
Abstract
Different lag-time of pulse-released nerve growth factor (NGF) from genipin-crosslinked gelatin within polycaprolactone (PCL) conduits was evaluated in large-gap peripheral nerve repair. In this study, 10% (w/v) gelatin was mixed with NGF, crosslinked with 0%, 0.1%, 0.5%, and 1% (w/v) genipin, and then sucked into the wall of PCL conduits. These controlled-release nerve conduits were named NCL (non-crosslink), LCL (low crosslink), MCL (medium crosslink), and HCL (high crosslink), respectively. The NGF releasing character showed four distinctive curves, including initial burst within 5 days, pulse releasing at 5–20 days, pulse releasing at 10–25 days, and steadily releasing. The bioactivity of the released NGF was shown by neurite outgrowth of PC12 cells after culturing in all groups. Finally, the controlled-release conduits were seeded with 9 × 103 Schwann cells. Conduits were used to bridge a 15-mm rat sciatic nerve defect, and the results were compared with the isografts (control group). Eight weeks after implantation, morphological analysis revealed that LCL, MCL, and HCL groups were similar to autograft treatment in the numbers and area of myelinated axons. The LCL group, although insignificant, showed a trend to have the highest myelinated axon counts of the conduit-treated groups. Thus, comparing the different NGF release characteristics among NCL, MCL, and LCL groups, we concluded that a high concentration of NGF at 5–10 days in LCL groups is needed in bridging a 15-mm peripheral nerve injury.
Introduction
Numerous approaches have been utilized to deliver growth factors into the peripheral nerve conduit.11,12 Controlled releases of growth factors can be achieved by many methods, such as delivery from polymeric microspheres, discs, and mini-pumps.13–20 However, the controlled-release conduits could not achieve comparable results to autografts in bridging large-gap injured nerves over long repair periods. 21 This may be attributed to the unknown optimal dosage and present time of released NGF in nerve conduits. Therefore, a time-controlled pulse-releasing system is desired in the controlled-release nerve guide conduits. 22
Gelatin, which is essentially denatured collagen, has a myriad of uses in the food, pharmaceutical, and cosmetic industries.23–25 Gelatin does not express antigenicity in physiological conditions; it is completely resorbable in vivo, and its physicochemical properties can be suitably modulated. Further, it is much more inexpensive than collagen and easier to obtain in concentrated solutions.26–28
Genipin is a naturally occurring and low-cytotoxic crossing agent, which can be obtained from its parent compound geniposide isolated from the Gardenia fruit extract.29,30 Genipin has been widely used in herbal medicine, 31 and the dark blue pigments obtained by its spontaneous reaction with amino acids or proteins have been used in the fabrication of food dyes. 32 Recently, genipin has been demonstrated to possess neuritogenic activity in PC12h cells (a subclone of PC12 cells) 33 and neuroprotective activity in cultured rat hippocampal neurons exposed to β-amyloid peptide. 34
The aim of this investigation was to develop a delivery system that would provide different lag-time of pulsed release of NGF into injury sites for use in large-gap nerve regeneration. Various crosslinked degrees of gelatin were employed as NGF carriers to afford distinctive releasing characteristics of NGF, which included initial burst within 5 days, pulse releasing at 5–20 days, pulse releasing at 10–25 days, and steady releasing. Animal studies were conducted by implanting the controlled-release cell-seeded polycaprolactone (PCL) conduit into a 15-mm right sciatic nerve gap in rats. After 8 weeks, implanted nerve grafts were harvested and evaluated for the regeneration of the myelinated axons and vessels by histological examination. This investigation established that a high concentration of NGF at 5–10 days is needed in bridging a 15-mm peripheral nerve injury.
Materials and Methods
Fabrication of PCL porous nerve conduits
Two-ply nerve conduits were prepared by dip coating. Briefly, a 10% (w/v) solution of PCL (Sigma, Milwaukee, WI) was prepared in 1,4-dioxane reagent (Sigma). Salt crystals (100 μm) were added in a 30/70 (w/w) polymer/salt ratio. A glass mandrel was immersed in the salt suspension, and then pulled out many times to form the inner layer; 1,4-dioxane was allowed to evaporate at room temperature (18–25°C) for 24 h. The outer layer underwent a similar procedure with 37-μm salt crystal suspension added. Finally, the salt particles within the porous PCL conduit were leached out in water for 5 days (with daily renewal of the water) and then dried at room temperature. The dimensions of the fabricated conduits are as follows: inner diameter = 1.6 mm; outer diameter = 1.8 mm; length = ∼ 30 mm.
Preparation of controlled-release conduits
Type A gelatin (Bloom number 300; Sigma) was extracted and purified from the porcine skin with an average molecular weight of approximately 50,000–100,000 Dalton. A 10% (w/v) solution of gelatin in distilled water was prepared with magnetic stirring. The NGF was added to the gelatin solution with a concentration of 5000 ng/mL (7S NGF: Chemicon, Temecula, NJ). The aqueous solution was crosslinked with 0%, 0.1%, 0.5%, and 1.0% (w/v) genipin (Mw 226; Challenge Bioproducts, Taichung, Taiwan), and named NCL (non-crosslink), LCL (low crosslink), MCL (medium crosslink), and HCL (high crosslink), respectively. A 0.06 mL of resultant solution was injected into the porous PCL conduit twice, and then the two ends were sealed. The conduits were rotated horizontally at a constant rate to reduce variations of the infiltration of the gelatin. Conduits were placed in a vacuum pump at −760 torr for 2 h. Then, the outer surfaces of the conduits were washed using phosphate-buffered saline (PBS). Finally, the controlled-release PCL conduits, which were coated with genipin-crosslinked gelatin, were maintained at 4°C for use in subsequent experiments.
Morphology of PCL nerve conduits
The PCL conduits were gold coated to a thickness of about 500 × 10−8 cm using a Hitachi IB-2 coater in a high vacuum at 0.1 torr, at a high voltage of 1.0 kV, and at 50 mA. Samples were observed by scanning electron microscope (FE-SEM, JSM 1025; JEOL, Tokyo, Japan). Additionally, the porosity of PCL conduits was determined using the method in our previous study. 35 Each step in the measurement of porosity was carefully performed at least 10 times.
Characterization of NGF release
The release profile of NGF in PCL conduits was determined by incubating a 17-mm-long PCL conduit, coated with genipin-crosslinked gelatin, in 1 mL of PBS that was placed in an incubator at 37°C. In each time interval, the PBS buffer was collected and replaced with fresh PBS buffer. The collected PBS was stored at −20°C for subsequent analysis.36–38 An ELISA was used to determine the content of NGF in the collected PBS. The concentration of NGF in conduit lumen is accurately obtained by multiplying the data and PBS/conduit volume ratio (1 mL/34.16 μL = 29.27). The NGF antibody of the ELISA kit (Chemicon; NGF sandwich ELISA kit) specifically reacts with the β-subunit of NGF in the 7S form and detects the released NGF.
Bioactivity of released NGF
This investigation assesses the neurite outgrowth of PC12 cells to evaluate the bioactivity of NGF that is released from the genipin-crosslinked gelatin conduit. PC12 cells are known to differentiate to a neuronal phenotype in response to NGF. The controlled-release conduits were placed in the apical chamber of the transwell system (Corning Incorporated, Corning, NY) as a source of controlled-release NGF. PC12 cells were cultured in the basal chamber at a density 106 cells/well. The percentage of neurite-bearing cells was determined by counting 200 cells in several randomly chosen fields using a digital camera (COOLPIX 995; Nikon, Tokyo, Japan) and an image analysis system (Image-Pro Lite; Media Cybernetics, Bethesda, MD) for every sample in 1, 2, and 3 weeks of incubation. Neurite-bearing cells are those with processes greater than or equal to the diameter of the cell body. 39 In addition, the cells, which were shorter than 40 μm, had all been expelled. The −NGF (control group) was comprised of PC12 cells that were incubated with medium alone (Blank). The +NGF (positive control group) consisted of PC12 cells that were incubated with 20 ng/mL of NGF-supplemented cell culture medium.
Culture of Schwann cells
Schwann cells were obtained according to the method of our previous study. 40 Sciatic nerves of 3-day-old Sprague–Dawley rats were harvested and incubated in Liebovitz's L15 (Gibco, Biologic Research Laboratory, Grand Island, NY), and washed three times with PBS (pH 7.4). The epineurium was separated, and nerves were dissected into discrete fascicles (less than 1 mm), which were enzymatically dissociated using 0.03% type I collagenase (Sigma) and 0.25% trypsin in 10 mL PBS for 60 min at 37°C. The resulting cell suspension was filtered (44 μm; Millipore, Billerica, MA), centrifuged (1500 rpm; 10 min), and placed in a 35-mm culture dish. The purity of cultures was determined on the following day; 10 μM, 0.05 mL AraC solution was added to the culture medium to remove fibroblasts, and the medium was incubated for 4 days. The culture was plated on poly-L-ornithine–covered 25-cm2 flasks (2 × 105 cells/5 mL) in an incubator with 5% CO2 at 37°C. The culture was fed with Dulbecco's modified Eagle's medium (Hy-Clone, Logan, UT) supplemented with 1% (v/v) antibiotics (10,000 U/mL penicillin G and 10 mg/mL streptomycin), 2 mM glutamine, and 10% fetal calf serum (Biologic Industries, Kibbutz Beit, Haemek, Israel). Cultures were maintained under standard culture conditions for about 2 weeks until a confluent monolayer was obtained. The identification of Schwann cells was verified using the S-100 staining method, and the purity of the culture exceeded 95%.
Preparation of Schwann cell–seeded controlled-release conduits
Schwann cells were expanded for 8 weeks in culture and prepared for use. The internal wall of the controlled-release conduit was coated with 10% poly L-ornithine (Sigma) to promote the adherence of Schwann cells. The suspension of Schwann cells (3 × 105 cells/mL) was injected into the conduits. The volume of each conduit was approximately 30 μL. Therefore, the seeding amount was around 9 × 103 cells per conduit. Cells were homogeneously seeded over the inner conduits by placing on a 5-rpm rotary deck for 20 min.
Morphological evaluation of Schwann cells cultured within PCL conduits
For morphological evaluation, the Schwann cells adhering to the lumen of PCL conduit were fixed with 10% formaldehyde in PBS for 24 h. After washed with PBS, the cell-seeded conduits were dehydrated through an ascending alcohol series, commencing at a solution of 70% alcohol, then dried in a critical point drier and gold coated. All specimens were then examined by scanning electron microscope (FE-SEM, JSM 1025; JEOL, Tokyo, Japan).
In vivo
Animals and experimental design
Forty-eight male Sprague–Dawley rats, weighing 250–300 g, were divided into five groups (Table 1). Control group (n = 8) received 15-mm reversed autografts into their right sciatic nerves. Groups NCL, LCL, MCL, and HCL (n = 10) were comprised of 17-mm PCL conduits with crosslinked gelatin (10% gelatin with 0%, 0.1%, 0.5%, and 1.0% genipin, respectively) and NGF in the conduit wall. Schwann cells (9 × 103 cells/conduit) were seeded within PCL conduits before surgery.
NCL, non-crosslink; LCL, low crosslink; MCL, medium crosslink; HCL, high crosslink; PCL, polycaprolactone; NGF, nerve growth factor.
Surgical procedure
Animals were deeply anesthetized with sodium pentobarbital (30 mg/kg i.p.) throughout the surgical procedure. Surgery was performed on the right leg of the rat under aseptic conditions. 41 After an incision had been made to the skin, the sciatic nerve was exposed by making a muscle-splitting incision. An operating microscope (Leica Microsystems, Deerfield, IL) was utilized to divide the sciatic nerve near its origin, and a 15-mm nerve segment was excised using microscissors. The nerve gap was interposed with 17-mm controlled-release PCL conduits from groups NCL, LCL, MCL, and HCL or 15-mm autograft (control group) for comparison. The autograft was reversed to prevent any branching of axons through the side branches during regeneration. The proximal nerve was anchored in the conduit by 9-0 nylon microsutures. The distal end was then sutured into the other end of the conduit. Nerve stumps at both ends were sutured into the conduit to a length of approximately 1 mm. Hence, 15 mm of the nerve gap remained between two stumps. The wound was then closed in layers using 4-0 Dexon sutures. The animals were housed in temperature- (22°C) and humidity (45%)–controlled rooms with 12-h light cycles; they had access to food and water ad libitum. All procedures followed the animal care and use guidelines set by the relevant committee of our university.
Histological analysis
Eight weeks after implantation, animals were sacrificed by administering a methoxyfluorane overdose. The implanted grafts were harvested and histologically analyzed. The nerve grafts were immediately fixed in a cold-buffered 3% glutaraldehyde solution, washed in 0.1 M phosphate buffer (Fischer Scientific, Fair Lawn, NJ), and divided into three segments to ensure precise localization. The transverse sections were obtained only at two sites: the start of the distal stump and the midpoint of the resected gap. These segments were then postfixed in 1% osmium tetroxide (Polysciences, Warrington, PA), dehydrated in a graded series of ethanol solutions, and finally embedded in spurs. The embedded nerves were cut to a thickness of 5 μm, and then stained with 1% toluidine blue.
All nerve sections were observed under an optical microscope, and photographs were taken using a Nikon digital camera. The cross-sectional area of the whole nerve section was measured under 40 × and 400 × magnification. Three randomly selected fields with an area of 80 μm × 60 μm in each nerve specimen were observed at a magnification of 400 × . Analyses were conducted using an image analysis system to determine the number of myelinated axons and vessels of the mixed nerve. The numbers of component axons were extrapolated by using the area algorithm to estimate the total number associated with each nerve. The mean area of myelinated axons in each nerve specimen was also determined.
All data were obtained and expressed as mean ± standard errors (SE). The data were analyzed by analysis of variance, followed by the post hoc analytic method of Scheffe.
Results
Morphology of PCL nerve conduits
Figure 1a–d presents the morphology of a porous PCL conduit. The PCL conduit had large porosity in the inner layer and dense layer in the outer; meanwhile, no residual salt was found. The porosity of the PCL conduit was 66.2 ± 4.2%.

SEM micrographs of PCL nerve conduits; (
Release of NGF in vitro
The amount of NGF released from NCL, LCL, MCL, and HCL groups was analyzed by ELISA reader, and the test extended for 60 days (Fig. 2). The burst release occurred during the first 5 days, and slowed down thereafter. Little NGF was present in NCL groups after 30 days. Meanwhile, the steady-released NGF of HCL groups was all maintained at 0.5–2.5 ng/mL. Interestingly, the release curves of NGF in LCL and MCL were both pulse released at 5–20 days and 10–25 days. The releasing curves of NGF in these four PCL groups were all affected by the diffusion-controlled and degradation-controlled mechanisms.
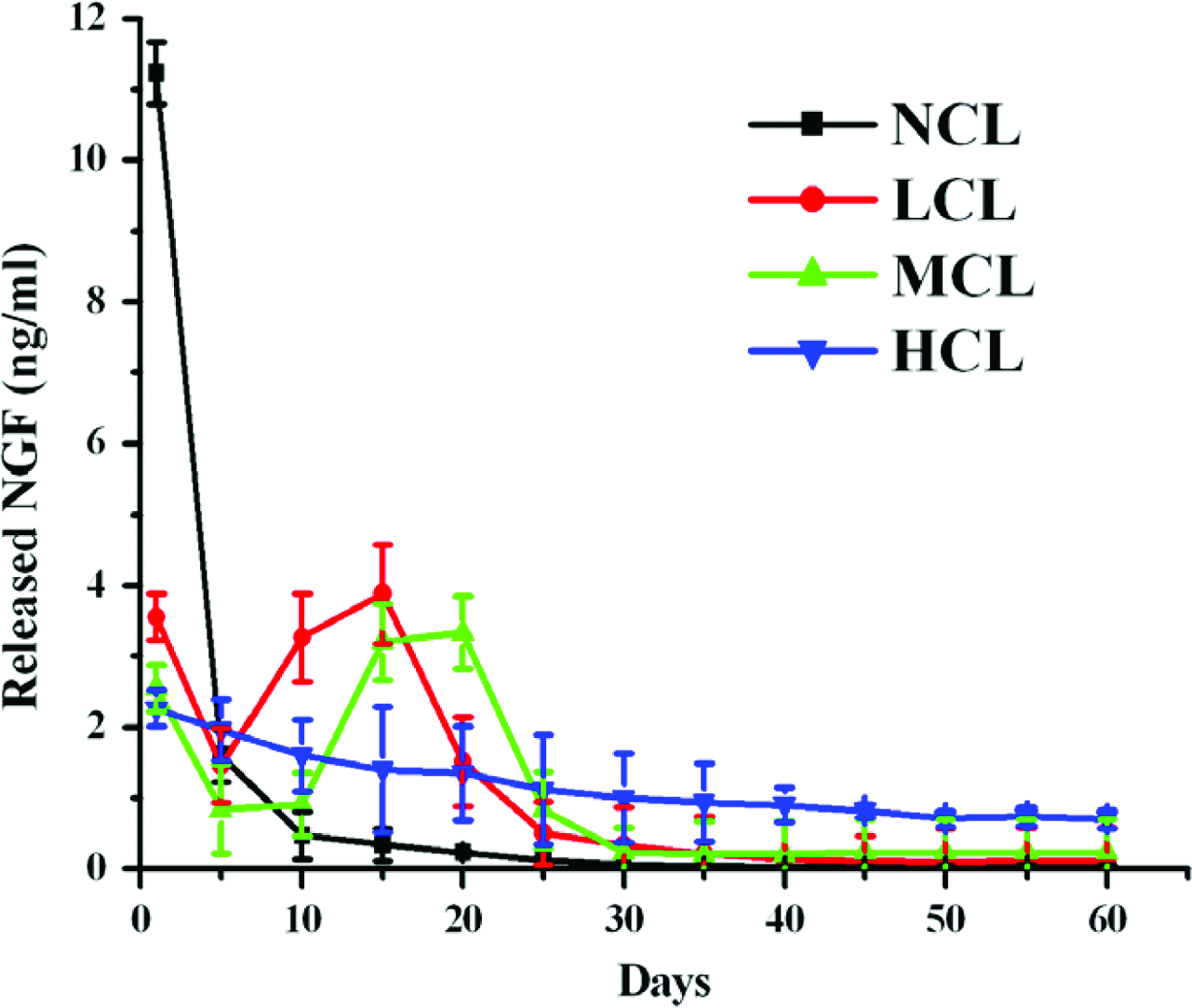
In vitro release of NGF from control-released PCL conduits, at pH 7.4 and 37°C. NCL (non-crosslink), LCL (low crosslink), MCL (medium crosslink), and HCL (high crosslink). Errors bars represent mean ± SE, n = 3 under each condition. Color images available online at www.liebertonline.com/ten.
Measurement of outgrowth of neurites from PC12 cells
The neurite outgrowth of PC12 cells was used to elucidate the bioactivity of the released NGF. Figure 3a plots the differentiation rate of PC12 cells after culturing for 1, 3, and 7 days in −NGF, +NGF, NCL, LCL, MCL, and HCL groups. About 79.7%, 82.6%, 84.5%, 76.3%, and 74.7% of PC12 cells exhibited neurite outgrowth in +NGF, NCL, LCL, MCL, and HCL groups after 7 days of culturing. Meanwhile, no statistical difference was observed between NCL, LCL, MCL, HCL, and +NGF groups. Rather, less than 3% of PC12 cells in the −NGF group exhibited fiber outgrowth in all period. These results demonstrated that the neurite outgrowth of PC12 cells differed markedly between groups with and without NGF. The bioactivity of NGF on the genipin-crosslinked gelatin is, therefore, maintained. As shown in Figure 3b, PC12 cells exhibited neurite outgrowth in NCL(−NGF), LCL(−NGF), and MCL(−NGF) groups after 7 days of culturing. In the contrary, less than 5% of PC12 cells show neurite outgrowth in PCL tube, 1.0% (w/v) genipin, and HCL(−NGF). In Figure 3c and d, the neurite outgrowth of PC12 cells show difference between the groups with and without NGF. It is demonstrated that the bioactivity of NGF was maintained within the NCL, LCL, MCL, and HCL groups at 2 and 3 weeks.
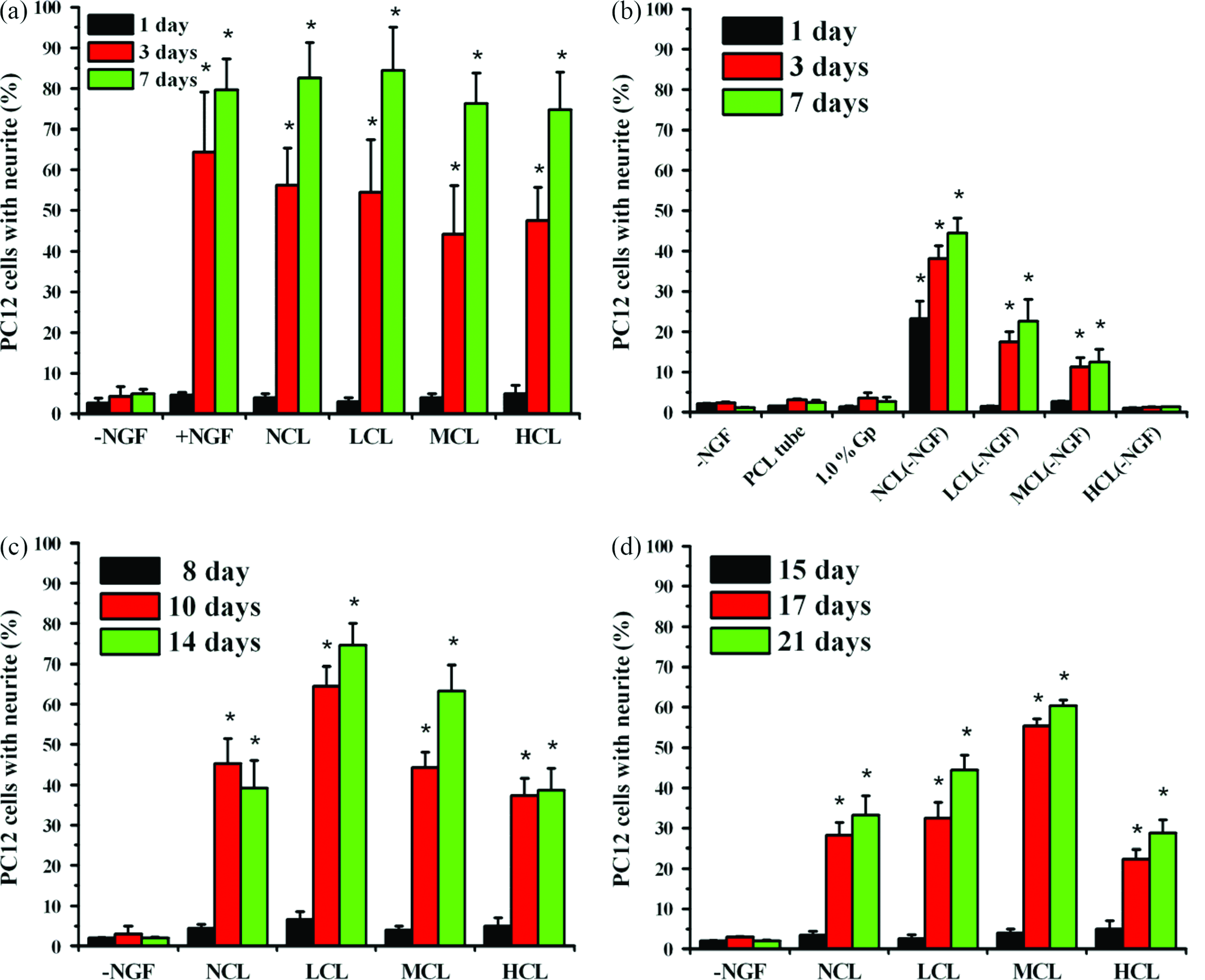
Neurite-bearing cells were counted: (
Morphological analysis
Figure 4 shows the morphology of the Schwann cells cultured in the lumen of LCL conduits. The cells adhered and extended processes well on the conduit lumen. In addition, SEM confirmed the presence of the Schwann cells on the genipin-crosslinked gelatin membranes throughout the 4 days of culture.
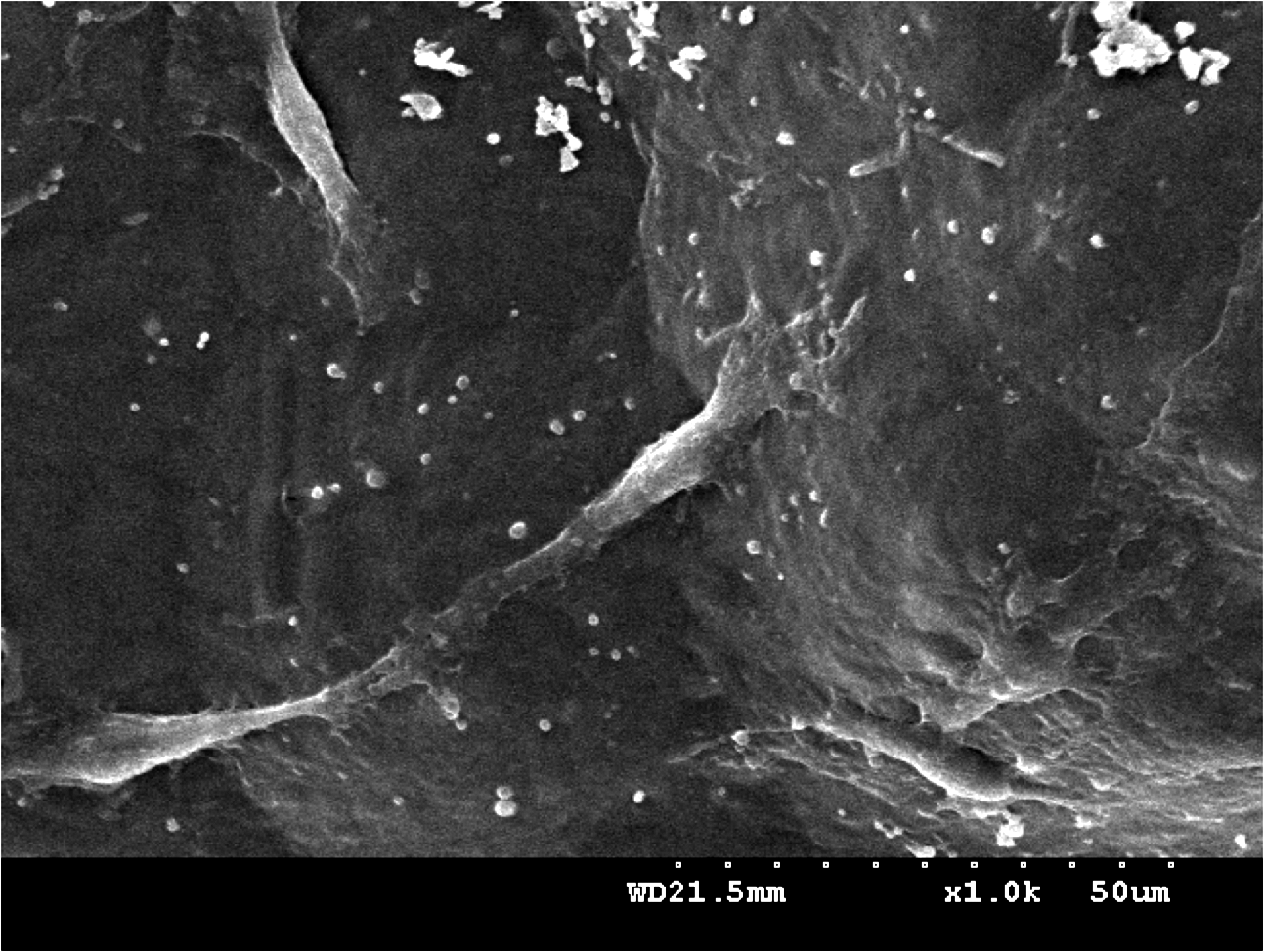
Scanning electron micrograph of Schwann cells attached and adhered to the lumen of LCL nerve conduit after 4 days in culture. The cells adhered and extended processes well on the surface.
In vivo
In the in vivo study, the regenerated axons growing across the 15-mm gap indicate successful nerve regeneration. By comparison, the unsuccessful nerve regeneration describes the case where none of axons connected the proximal and distal nerve stumps separated by 15 mm. Figure 5 illustrates the rates of successful nerve regeneration across the 15-mm gap in all groups. The NCL groups displayed low rates of successful regeneration, of 40.0% (4 of 10), compared to LCL, NCL, and HCL groups: 60.0% (6 of 10), 50% (5 of 10), and 50% (5 of 10).
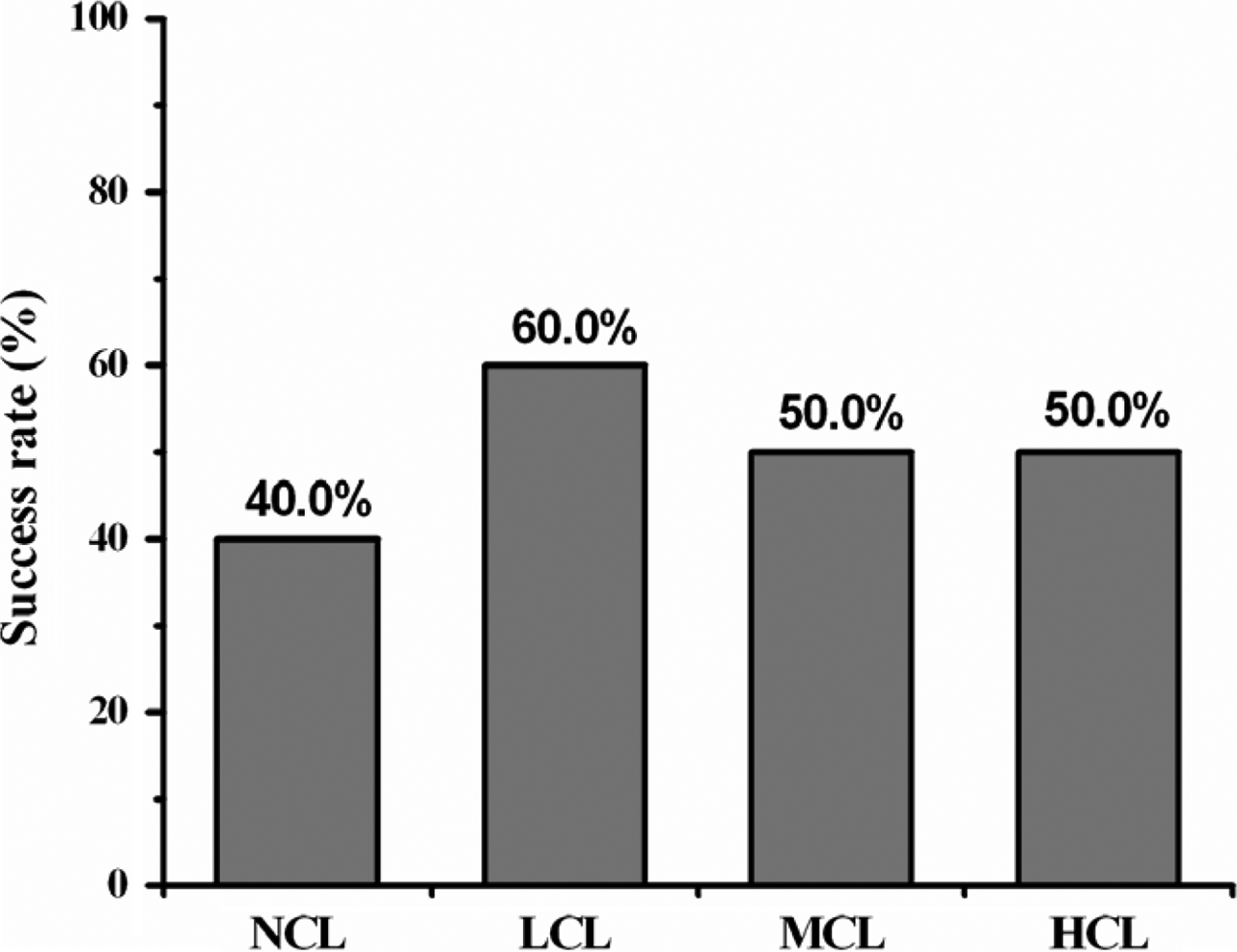
Success rates of nerve regeneration across a 15-mm gap.
After 8 weeks, animals were sacrificed to investigate the morphology of the implanted nerve specimens. Histological and statistical analyses were only conducted on the 28 successful regeneration animals in the conduits of which axons were detected. In histological examination, connective tissue and small blood vessels invaded the conduits. The surrounding tissue exhibited no obvious foreign body reactions. Some axons were myelinated with myelin sheaths, while some thinner ones remained unmyelinated (Fig. 6a–f). As shown in Figures 7 and 8, the histological data revealed the numbers and area of myelinated axons in all groups. The LCL, MCL, and HCL groups were similar to autograft treatment in the numbers and area of myelinated axons. Notably, the LCL group, although insignificant, showed a trend to have the highest myelinated axon counts of the conduit-treated groups. As for the regenerated blood vessels, the autograft, LCL, and HCL groups were the highest groups in midconduit. Further, no significant differences were found between autograft and the other conduit groups in the distal nerve (Fig. 9).
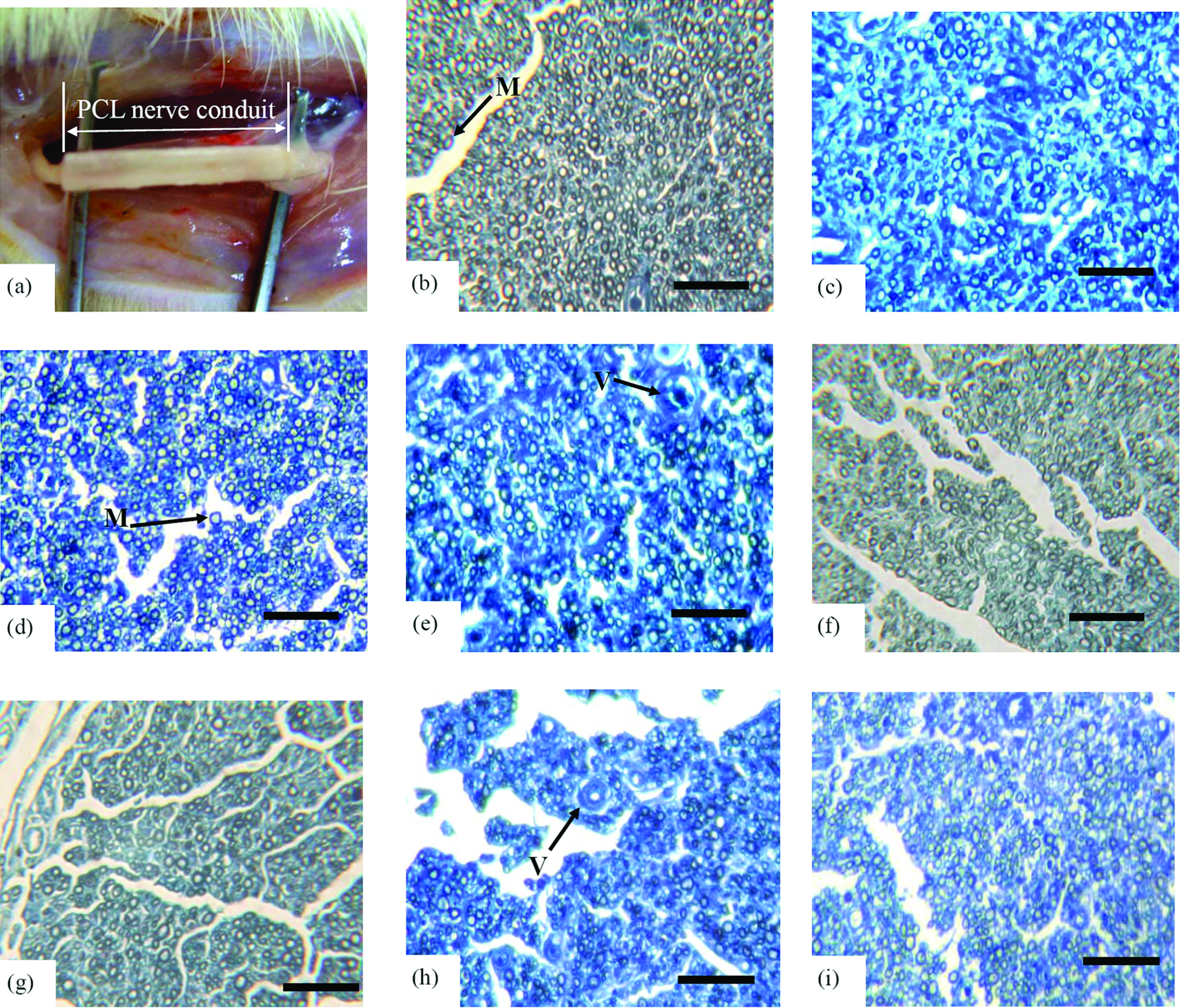
(
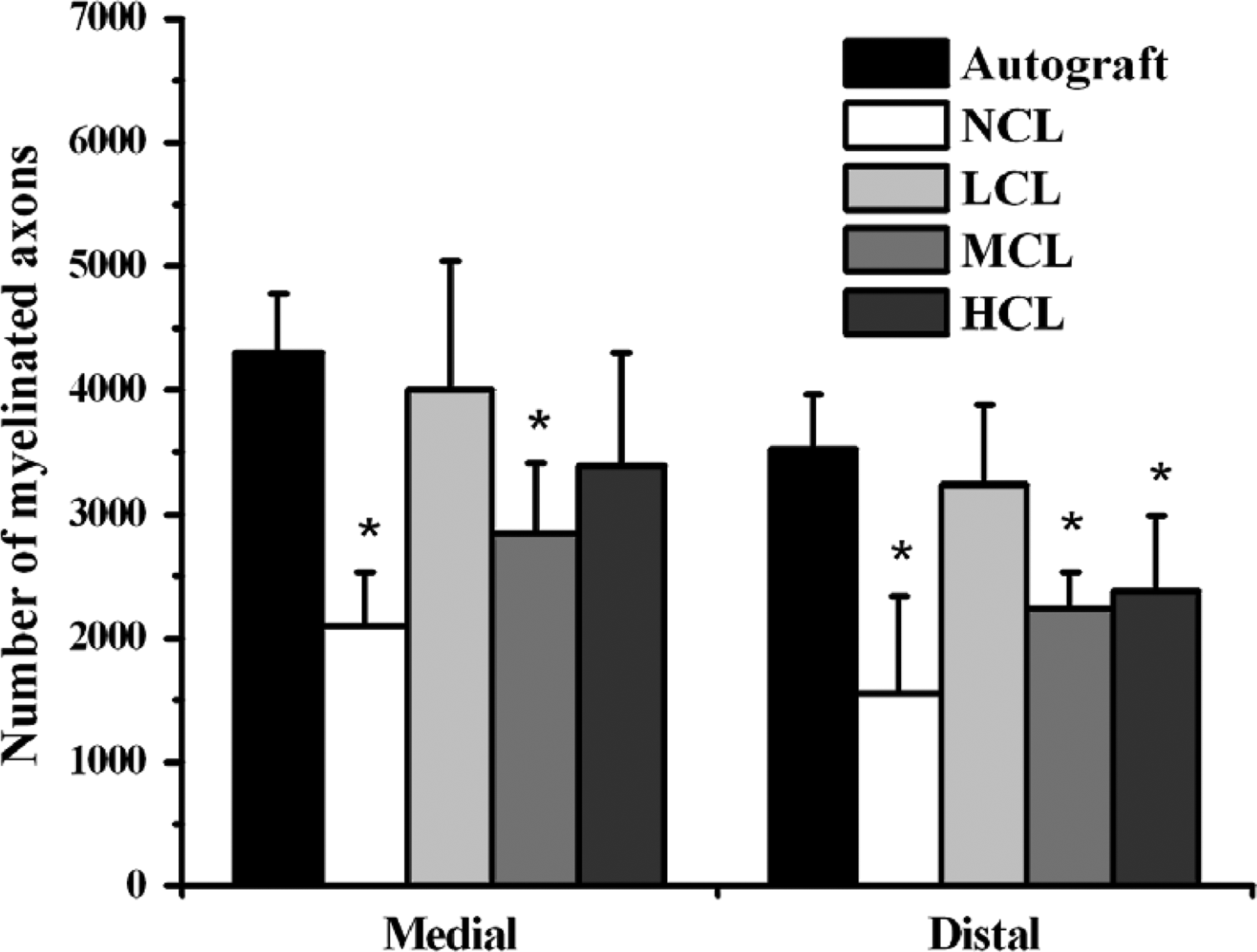
Number of myelinated axons calculated from sections at the midconduit/midgraft and distal nerve for control autograft and PCL conduit implants harvested at 8 weeks. Asterisk (*): different to control group (autograft), p < 0.05.
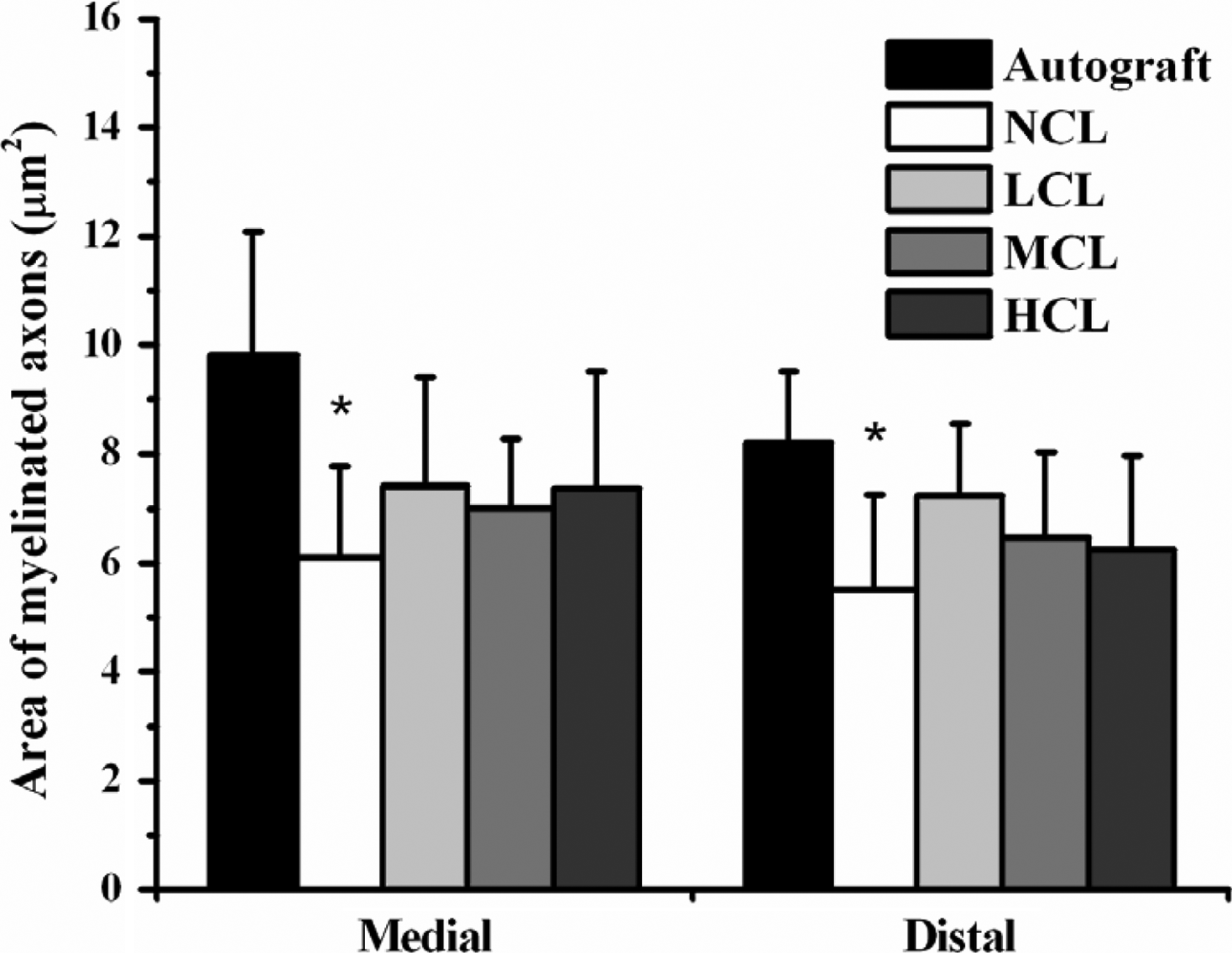
Mean area of myelinated axons (μm2) calculated from sections at the midconduit/midgraft and distal nerve for control autograft and PCL conduit implants harvested at 8 weeks. Asterisk (*): different to control group (autograft), p < 0.05.

Number of regenerated blood vessels calculated from sections at the midconduit/midgraft and distal nerve for control autograft and PCL conduit implants harvested at 8 weeks. Asterisk (*): different to control group (autograft), p < 0.05.
Discussion
The permeability is one of the factors to be considered in optimizing the design of a synthetic nerve conduit. According to our previous studies, the asymmetric nerve conduits possess the favorable effects that will enhance the outward diffusion of waste products in the early inflammatory period. 35 The morphology of the asymmetric PCL conduits was characterized by SEM. The polymers were processed into porous two-ply conduits by dipping and salt leaching. The outer layer of the conduit provides mechanical support and strength to the graft. Additionally, they have a barrier function and prevent the ingrowth of fibrous tissue that leads to scarring. 42 Meanwhile, the 37-μm pore sizes on the outer surface of the conduit also enhanced the fibroblast binding. 43 The sponge-like inner layer of the conduit allows the exchange of nutrition and fluids. In this study, sucking the added gelatin carrier into the massive space of the sponge layer further offered a new approach to prevent the inhibition of the growth of regenerated axons and vessels within conduit lumen.
NGF was first noted for its stimulatory effect on nerve fiber growth, probably via an action on transcription-dependent processes in the neuronal cell body. Additionally, the NGF high-affinity receptor (trkA) has been shown to mediate local events in guidance of dorsal nerve growth cones in vitro. 44 NGF has been demonstrated to prevent the death of axotomized sensory neurons completely following exogenous administration.45,46 Anton et al. 47 suggested that NGF and low-affinity NGF receptor promote Schwann cell migration. Schwann cells migrate more rapidly on denervated sciatic nerve than on normal sciatic nerve, and migration was inhibited with NGF antibodies. However, NGF exhibits limited stability under physiological conditions;48–50 its stability may be further reduced in protein encapsulation or chemical fixation by exposing to organic solvents, shearing, and acidic degradation of the carrier polymers. Accordingly, genipin-crosslinked gelatin was considered as a carrier in this work.
Genipin can induce neurite outgrowth via the activation of several protein kinases, including extracellular signal–regulated kinase, which is a member of the mitogen-activated protein kinase family, and activation of nitric oxide synthase (NOS) in PC12h cells. With respect to the mechanism of neuritogenesis by genipin, Dr. Yamazaki demonstrated the NO-cGMP-protein kinase G signaling pathway in PC12h cells. 51 Further, this pathway is similar to the pathway of the PC12h cells' response to NGF. Notably, PC12h cells exhibited marked neurite extension in response to an NO donor or cGMP analog, while PC12 cells did not.52,53 Dr. Chen also noted that gelatin can accelerate the differentiation of PC12 cells and extend their neurites. 54 In this research, the cells, which extended processes shorter than 40 μm, had all been expelled. Dr. van Bergeijk indicated, “cells were counted when they met the following criteria: neurites longer than one cell diameter and neurites equal or longer than 40 μm.” 55 In Figure 3, the PC12 cells in NCL, LCL, MCL, HCL, and +NGF groups all present extensive neurite outgrowth within day 21. However, no obvious neurite outgrowth was identified on PC12 cells in the −NGF group. This result indicated that the bioactivity of NGF on the genipin-crosslinked gelatin was maintained. Further, as shown in Figure 3a, NCL(−NGF), LCL(−NGF), and MCL(−NGF) groups exhibited neurite outgrowth without the stimulation from NGF. This result is in agreement with the previous finding that gelatin may promote neurite outgrowth in PC12 cells.
The cytocompatibility of materials was usually obtained by utilizing cell morphology on the different types of material surfaces.56,57 The whole process of adhesion and spreading that consists of cell attachment, filopodial growth, cytoplasmic webbing, and flattening of the cell mass was in a sequential fashion. 58 Schwann cells on genipin-crosslinked gelatin were morphologically indistinguishable from cells plated on control dishes. SEM examination also revealed that the Schwann cells flattened and extended processes in a sequential fashion as cultured within LCL conduit groups for 4 days.
The dose of added genipin was used to regulate the crosslinked degree and degradable rate, which means that we can control the rate of NGF release by changing the added genipin. As shown in Figure 2, the NCL groups show the highest initial burst release of NGF at first. The releasing is fast and ceases within 5 days and little is present after 30 days. We consider that this may refer to the quick and tremendous degradation of the uncrosslinked gelatin. On the contrast, the high-crosslinked HCL groups showed initial smallest release of NGF and maintained at 0.5–2.5 ng/mL until the 60th day. This may attribute to the bonding between NGF molecules and genipin on the gelatin carrier.59,60 The genipin molecules close to the carrier surface can strongly trap the amino groups of NGF, resulting in the decreased rate of NGF release. As for the low- and medium-crosslinked groups (LCL and MCL), the NGF dissociates from the carrier surface at the first 5 days. Then, the releasing rate is slower because the bonding between NGF and genipin was gradually disrupted by hydrolysis and enzymatic degradation. At 5 and 10 days, a considerable pulse releasing of NGF in LCL and MCL groups started, respectively. We regarded that this may be due to the increased rate of monolithic degradation in the gelatin carrier. Additionally, this is consistent with results of our previous study. 61
The optimal time course and concentration of released NGF in the bridging of a large nerve gap is critical during nerve regeneration. The bridging of short gaps (10 mm) using conduits is well known, but empty conduits have not been proven to suffice bridging gaps that are larger than 10 mm in a rat sciatic nerve.62,63 Previous studies on the axon growth across a 10-mm gap in empty silicone nerve conduits have demonstrated that axons took 2 weeks to advance from the proximal nerve stump and 4 weeks to reach the distal stump. Notably, the number of Schwann cells increased until 5 weeks after implantation of the nerve conduits, and then slowly declined. 64 Nerve injury leads to massive synthesis of NGF with a biphasic time course. In transection nerve injuries, the first increase is rapid, occurring within the first 6 h at the tips of both the proximal and distal nerve stumps. The second phase commences 2–3 days postinjury and correlates closely with infiltration of the nerve by blood-borne macrophages and maximal Schwann cell proliferation in the proximal nerve stump. Accordingly, completing 15-mm nerve gap bridging, which is significantly larger than the width of transection nerve injuries, should take delay time course (more than 3 days) of NGF to enhance the proliferation of Schwann cells at the distal nerve stump, which are considered to be the obligatory role in bridging injured nerve defects. This may explain why the LCL groups, with a pulse released of NGF at 5–20 days, have significantly more myelinated axons than NCL groups (within 5 days) in midconduit and distal conduit. Further, as comparing the different present time of pulse-released NGF between LCL (5–20 days) and MCL groups (10–25 days), we concluded that a high concentration of NGF at 5–10 days is required in bridging a 15-mm peripheral nerve injury. In terms of the concentration, the released NGF has been proven to drop rapidly in the nerve conduit, due to degradation in aqueous media, leakage from the nerve graft, and dilution by entering fluids. 65 Therefore, the LCL groups with high pulse-released concentration of NGF may maintain effective dosage as compared to the HCL groups that released low concentration of NGF. Consequently, this may clarify that the LCL groups, although insignificant, showed a trend to have more myelinated axon numbers than HCL group.
Conclusion
In this study, we demonstrated that optimal time coursed and pulse-released NGF will enhance the regeneration of myelinated axons across a large-gap nerve injury, as compared to a low dose, constant release of NGF. Further, we also found that the optimal period for NGF release in bridging a 15-mm sciatic nerve defect of rat is about 5–10 days. In addition, sucking the added gelatin carrier into the massive space of the sponge layer offered a new approach to prevent the inhibition of the growth of regenerated axons and vessels within conduit lumen.
Footnotes
Acknowledgments
The author would like to thank Miss Shih-Miao Li and Dr. Han Chang for their technical assistance and guidance in histological work. This work was financially supported by the National Science Council of the Republic of China (NSC 95-2221-E-166-003-MY2), Taiwan, and the Central Taiwan University of Science and Technology, Taichung, Taiwan.