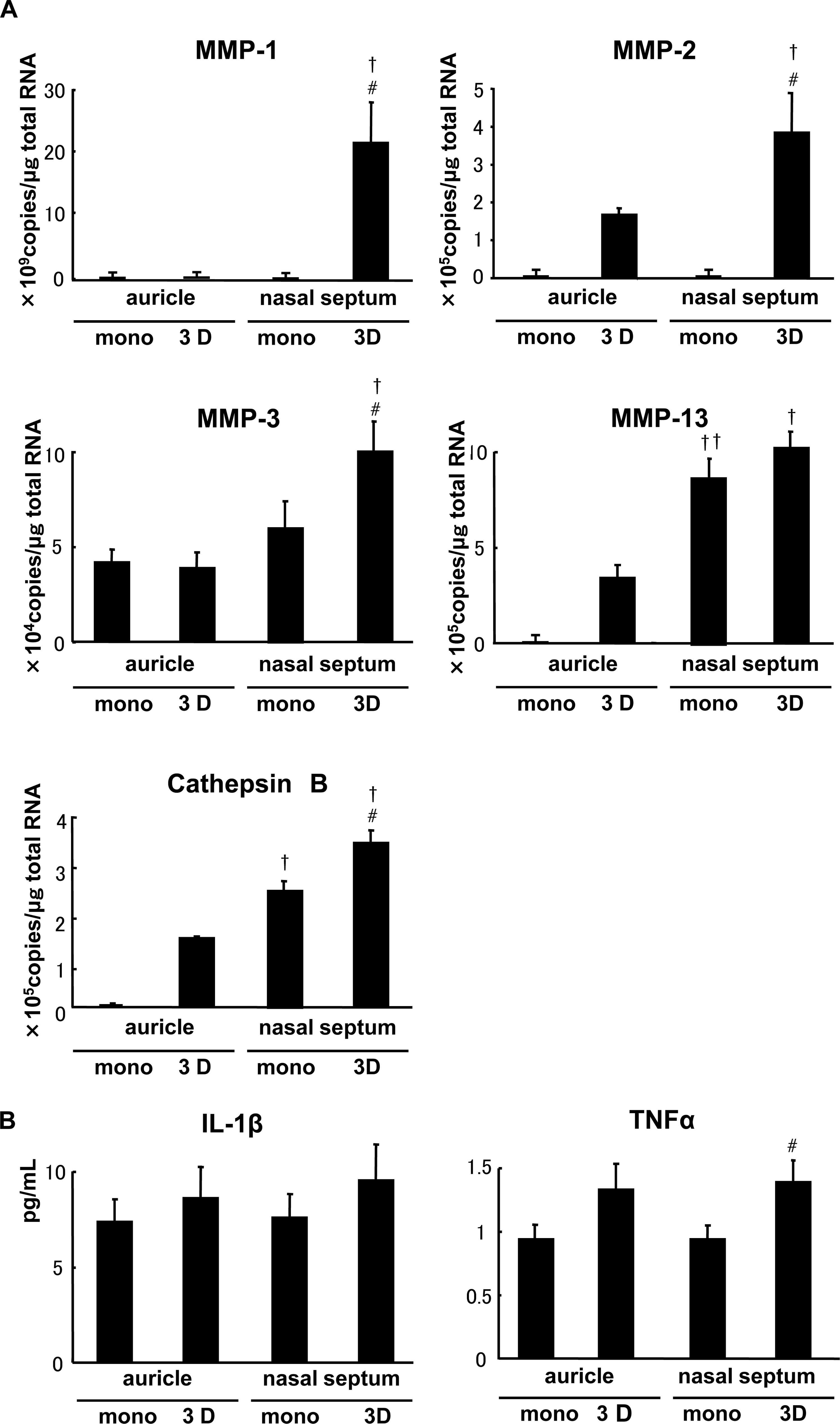
The mechanisms of downsizing of tissue-engineered cartilage pellets in nasoseptal 3D cells. (
In addition, we evaluated the protein contents of inflammatory mediators, TNF-α and IL-1β, which may induce various proteinases in chondrocytes, 19 in the medium during proliferation culture by ELISA. Both cytokine levels in 3D cells tended to increase compared to monolayer cells, while TNF-α in nasoseptal 3D cells was more significantly abundant than in the monolayer counterpart (Fig. 6B).
Discussion
The chondrocytes originating from both nasal septum and auricles showed potent proliferation in either monolayer or 3D condition. Although proliferation of auricular ones under a 3D condition seemed more preferable than the others, both cells under each condition showed no less than a 1000-fold increase after three to four passages. Because the age range showed some differences between the patients providing the auricular chondrocytes and the nasoseptal ones (10–15 years vs. 20–23 years, respectively), the possibility might not be neglected that the difference in proliferation ability under the 3D condition was not due to the chondrocyte origins, but the age of the cartilage donors. However, the complete matching of the donor ages of the auricular cartilage and nasoseptal ones were actually a difficult task due to ethical reasons. When we compared the chondrocytes from the donors, which were matched in ages as closely as possible (10–15 years vs. 20–23 years), interestingly, unlike the 3D condition, the proliferation ability of the auricular monolayer cells showed no significant difference from that of the nasoseptal counterparts in the present experiments (Fig. 1B). If the difference in the proliferation should be related to the ages of the donors and the biological activities were generally diminished in the nasoseptal chondrocytes compared to the auricular ones, some decrease in proliferation ability would be observed not only under the 3D condition, but also under the monolayer condition. Therefore, we speculated that the difference in the proliferation ability between the auricular 3D cells and nasoseptal counterparts were based not on the ages of the donors but on the chondrocyte origins.
Regarding the matrix synthesis of the auricular and nasoseptal chondrocytes cultured under the monolayer condition, cartilaginous matrices including COL2 and GAG were accumulated in the tissue-engineered cartilage pellets. Compared with the pellets of auricular chondrocytes, those of nasoseptal ones had less COL2 but were more enhanced in GAG production. The chondrocytes obtained from different cartilaginous origins appeared to show distinct responses during maturation of tissue-engineered cartilage. The previous study also revealed significant differences in total GAG content between macroaggregates derived from human auricular and nasoseptal chondrocytes, in which the former ones contained significantly less GAG than the latter. 11 As well as in bovine cartilage, the expression of aggrecan in copolymer discs seeded with nasoseptal chondrocytes was higher than that with auricular chondrocytes, while the results of the COL2A1 was opposite. 20 Such a tendency may reflect the original composition of those cartilage subtypes. In bovine cartilage, the GAG content of auricular cartilage showed a lower level compared with that of native hyaline cartilage, but the COL2 level in the former was higher than that in the latter. 21
In 3D cells, the COL2 and GAG were accumulated in the pellets of auricular chondrocytes more abundantly or at least equivalently compared with those of the monolayer counterparts. However, nasoseptal 3D cells minimally contained the cartilaginous matrices in comparison with monolayer counterparts. Because the gene expression of COL2A1 in nasoseptal 3D cells had been confirmed similarly to that in the monolayer counterparts, the drastic decrease in cartilaginous matrices as well as pellet sizes suggested that the materials produced by cells could not be accumulated around the cells and hardly supported the construction of matrix in the engineered tissues. Gel contraction was also observed when bovine articular chondrocytes were seeded in COL1 gel, while MMP-13 was strongly upregulated during the culture. 22 The exact reason for downsizing in tissue-engineered cartilage remains unknown yet, but some proteinases seemed to prevent the matrix accumulation through degradation of synthesized proteins. In the present study, various kinds of proteinases, including MMP-1, -2, and -3, and cathepsin B, were significantly expressed in nasoseptal 3D cells compared to the monolayer counterparts (Fig. 6A).
The proteinase expression was speculated to be induced by the artificial 3D environment around the chondrocytes that were physiologically embedded within the COL2 matrix but not COL1-based “atelocollagen gel.” It was reported that embedment of a murine articular chondrocytic cell line (MC615) in COL1 gel triggered the activation of MMP-13 expression. 23 MMP-13 (collagenase-3) very efficiently degrades fibrillar collagens such as COL2 and COL1, 24 which are expressed in articular cartilage in human osteoarthritis 25 or its experimental models.26–28 This suggested that excess production of such a proteinase was correlated with cartilage destruction. Similarly to MMP-13, the presence of COL1 has also been reported in osteoarthritic cartilage. 29 COL2 is the major protein of cartilage, but a switch from COL2 to COL1 may trigger the MMP-13 activation and its involvement in cartilage degradation. The murine chondrocytic cell line MC615 that mimics the phenotype of the articular cartilage showed high expression of MMP-13 under 3D COL1 gel culture and α1β1 integrin–mediated 3D collagen signaling responsible for MMP-13 expression. 23 The MMP-13 upregulation in the murine articular chondrocyte cell line was slightly discrepant to the present study in that the difference in expressions between the human nasoseptal 3D and monolayer cells was more pronounced in MMP-1. When the proteinase expressions were compared between the 3D cells and monolayer ones from the human nasal septum in the present study, each of MMP-1, -2, and -3, and cathepsin B, especially MMP-1, was prominently upregulated in the 3D cells that were exposed to the COL1 derivative than in the monolayer counterparts. However, only the MMP-13 expression showed no significant difference between the nasoseptal 3D cells and monolayer counterparts.
We considered that the discrepancy was due to the difference in the cell origins. Both the nasal septum and the articular cartilage are classified as hyaline cartilage. The former is permanent cartilage in which the mature chondrocytes hardly become hypertrophic in any pathological conditions. On the other hand, the articular chondrocytes occasionally become hypertrophic or suffer from endochondral ossification under some pathological conditions, although the articular cartilage also belongs to the permanent type. The previous findings about osteoarthritis showed that some human articular chondrocytes began to hypertrophy and expressed COL10A1. 30 The MMP-13 in the articular chondrocytes principally follows such chondrocyte hypertrophy or COL10A1 expression. 28 The nasoseptal chondrocytes may hardly hypertrophy, express COL10A1 or subsequently produce MMP-13, unlike the articular chondrocytes.
Although a marked decrease in MMP-13 synthesis was observed when the cells were cultured in the presence of 100 ng/mL BMP-2 in the MC615 study, 23 matrix degradation and reduction of tissue-engineered cartilage pellets were exaggeratedly observed in the human nasoseptal 3D cells, even with the stimulation of not less than 200 ng/mL BMP-2 in addition to insulin, suggesting the abundant production of proteinases in human nasoseptal 3D cells. Such induction of proteinases may be related with some inflammatory cytokines. Bovine articular chondrocytes seeded in COL1 scaffolds responded to IL-1 and enhanced the expression of MMP-1, -3, and -13, and ADAMTS-4 and -5. 19 Also in the present study, TNF-α was significantly secreted in the nasoseptal 3D cells compared with the monolayer counterpart, while the inflammatory cytokine may promote the expression of various proteinases, especially MMP-1. In contrast, the auricular chondrocytes under the 3D condition did not show any upregulation of the MMP-1 expression, compared with those under the monolayer. This contradiction could be explained by the difference in the extracellular matrices physiologically surrounding each kind of cell. Although the nasoseptal chondrocytes, which originate from hyaline cartilage, are principally separated from the COL1 proteins under the physiological situations, auricular chondrocytes derived from elastic cartilage are intermixed with the elastic fibers or remnants of the COL1. 31 The nasoseptal chondrocytes appeared sensitive against the COL1 exposure and showed the upregulation of the MMP-1 expression. Otherwise, the auricular chondrocytes may possess a resistance to the COL1 exposure and could prevent the exaggerated production of various proteinases even under the COL1 3D culture conditions.
Like the proliferation ability, we speculated that these differences in the gene expressions and protein productions between the auricular chondrocytes and nasoseptal ones may not be based on the ages of donors. Various changes including a drastic downsizing of the cartilage pellets, decreased matrix accumulation, and extreme upregulation of the MMP-1 expression in the nasoseptal chondrocytes were found only in the 3D culture, and not in the monolayer one. If these might be caused by the general decrease in the matrix production of the chondrocytes originating from the aged population (donors of nasal septum), such changes should be noticed to some extent in the nasoseptal chondrocytes also under monolayer culture. Therefore, we considered that the changes in the nasoseptal 3D cells were due to the biological specificities for their cell origins.
To meet the increasing medical needs for cartilage tissue engineering, we should introduce greater flexibility into cell sources and culture methods. According to our data using the chondrocytes from donors who were matched in ages as closely as possible, the auricular chondrocytes are speculated to be more suitable for this 3D proliferation culture than the nasoseptal ones, because the tissue-engineered cartilage pellets of nasoseptal 3D cells significantly shrunk. However, the nasoseptal monolayer cells produced pellets with abundant GAG content. Accumulation of GAG may improve the mechanical properties due to the increase in hydrophilicity and may help to maintain the character of permanent cartilage by the inhibition of ectopic calcification. 32 Those cell characters of nasoseptal chondrocytes would offer some advantages when they are cultured in the monolayer culture. Therefore, we think that we should obtain many different cell sources for cartilage tissue engineering and choose them according to their chondrocyte specificity resulting from the difference in the donor sites or culture methods. Further investigation on the chondrocyte specificity resulting from the difference in donor sites or culture methods is needed to increase the options of chondrocyte sources in cartilage tissue engineering and to widen the indication range of cartilage diseases that can be treated by tissue engineering technology.
Footnotes
Acknowledgments
We thank Mr. Motohiro Harai and Ms. Yoko Isogai for technical support and useful discussion. This work was supported by Grants-in-Aid for Scientific Research from the Japanese Ministry of Education, Culture, Sports, Science, and Technology (MEXT, #15390539, #16390431, and #16659546), and Research and Development programs for Three-Dimensional Complex Organ Structures from New Energy and Industrial Technology Development Organization (NEDO), and for Resolving Critical Issues from Special Coordination Funds for Promoting Science and Technology (SCF) commissioned by MEXT.
Disclosure Statement
No competing financial interests exist.