
Abstract
Abstract
Exploiting embryonic stem cell (ESC)-derived cardiomyocytes as a vital source for cell therapies and tissue engineering will depend on robust, large-scale production processes. Recently, we have reported stirring-controlled formation of embryoid bodies, enabling the generation of pure cardiomyocytes in 2-L scale. Expansion and differentiation of genetically engineered mouse ESCs was followed by antibiotic-based cardiomyocyte enrichment. Here we have investigated modification of various parameters aiming at process optimization in a 250-mL spinner flask system. Duration of the differentiation phase, timing of retinoic acid addition, and a slower medium exchange rate were found to be crucial to enhancing cardiomyocyte yield. Improved process conditions were consequently transferred to a 2-L controlled bioreactor. Employing a manual fill-and-draw medium change resulted in the formation of 0.86 × 109 cardiomyocytes in a single 2-L batch, thereby reproducing our previous findings. In contrast, an automated perfusion-based strategy enabled the production of 4.6 × 109 cardiomyocytes in a single run. This is significantly higher than previously reported and highlights the enormous process optimization potential in the scalable generation of ESC-derived cell lineages.
Introduction
However, the differentiation of cardiomyocytes from bone marrow cells after myocardial transplantation was questioned,12,13 and the functional coupling of skeletal myoblast with heart muscle cells post engraftment was denied. 14
ESCs, on the other hand, have several limitations regarding their clinical application, with immunogenicity and the inherent potential to form teratomas being most prominent.15,16 Given that these issues have been comprehensively investigated in long-term animal studies, their enormous proliferation potential and their ability to form transplantable bona fide cardiomyocytes in vitro make ESCs a promising raw material for replacement therapies. 17 For most future cell therapy applications and medium- to high-throughput drug screens, large numbers of selected cell types without contamination from other lineages will be required. Here, we describe a method by which this can be achieved for cardiomyocytes from mouse ESCs (mESCs). The methodology developed may later be applicable to the more clinically relevant human ESCs (hESCs) and other differentiated derivatives.
Laboratory-scale suspension cultures in hanging drops or Petri dishes are useful tools for process development and initial optimization,18–21 but these approaches are not suitable for further scale-up. We recently described a 2-L bioreactor process enabling the controlled generation of embryoid bodies (EBs) directly from a single-cell suspension. Our system included automated process control and online measurement and adjustment of culture parameters. 22 To achieve pure cardiomyocyte cultures, we took advantage of an ESC line designated CM7/1, which permits efficient purification of cardiomyocytes.20,21 CM7/1 cells carry a transgene comprising the cardiac-restricted alpha major histocompatibility complex (αMHC) promoter and a neomycin resistance cassette. For cardiomyocyte enrichment, a selection medium was added for 9 days after a 9-day-long expansion and differentiation phase. This process readily yielded 1.2 × 109 essentially pure cardiomyocytes in a single 2-L batch.
Herein we report our current investigations in a 250-mL spinner flask and the 2-L controlled bioreactor system, aiming at enhanced cardiomyocyte production. To this end, duration of the expansion and selection phases was modified, and a time- and concentration-dependent effect of retinoic acid (RA) was studied. Finally the fill-and-draw feeding protocol was replaced with a continuous perfused culture, which allowed constant medium supply and avoided daily fluctuations of medium components. An optimized protocol resulted in more than five times greater cardiomyocyte yield, whereas medium consumption was 40% less in parallel.
Materials and Methods
Media and cell cultivation
ESC expansion medium consisted of N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid–buffered Dulbecco's modified Eagle medium (Invitrogen, Karlruhe, Germany), 15% fetal calf serum (FCS; PAA, Linz, Austria), 0.1 mM 2-mercaptoethanol (Sigma, Taufkirchen, Germany), 0.1 mM NAA (Invitrogen), and leukemia inhibitory factor (LIF, 1000 U/mL; Chemicon, Hofheim, Germany). For differentiation, LIF was omitted, and the FCS content was reduced to 10% (differentiation medium). Selection of cardiomyocytes was initiated with Geneticin (G418, 400 mg/mL; Invitrogen) and all-trans RA (10 nM; Sigma). Addition of Antifoam C (0.025% v/v;Sigma,) was necessary for all bioreactor cultivations. Cultivation of J1-ESCs and generation of the genetically engineered clone CM7/1 were performed as previously described. 21
Spinner cultures
For inoculation, CM7/1 ESCs were washed with phosphate buffered saline (PBS) (Biochrom, Berlin, Germany) and dispersed into single cells using 1 × trypsin– ethylenediaminetetraacetic acid (EDTA; Sigma). After resuspension in differentiation medium, spinner flasks equipped with a bulb-shaped glass stirrer (Integra Biosciences, Fernwald, Germany) were inoculated with 125 mL of medium containing 2 × 105 cells/mL and stirred at 60 revolutions per minute (rpm). Flasks were filled up to 250 mL after 48 h, followed by a daily half medium exchange. Cardiomyocyte selection was initiated by adding Geneticin (G418, 400 mg/mL) to eliminate non-cardiac cells. RA was supplemented at 1 to 100 nM final concentration. Three-mL samples for cell and EB count were taken daily, and metabolic analysis of the medium was also performed daily. Three independent spinner runs were performed for every experimental setting.
Controlled bioreactor cultures
A suspension of 2 × 105 cells/mL in differentiation medium was seeded into the 2-L stirred bioreactor Biostat MD (Sartorius BBI Systems, Melsungen, Germany; height to diameter relation of the vessel, 2:1; vessel diameter, 130 mm). An 8-bladed pitched-blade turbine, engineered as previously described at the Institute of Cell Culture Technology, University of Bielefeld, Germany, was used for stirring. 22 The water-jacketed bioreactor was equipped with a microsparger; temperature sensor; pH and partial pressure of oxygen-electrodes; and inoculation, harvest, and sample ports. Data acquisition and process control were done with a digital control unit and MFCS software (Sartorius BBI Systems). After trypsination, 2 × 108 cells (2 × 105 cells/mL) were seeded into 1 L of differentiation medium, and 2 days after inoculation, the volume was adjusted to a final suspension volume of 2 L. Aeration was performed via head space for the first 2 days of differentiation and switched to sparged aeration afterwards. To prevent foam formation, Antifoam C was added at a final concentration of 0.025% (v/v). Dissolved oxygen was controlled at 40% with air or oxygen supply, pH was controlled at 7.2 through addition of carbon dioxide, and temperature was set at 37°C. Stirring was performed at 65 rpm. For manual medium exchange in the fill-and-draw process, EBs were allowed to settle down by gravity for 10 min before medium replacement.
Using a Watson Marlow pump (Rommerskirchen, Germany) (101 U, outlet flow), continuous medium perfusion was implemented by attaching a settling tube to the feed outlet. The diameter of the settling tube (15 mm) was specified so that the velocity of the medium outlet was slower than the minimum EB settling velocity. By replacing one-quarter reactor volume per day (0.5 L medium), a medium outlet velocity of 13.5 cm/h was obtained, which was slower than the settling velocity of d3 EBs (46 cm/h). Differentiation medium was fed with a Watson Marlow 503 U pump and regulated using a level sensor, which activated the feeding pump whenever contact between suspension surface and sensor was interrupted. Samples (5 mL) were taken aseptically before and after medium exchange for quantitative analysis. For cardiomyocyte enrichment selection, medium was added after 9 and 11 days of differentiation depending on the experimental set-up. Selected cells were cultivated for additional days to sustain a pure cardiomyocyte population. Contracting spheres generated upon selection were termed cardiac bodies (CBs). For process optimization, we analyzed the effect of the following culture protocol changes: duration of expansion phase, time-dependent addition of RA, concentration-dependent addition of RA, and feeding strategy.
Analytical methods
Cell count
To obtain a single cell suspension, EBs and CBs were washed with PBS (Biochrom) for 30 min at room temperature and incubated with 1 mL trypsin–EDTA (Sigma) at 37°C for another 10 to 30 min followed by cautious mechanical dissociation. Count of viable cells was determined using a hemocytometer and trypan blue staining.
Measurements of glucose and lactate concentrations were performed using an automatic analyzer (YSI 2700 Select, Yellow Springs Instruments).
Immunocytochemistry and confocal laser scanning microscopy
Cardiac body-derived cells were seeded on fibronectin (Roche Diagnostics, Mannheim, Germany)-coated Permanox chamber slides (Nunc, Wiesbaden, Germany), and staining was performed 3 to 7 days after plating. Briefly, cells were fixed in ice-cold methanol for 5 min, followed by permeabilization in acetone for 3 min at −20°C. After rehydration, specimens were blocked in 10% donkey serum in PBS for 30 min. The slides were incubated with the monoclonal antidesmin antibody RD301 (Dianova, Hamburg, Germany; 1:100); the polyclonal rabbit anti-desmin serum (Progen, Hamburg, Germany; 1:100); the monoclonal antibodies against α-actinin, tropomyosin, and rabbit anticonnexin-43 (all from Sigma-Aldrich, Taufkirchen, Germany; 1:10, 1:800 and 1:50, respectively); monoclonal anti-cardiac-α-actin; anti-desmoplakin 1 & 2 (both from Progen; 1:10 and undiluted, respectively); or anti-titin 9D10 (a generous gift from W. Linke, Germany; 1:10) for 60 min at room temperature. After thoroughly rinsing in PBS, a Cy-3 labeled donkey-antimouse antibody (Dianova, Germany) and Alexa 488 labeled donkey–anti-rabbit antibody (Invitrogen, Germany) were applied for 30 min together with 4,6-diamidino-2-phenylindole (Roche Diagnostics) for nuclear staining. The slides were finally mounted with cover glasses in Fluoromount G (Southern Biotechnology Associates, Birmingham, AL). Cells were viewed using confocal laser scanning fluorescence microscopy (DMIRE 2, Leica, Wetzlar, Germany).
Statistics
All statistical analyses were performed using Origin 7.5 graphing and data analysis software (OriginLab, Northampton, MA). All results from the spinner runs, generated from three independent experiments, were analyzed using a significance level of p = 0.05. Values are presented as means ± standard deviations.
Results
Process optimization in 250-mL spinner scale
In a previous study, we established a 2-L bioreactor process enabling stirring-controlled EB formation from a single ESC suspension. Concomitant differentiation and selection yielded more than 109 essentially pure cardiomyocytes in a single 2-L batch. 22 To ascertain further optimization, we employed spinner flask cultures in 250-mL scale, a meaningful system to test multiple parameters in parallel before further scale-up. Although numerous additional process variables such as glucose concentration and time-dependent RA-supplementation were tested, we will focus on conditions that ultimately resulted in higher overall cell counts and better cardiomyocyte yield. Our former bioprocess strategy (termed BPS control) comprised a 9-day-long expansion and differentiation phase, followed by 9 days of G418 treatment for cardiomyocyte enrichment RA, known to enhance cardiac differentiation and deplete undifferentiated ESCs,19,22 G418 was supplemented at 10 nM from day 9 or in combination with an antibiotic-based cardiomyocyte selection. During the whole process, 50% of the medium was manually replenished every 24 h.
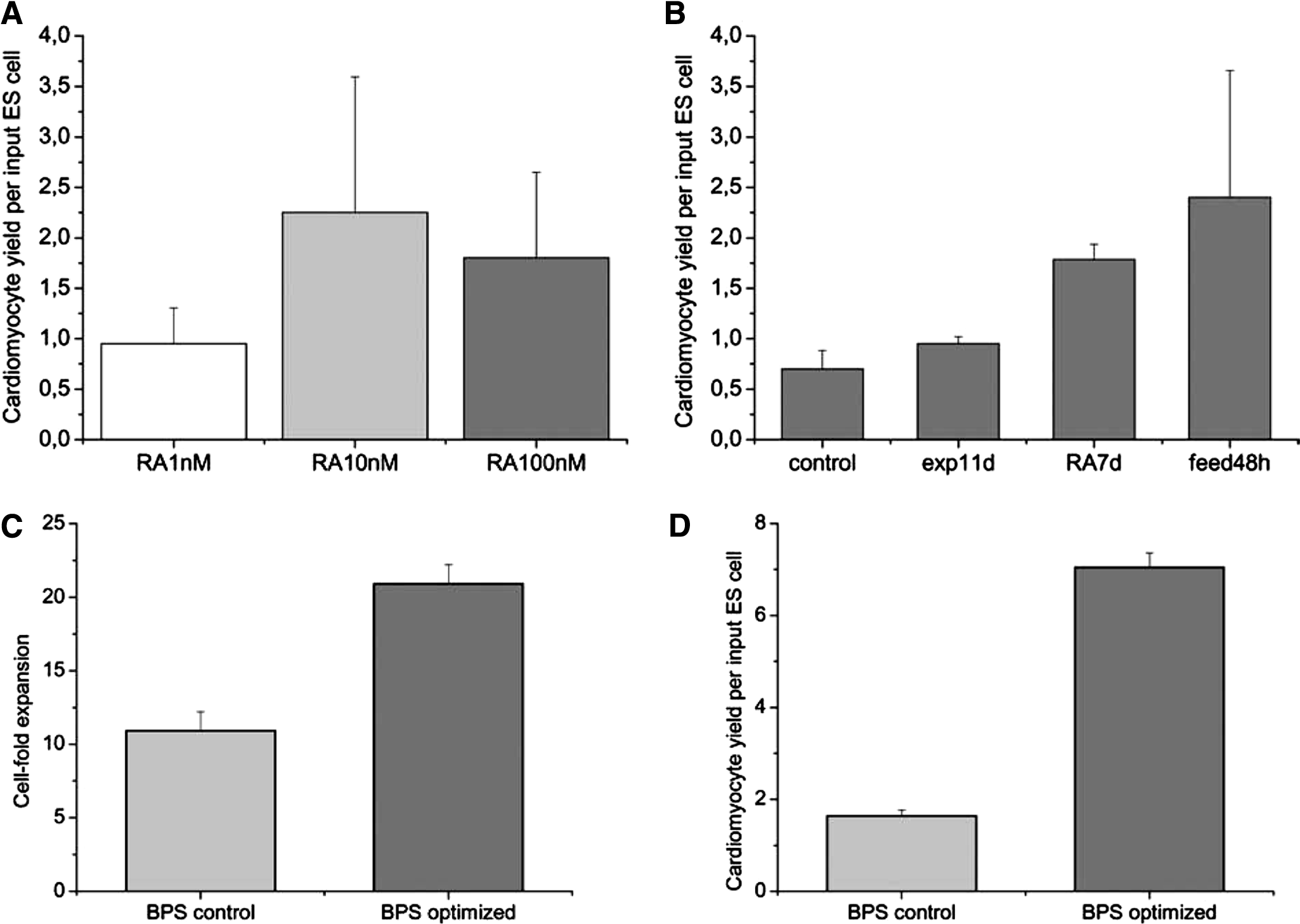
In a first step, the concentration- and time-dependent effect of RA supplementation was investigated. By testing a 1- to 100-nM RA range, we noted that the former 10 nM concentration proved to be optimal with respect to cardiomyocyte production in our stirred system. The cardiomyocyte output can be specified according to a coefficient comprising the number of cardiomyocytes generated per input ESC (CM/ESC). Addition of 10 nM of RA resulted in 1.5 × 105 cardiomyocytes after Geneticin selection, corresponding to a CM/ESC coefficient of 2.3, whereas addition of 1 nM and 100 nM, respectively, led to CM/ESC coefficients of 1.0 and 1.8, respectively (Fig. 1A). The beneficial effect of 10 nM of RA on cardiac differentiation confirms previous findings in static hanging drop cultures.23,24However, earlier addition of RA in BPS modified on day 7 instead of day 9 resulted in a CM/ESC coefficient of 1.8 (RA7d; Fig. 1B), whereas addition of RA on day 5 yielded lower cardiomyocyte numbers, resulting in a CM/ESC coefficient of only 0.42 relative to BPS control.
(
Next, we extended the duration of the expansion and differentiation phase before applying cardiomyocyte enrichment. The enrichment phase was shortened, and the overall culture period was retained for all experiments. Previous experiments suggested that cells in EBs still proliferated at day 9. 22 Indeed, higher cell densities were achieved with a prolonged, 11-day-long differentiation phase, resulting in 3.0 × 105 cells/mL after 9 days but 3.8 × 105 cells/mL after 11 days of EB culture. Higher cell densities also translated into a better CM/ESC coefficient in the modified process. This coefficient was 1.3 times higher for modified BPS than for BPS control (exp11d; Fig. 1B). Taken together, after modifying the original process strategy, the optimal feeding protocol was found to be an 11-day expansion phase with the addition of 10 nM of RA on day 7.
After optimization of these factors, we focused on the feeding protocol. A fill-and-draw strategy was used in the control process to replenish half of the medium daily. The feeding protocol was adapted from published protocols for non-ESC cell lines,25–28 being designed for optimal nutrient supply and preventing accumulation of toxic metabolites even at high cell densities in suspension. However, this regimen also results in a severe dilution of cell-secreted factors that might influence cell proliferation and differentiation. Thus, an alternative strategy was tested, and 50% of the medium was replaced only every 48 h. Daily monitoring revealed that glucose levels did not drop below 4.4 mM, and lactate did not render toxic. The highest lactate concentration of 25.6 mM measured on day 9, before cardiomyocyte selection was initiated. Cultures that were fed every second day resulted in a CM/ESC coefficient of 2.4 on day 9 (feed48 h; Fig. 1B). These results indicate that a lower medium turnover was advantageous with respect to overall cell and final cardiomyocyte yield, providing important data toward establishing a perfusion-fed process.
In essence, a new process strategy termed BPS optimized (overall 16 days: 11-day expansion + 5 days of selection) was defined and compared with BPS control (16 days: 9-day expansion + 7 days of selection). Although inoculation was unchanged (2 × 105 cells/mL in 125 mL), medium was adjusted to 250 mL after 48 h, cell feeding was performed only every second day for BPS optimized, RA was added from day 7 on, and EBs were cultured for 11 days before cardiomyocyte selection was initiated via G418 addition.
Immunocytochemistry specific to cardiac proteins unraveled the reliable elimination of non-cardiomyocytes from the culture after antibiotic treatment for 5 days. Harvested aggregates were enzymatically dispersed and plated in a density that enabled single-cell examination. Analysis of 1 × 106 revealed that 99% (±1) of the cells were positive for cardiac α-MHC or Troponin T-C (data not shown). Figure 1C documents a greater overall cell expansion before selection in the optimized process (20.9-fold ± 1.3 vs 10.9-fold ± 1.3). Furthermore, BPS control yielded 4.5 × 107 (±1.1 × 104) cardiomyocytes, but 19 × 107 (±2.9 × 104) cardiac cells were generated in the newly designed protocol, translating into a 4.3-fold higher CM/ESC coefficient in BPS optimized (Fig. 1D).
Optimized cardiomyocyte generation in a bioreactor system with automated process control
Based on the spinner flask results, alterations of the process strategy were translated to the 2-L scale system, which allows full process control. As opposed to a 9-day-long expansion and differentiation phase followed by 9 days of selection, the spinner experiments suggested an extended expansion time of 11 days followed by a cardiomyocyte harvest on day 16 after 5 days of selection. Ten nM of RA were supplemented from day 7 on instead of day 9, according to the results given above. To ensure more-constant culture conditions, a major goal in the 2-L reactor was to implement a continuous perfusion system. Perfusion was enabled by employing a settling tube for cell retention in the reactor. In our published 2-L fill-and-draw process, 1 L of medium was replenished daily, resulting in a dilution rate of 0.5d−1. 22 However, our spinner flask results suggest that a lower medium throughput might be favorable for cardiomyocyte yield. Thus, a perfusion rate of 0.5 L/d, corresponding to a dilution rate of 0.25d−1, was chosen, and the newly established protocol, termed bioreactor (BR) optimized, was compared with the previously used strategy BR control. 22
CM7/1 cells were inoculated into the Biostad MD reactor system at a density of 2 × 105 cells/mL in 1-L differentiation medium, and adjustment to the final volume of 2 L followed after 48 h. Before selection, cell densities of 6.0 × 106 cells/mL (12 × 109 cells/2 L; day 9; BR control) and 4.9 × 106 cells/mL (9.8 × 109 cells/2 L; day 11; BR optimized) were observed (Fig. 2A, B). Addition of G418 initiated elimination of non-cardiomyocytes. As a result, cell numbers dropped markedly within 48 h but stabilized until day 16, yielding 0.43 × 106 cells/mL (0.86 × 109 cells/2 L) in BR control and 2.3 × 106 cells/mL (4.6 × 109 cells/2 L) in the optimized process (Fig. 2A, B).
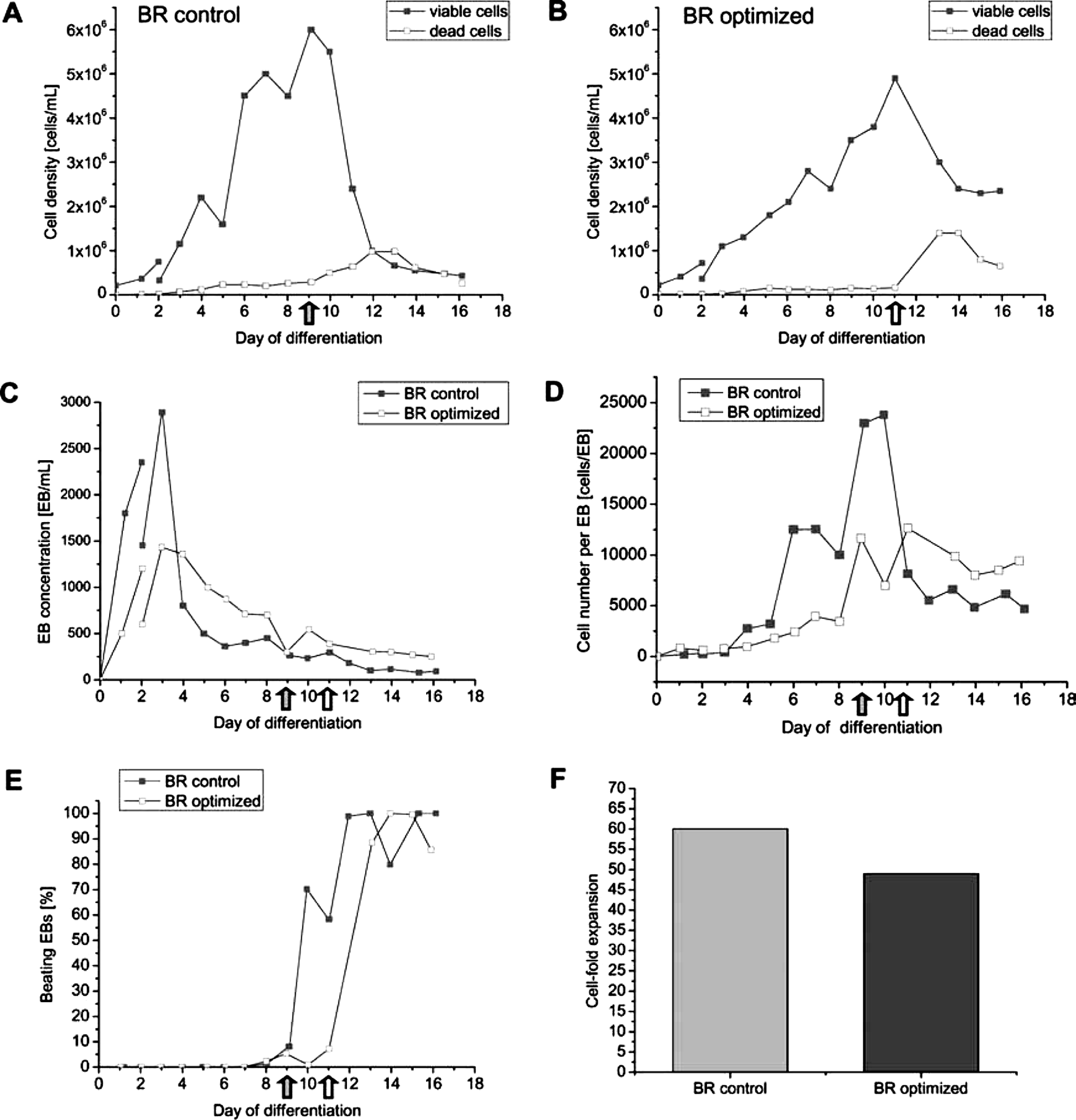
Process development in the fully controlled 2-L bioreactor (BR). Embryonic stem cells (ESCs) were seeded into 1 L of differentiation medium, and on day 2 after inoculation, the volume was adjusted to the final 2-L volume (indicated as a break in the curves describing cell and embryoid body (EB) yields in
After a peak, on day 3, EB concentrations showed a marked drop at day 4 of differentiation, but EB numbers concomitantly stabilized between days 6 and 8 in both processes (Fig. 2C). This pattern might reflect the known effect of EB agglomeration in dense suspension cultures.22,29 In addition, high numbers of small cell aggregates on days 1 through 3 might result in an initial overestimation of the EB concentration. However, once stabilized, EB numbers were constantly higher in BR optimized (e.g., 450 EBs/mL and 699 EBs/mL on day 8, respectively; Fig. 2C), suggesting that the perfusion feeding strategy might have limited EB agglomeration to some extent. The EB size distribution shown in Fig. 2D supports this interpretation. Before selection, EBs in BR optimized were constantly smaller in BR optimized, but higher cell numbers per aggregate were found after selection. After the addition of G418, EB numbers dropped as expected and stabilized to approximately 90 (BR control), versus 250 EBs/mL (BR optimized) on day 16. In both processes, first contracting EBs were observed on day 7. Because of the selection step, most if not all clumps remaining in the culture at the end of the process were vigorously beating (Fig. 2E) and were thus termed CBs (CBs).
Overall cell expansion before selection was higher in BR control, resulting in a multiplication ratio (MR: total vital cell count divided by the number of ESCs used for inoculation) of 60 (day 9) versus MR 49 in BR optimized (day 11, Fig. 2F). In strong contrast, the optimized process finally resulted in the formation of 4.6 × 109 cardiomyocytes/2 L (vs 0.87 × 109 cardiomyocytes/2 L BR control) and thus an approximately 5.3-fold higher cardiomyocytes yield per input ESC than in BR control (23 vs 4.3 C
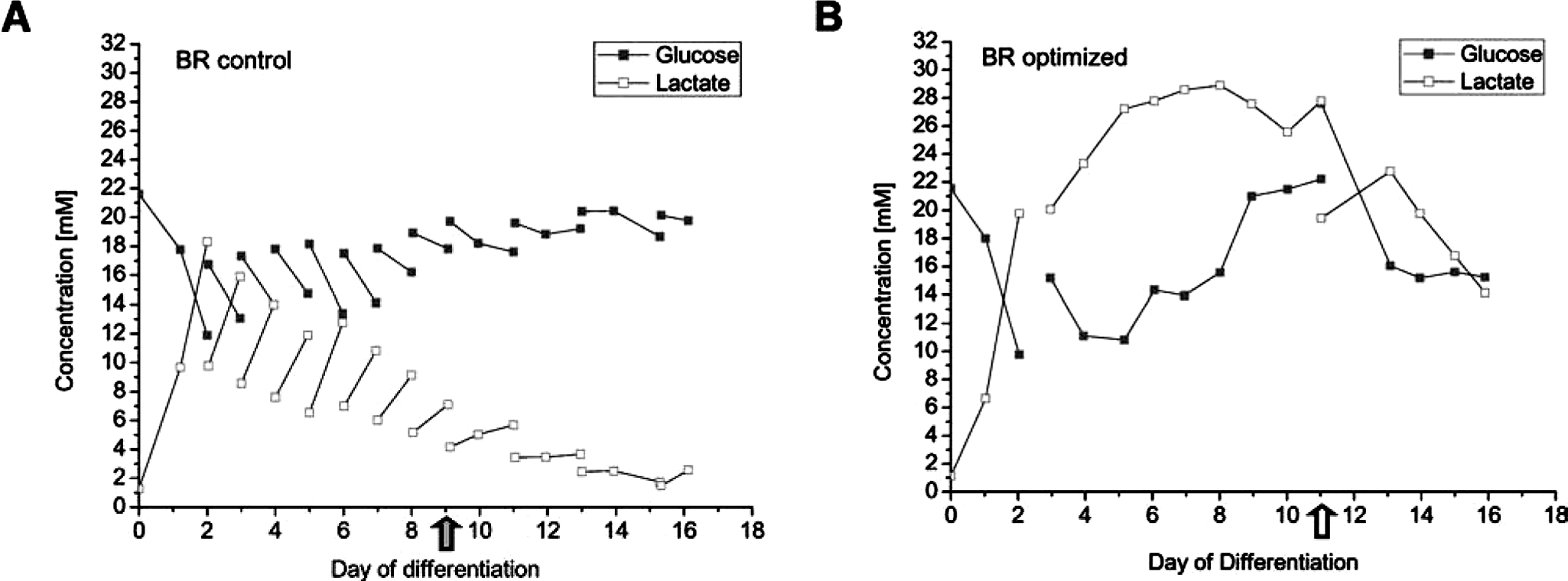
For process control, several culture parameters such as glucose and lactate concentration were monitored daily. As expected, higher lactate concentrations were measured in BR optimized according to the lower medium dilution rate of 0.25d−1 in the optimized process, compared with 0.5d−1 in BR control. Despite the high cell densities of 5 to 6 × 106 cells/mL obtained in both bioreactors, lactate concentration did not reach toxic levels at any time. The highest concentration measured was 28.9 mM (Fig. 3B) which is comparable with the level observed in our previous process. 22 Studies on extensively investigated cell lines such as CHO, BHK, and hybridoma cells have shown that proliferation rates are unaffected at a lactate concentration of up to 50 mM, especially in a pH-controlled environment, as we have applied.25–28 Because glucose concentration also did not drop below 10 mM (Fig. 3A, B), we are assuming that neither nutrient supply nor toxic metabolites limited ESC growth and differentiation in either process.
(
Immunocytochemistry and confocal microscopy confirmed cardiomyocyte purity and formation of characteristic myofilaments.
Immunocytochemistry was used to characterize G418-selected cells and to demonstrate purity of the thereby enriched cardiac cell population. Selected cultures were therefore digested with trypsin and re-plated at a density enabling analysis of individual cells. Seeded cells exhibited a typical cardiac morphology, with highly organized titin, cardiac α-actin, tubulin, and α-actinin structures. The extra-sarcomeric cytoskeletal protein desmin was found to form extended filamentous networks. Desmoplakin staining revealed the presence of intercellular desmosomal junctions (Figs 4A–F, H). Likewise, the presence of gap junctions between neighboring cells was documented using connexin43 immunostaining (Fig. 4G). 30 The incidence of cells that did not show immunoreactivity specific to cardiac markers, in particular to extensively applied α-MHC immunoreactivity (in combination with a peroxidase-coupled secondary antibody and 3,3-diaminobenzidine staining, data not shown) was less than 1%, in agreement with previous findings.21,22 In summary, these results strongly suggest the generation of essentially pure bona fide cardiomyocytes from genetically engineered CM7/1 ESCs in our large-scale process.
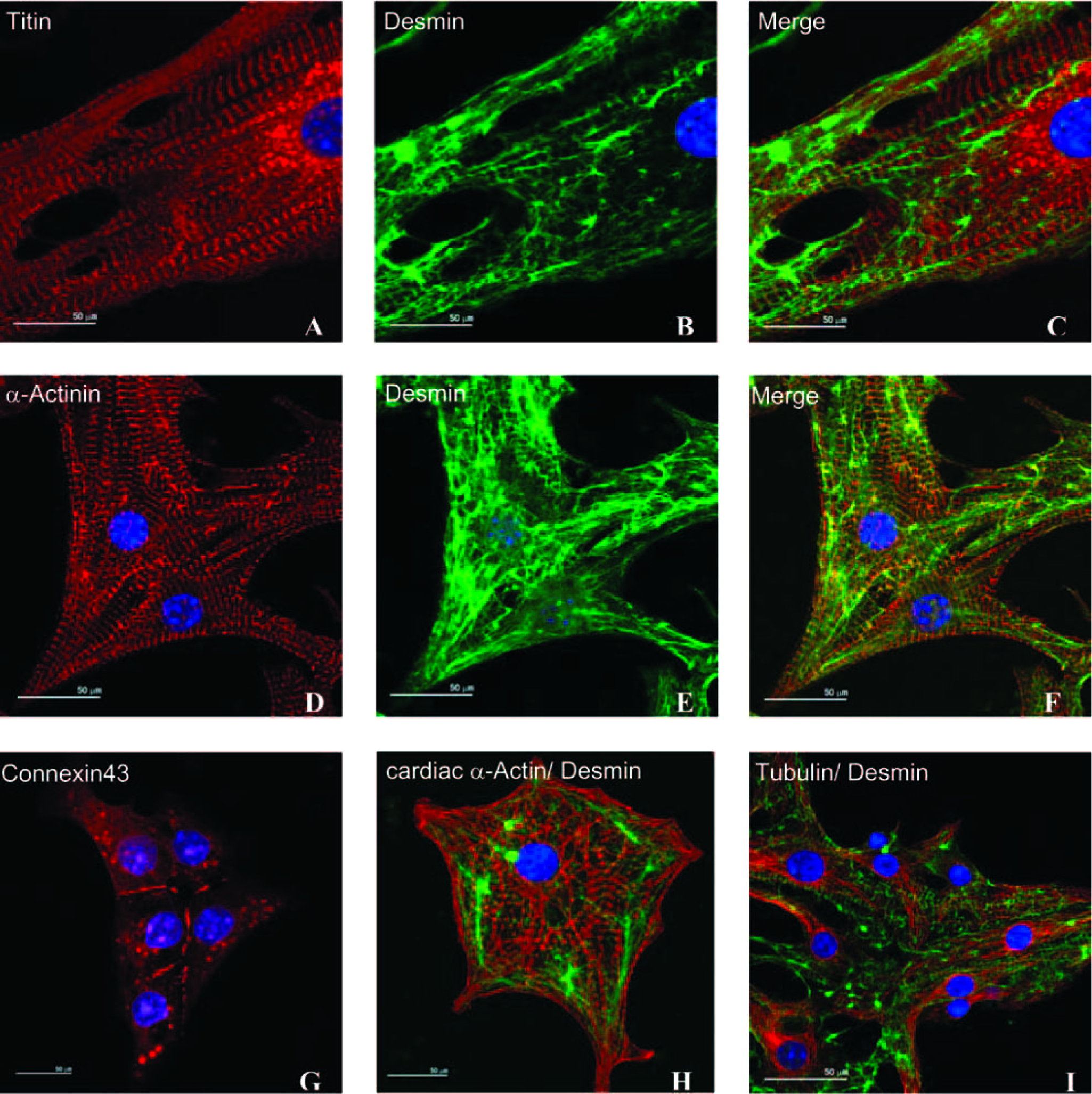
Double fluorescence labeling of bioreactor-generated cardiomyocytes. Cardiac bodies were dissociated to single cells, plated on chamber slides, and cultured for an additional 3 days before staining. Well-developed myofibrillar structures were detected using titin (
Discussion
We and others have recently demonstrated the successful differentiation of mESCs in suspension culture to provide large numbers of pure cardiomyocytes.20,22,31 Optimized process inoculation and stirring conditions enabled the generation of 1.28 × 109 essentially pure cardiomyocytes in a single 2-L bioreactor run applying a fill-and-draw feeding strategy.20,22 Targeting greater cardiomyocyte yield, we here focused on other process parameters. Spinner flask experiments in 250-mL scale indicated that prolonged differentiation, earlier supplementation of RA, and reduced medium throughput are favorable for cardiomyocyte production. By translating these modifications to the 2-L system, we have also introduced another key parameter: continuous perfusion feeding. The resulting process leveraged the final yield to 4.6 × 109 cardiomyocytes in a 2-L reactor run, a yet unmatched value that underscores the enormous process optimization potential in ESC-derived cell lineage production.
Cardiomyocyte generation from ESC crucially depends on three factors: achieving a high overall cell density, inducing efficient cardiomyogenic differentiation in EBs, and facilitating potential further expansion of cardiomyocytes or respective progenitors. After inoculation with 2 × 105 ESCs/mL, highest cell densities of 4.9 × 106 cells/mL (BR optimized; Fig. 2B) and 6.0 × 106 cells/mL (BR control; Fig. 2A) were achieved. Higher cell densities in BR control did not translate into higher cardiomyocyte numbers. In contrast, the highest yield of 4.6 × 109 cardiomyocytes (23 CM/ESC) was observed under perfusion conditions, coversus 0.86 × 109 cells (4.3 CM/ESC, one-fifth as many) in the fill-and-draw process. These data might indicate that the perfusion feeding strategy improved cardiac differentiation. Alternatively, or in addition, the collective modifications in BR optimized (i.e., perfusion feeding, lower medium exchange rate, prolonged differentiation time, earlier addition of RA) might also support a greater expansion of pre-induced cardiomyocytes or progenitors.
Although single bioreactor runs were performed in this study, this is an accepted practice in large-scale bioprocesses development because of the expenditure of material requirements. It is therefore worth considering our former study in which we obtained similar growth kinetics, EB concentration, and cardiomyocyte yield in a fill-and-draw process equivalent to BR control. For example, the highest cell density of 6.9 × 106 cells/mL and a final yield of 0.64 × 106 cardiomyocytes/mL was obtained previously. 22 Respective values of 6.0 × 106 cells/mL and 0.43 × 106 cardiomyocytes/mL were observed in BR control, underscoring the reproducibility and robustness of the overall process conditions.
In both 2-L scale reactor runs documented here, EB numbers decreased between days 3 and 5, potentially as result of EB agglomeration, a known phenomenon induced by the adhesion of undifferentiated mESCs due to high levels of E-cadherin expression.18,31–34 The drop in EB concentration was most prominent in BR control (Fig. 2C). Our impeller design and optimized agitation allowed for the control of EB formation and agglomeration, resulting in well-defined EB dimensions. 22 However, the mandatory, daily EB collection in the fill-and-draw process might facilitate uncontrolled aggregation due to the high, transient EB density upon medium change. Automated perfusion feeding overcomes this limitation, thus further facilitating process reproducibility.
Perfusion systems are designed to reduce the severe daily fluctuation of medium components and cell-secreted molecules such as metabolites and growth factors.35–39 More-homogeneous culture conditions might support better-defined ESC differentiation. In vitro differentiation of ESCs is notoriously variable because of the ongoing changes in cell density, paralleled by the occurrence of differentiating cell lineages and thus changes in cell physiology, cell–cell interactions, growth factor secretion, etc. Inhibition of ESC differentiation after a noncontinuous, daily medium exchange was described by Viswanathan 34 as cell-secreted factors were diluted. It has also been reported for human bone marrow cells cultured in perfusion reactors that the consumption and release of a variety of growth factors can affect the cell type generated. 40 Perfusion-feeding further ensures optimal process uniformity with respect to temperature, pH, and gas flow, whereas manual medium exchange, at least transiently, encounters alternating pH and gassing conditions. For example, it has been suggested that high oxygen tension inhibits cardiac differentiation. In a study by Bauwens and coworkers, 31 a controlled, perfusion-fed system at a 250-mL scale was employed. The same cell line and similar differentiation and selection conditions as in our study were used, but EBs were formed from encapsulated ESCs. Highest cardiomyocyte yield was archived under hypoxic conditions (4% oxygen tension), resulting in a CM/ESC coefficient of 3.77, and a dramatically lower value of 2.56 CM/ESC was found at normoxia. Here, in BR optimized, a significantly higher CM/ESC coefficient of 23 at an oxygen tension of 40% was found; whether hypoxic conditions would further increase this value needs experimental evidence.
Defining the number of cardiomyocytes generated per input ESC is a useful parameter to assess process efficiency. Another coefficient, the number of cardiomyocytes generated per liter of culture medium throughput (CM/L), is of particular relevance for process scale-up if expensive, growth factor–supplemented media are used. In BR control 16-L medium throughput (2 L until day 2, 1 L × 14 days) resulted in the total generation of 0.86 × 109 cardiomyocytes, and thus a coefficient of 0.54 × 108 CM/L was calculated. The reduced medium throughput of 9 L in BR optimized (2 L until day 2, 0.5 L × 14 days) combined with the total yield of 4.6 × 109 cardiomyocytes added up to 5.1 × 108 CM/L. Whereas the CM/ESC coefficient was 5.4 times higher in BR optimized, the second index, CM/L, was even 9.5 times greater.
We have also performed studies to further characterize identity and purity of enriched cardiomyocytes. Immunocytology of dissociated cardiac bodies detected expression of α-actinin, titin, cardiac-α-actin, tubulin, connexin43, and desmin, demonstrating a well-organized myofibrillar and intermediate filament structure typical of cardiomyocytes (Fig. 4).41–43 Regarding the purity of selected cultures, no tumor formation was found after transplantation of our bioreactor-derived cardiomyocytes into infarcted mouse hearts after permanent coronary artery occlusion (n = 13 mice, analysis at 28 days after cell transplantation 3 ). It was found that transplantation led to a long-lasting cardiomyocyte survival and improvement of left ventricle function potentially due to an improvement in tissue architecture.
Recently, several studies have reported cardiomyocyte generation from hESCs. In most studies EBs were employed to induce cardiomyogenesis.44–47 Cameron and coworkers 48 also published a first step toward hESC differentiation in impeller-stirred systems employing a 250-mL Spinner flask system. This and other approaches7,49–52 suggest that culture techniques previously developed for murine ESCs are transferable to their human counterpart, because basic mechanisms such as differentiation via EBs are similar. Consequent development leads toward combining ESC propagation and differentiation in a one-step process. zur Nieden et al. 53 recently described controlled expansion of mESCs in stirred suspension bioreactors Also, the propagation of hESCs as single cells rather than the typical colony scoring was described 54 and will facilitate scale-up of hESC expansion and EB formation from single cells. Merging these findings with our protocol will lead toward a universal process for large-scale generation of hESC-derived lineages.
Conclusion
In summary, we have established a continuously fed bioprocess strategy capable of providing 4.6 × 109 essentially pure cardiomyocytes in a single 2-L bioreactor run. Reduced medium throughput strongly increased process efficiency, enabling the generation of 23 CM/ESC and 5.1 × 108 CM/L. Harvested cells were characterized and exhibited both: not only typical properties of bona-fide cardiomyocytes, but also a high purity of this desired cell type. Besides providing valuable data for mESC differentiation in large scale, our strategy may also pave the way toward scalable production of hESC-derived specialized cell types.
Footnotes
Acknowledgments
Harald Bär acknowledges a grant from the German Research Foundation (DFG; BA 2186/2–1).