
Abstract
Microparticles (MPs) are membrane vesicles harboring cell surface proteins and containing cytoplasmic components of the original cell. High levels of circulating MPs have been detected in pathological states associated with vascular dysfunction. We took advantage of the self-assembly method of tissue engineering to produce in vitro three vascular constructs from human vascular smooth muscle cells and fibroblasts to investigate the role of the adventitia in the modulation of vascular tone by MPs, comparing the contractile response of each of these constructs to histamine. The first two were composed of an adventitia (tissue-engineered vascular adventitia (TEVA)) or a media (tissue-engineered vascular media (TEVM)) solely, and the third one contained a media and an adventitia (tissue-engineered vascular media and adventitia (TEVMA)). In the three constructs, the results show that histamine induces contraction insensitive to blockade of inducible nitric oxide (NO) synthase (iNOS) and cyclooxygenase-2 (COX-2) and not affected by MP treatment. MPs decreased NO production and nuclear factor (NF)-κB expression but did not affect superoxide anion (O2−) release in TEVA. MPs enhanced NF-κB expression but did not affect iNOS and COX-2 expression or NO or O2− release in TEVM. In TEVMA, MPs did not enhance NF-κB expression, but COX-2 expression was higher, and O2− release was lower. Thus, MPs affected NO, O2−, NF-κB, and COX-2 in a subtle fashion to maintain the contractile response to histamine. The use of tissue-engineered vascular constructs results in a better understanding of the effect of MPs on human adventitia and media.
Introduction
It is well accepted that the media of a blood vessel is responsible for the control of vasomotor tone by contracting and relaxing in response to different hormonal factors released such as histamine and endothelin. The adventitia, on the other hand, has long been thought to serve mainly as a structural support for the media its main contribution to vascular compliance being controlled by autonomous perivascular innervation.7,8 Recent studies suggest that the adventitia influences vascular function.9–11 Nonetheless, whether the adventitia can directly participate in the regulation of vasomotor tone of blood vessels in response to MPs remains to be demonstrated.
The lack of appropriate technical procedures to separate the adventitia tunica from the other components of a native blood vessel have prevented direct investigations into the possible role of this layer in the regulation of vasomotor tone. We have developed, using the self-assembly approach, a human tissue-engineered blood vessel produced in vitro and composed of the three tunicae of native blood vessels. 12 This process consists of coaxing cells to secrete their own extracellular matrix (ECM) by adding ascorbic acid to obtain a living tissue sheet. Once formed, the living tissue sheets can be rolled or stacked to create three-dimensional organs such as blood vessels or skin substitutes.13–16 These tissue-engineered human vascular constructs possess characteristics of native blood vessels such as agonist reactivity (bradykinin, endothelin, histamine and uridine triphosphate (UTP)) 14 and structure. Thus, they allow the study of different pharmacological responses on human constructs.
In the present study, we took advantage of the self-assembly method to produce in vitro three independent vascular constructs from vascular SMCs of human umbilical artery and fibroblasts from saphenous vein. The first vascular construct was composed of only an adventitia (tissue-engineered vascular adventitia (TEVA)), a second vascular construct contained only a media (tissue-engineered vascular media (TEVM)), and the third one contained a medium and an adventitia (tissue-engineered vascular media and adventitia (TEVMA)). These three vascular models (TEVA, TEVM, and TEVMA) were reconstructed in vitro from cultured human cells to investigate the role of the adventitia in the modulation of vascular tone by MPs, comparing the response of each of these vascular constructs to histamine.
Materials and Methods
Production of MPs
The human lymphoid CEM-T cell line (CRL-2265, American Type Culture Collection, Manassas, VA) was cultured in free-serum X-vivo 15 medium (Cambrex,Verviers, Belgium). MP production was induced by treatment of 200 × 106 cells with actinomycin D (0.5 μg/mL) (Sigma-Aldrich, Oakville, Canada) for 24 h. The cell supernatant was obtained after two centrifugation steps: 750 g for 15 min and then 1500 g for 5 min to remove cells and debris, respectively. MPs from the supernatant were washed after three centrifugation steps (45 min at 14,000 g) and recovered in 400 μL sodium (NaCl) 0.9%. Determination of the amount of MPs was carried out by measuring MP-associated proteins using the Bio-Rad DC Protein Assay (Bio-Rad, Hercules, CA).
Tissue culture
Human arterial SMCs were isolated using the explant method of Ross as described before17,18 from umbilical cord obtained after informed consent of the mother. Briefly, arteries from umbilical cords were slit longitudinally, and the endothelium was removed by scrubbing with gauze. Strips of the medium were dissected, cut into small sections, and placed in a gelatin-coated, prewetted petri dish to allow their attachment to the plastic. Explants were cultured in Dulbecco's modified Eagle's medium—Ham's F12 modified medium (DMEM-Ham, ratio 3:1; Invitrogen, Burlington, ON, Canada), 20% fetal bovine serum (FBS; HyClone, Logan, UT’), and antibiotics (100 U/mL penicillin and 25μg/mL gentamicin). After 2 weeks of culture, SMCs migrated from the explants and proliferated. Two weeks later, the cells were trypsinized, plated at a density of 104 viable cells/cm2 in tissue culture flasks, and maintained at 37°C in a humidified atmosphere (92% air and 8% carbon dioxide (CO2)). Human fibroblasts were obtained from the perivascular connective tissue of saphenous vein as previously described by Laflamme et al. 9 Briefly, fragments of perivascular connective tissue were cut into smaller pieces, placed in a gelatin-coated, prewetted petri dish, and cultured, as with the SMCs. The time necessary for vascular fibroblasts (VFs) to migrate out of the explants was 9 days.
Production of tissue-engineered vascular constructs
TEVMA, TEVM, and TEVA were produced using a tissue-engineering method previously described.9,12 Briefly, human artery SMCs or VFs were cultured in DMEM-Ham's (3:1) supplemented with 30% FBS, 20 μg/mL endothelial cell growth supplement (Calbiochem, EMD Biosciences, LaJolla, CA), antibiotics, and 50 μg/mL of sodium ascorbate (Sigma-Aldrich) to stimulate ECM synthesis. After 10 to 15 days of culture, cells formed thick living tissue sheets made up of cells embedded in the ECM they secreted, which could be peeled off from the culture flask using fine forceps. TEVM and TEVA constructs were obtained by wrapping a tissue sheet of arterial SMCs or VFs around a tubular support (inside diameter 4.5 mm), resulting in four-layer-thick constructs. For the TEVMA constructs, two layers of arterial SMC sheet and two layers of fibroblasts sheet were wrapped over the tubular support to obtain a construct with the same number of layers (4) as the TEVM and TEVA constructs. These different constructs were maturated for 21 days in DMEM-Ham (3:1) supplemented with 10% FBS (FetalClone II, HyClone, Logan, UT), antibiotics, and 50 μg/mL of sodium ascorbate.
Treatment of constructs with MPs
After 21 days of maturation, each construct was cut in two while being kept on the tubular support, and each half was incubated with medium (DMEM-Ham's supplemented with 10% FBS and antibiotics) or the same medium plus MPs at a concentration of 10 μg/mL. After 24 h of incubation at 37°C in a humidified atmosphere (8% CO2), each half was cut into 4-mm-long rings (4.5 mm in diameter). The concentration and the time used to treat the three constructs corresponded to the maximal effectiveness of this type of MP in inducing endothelial dysfunction and vascular hyporeactivity in cultured cells in vitro and in aortas and small mesenteric arteries ex vivo.5,6,19
Contraction experiments
Rings of TEVM, TEVA, and TEVMA were removed from their respective tubular support used for the culture and rinsed in physiological salt solution (Kreb's solution): 119 mM NaCl, 4.7 mM potassium chloride, 1.2 mM monopotassium phosphate, 25 mM sodium bicarbonate, 1.2 mM magnesium sulfatte, 2.5 mM calcium chloride, and 10 mM glucose. Rings were separately mounted in isolated organ baths containing Kreb's solution maintained at 37°C and gassed with a mixture of 95% oxygen (O2), 5% CO2 (pH 7.4). They were set up between two L-shaped wires for isometric force measurements (Radnoti, Harvard Apparatus, Ville St-Laurent, Quebec, Canada). After being mounted, each ring was equilibrated for 30 min before being passively stretched with a preload of 500 mg. During the next 60 min, each tissue was rinsed three times, and tension was readjusted until a stable tension of 500 mg was observed. Tissues were challenged with increasing concentrations of histamine, from 10−8 to 10−4 M in absence or presence of the iNOS inhibitor, compound 1400W (100 μM), or the COX-2 inhibitor NS398 (1 μM) (Calbiochem, Meudon, France) alone or in combination.
NO spin-trapping and electron paramagnetic resonance studies
NO content was assayed after formation of electron paramagnetic resonance (EPR)-detectable Fe(II)NO (diethyldithiocarbamate)2 (Fe(II)NO(DETC)2), in rings of TEVM, TEVA, and TEVMA. Samples were placed in 24-well clusters filled with 250 μL of Kreb's solution and then treated with 250 μL of colloid ferrous-diethyldithiocarbamate (0.5 mM) and incubated at 37°C for 1 h. After incubation, the samples were rapidly frozen and kept in liquid nitrogen until EPR measurements were made. These studies were performed using a table-top x-band spectrometer Miniscope (Magnettech, Berlin, Germany). Recording were made at 77°K using a Dewar flask. Instrument settings were 10 mW of microwave power, 1 mT of amplitude modulation, 100 kHz of modulation frequency, 60 s of sweep time, and five scans. After EPR measurements, the rings were dried and weighted. The relative Fe(II)-NO(DETC)2 concentrations were determined by dividing the third component amplitude of the three-line EPR signal by the weight of the dried sample.
Superoxide anion spin-trapping
Rings of TEVM, TEVA, and TEVMA were allowed to equilibrate in deferoxamine-chelated Kreb's-4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid solution (pH 7.4) containing 1-hydroxy-3-methoxycarbonyl 2,2,5,5-tetramethylpyrrolidin (500 μM) (Noxygen, Elzach, Germany), deferoxamine (25 μM), and DETC (5 μM) (Sigma-Aldrich) under constant temperature (37°C) for 60 min. The reaction was stopped by putting samples on ice. They were frozen in liquid N2 and analyzed in a Dewar flask using EPR.
Western blotting
Western blot was performed as previously described.6,20 Protein extracts were prepared by homogenizing rings of TEVM, TEVA, and TEVMA in a lysis buffer containing (50 mM Tris, 250 mM NaCl, 8 mM magnesium chloride, 1 mM phenylmethanesulphonylfluoride, 5 mM ethylenediaminetetraacetic acid, 0.5 mM ethyleneglycoltetraacetic acid, 2 mM sodium orthovanadate, 10 μg/mL each of aprotinin, leupeptin, and pepstatine, and 1% Triton X-100). Samples were centrifuged at 14,000 × g for 10 min, and the supernatants were transferred to microcentrifuge tubes and maintained in an ice bath for immediate use. Protein concentration was determined using the Bradford method. Approximately 70 μg of total protein from supernatant fractions was separated on 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes to be probed with antibodies against iNOS, eNOS, neuronal NO synthase (nNOS), and COX-2 (BD Transduction Laboratories, Montluçon, France), NF-κB p65 (Abcam, Cambridge, UK), and β-actin (Sigma-Aldrich).
Histology and immunodetection
TEVA, TEVM, and TEVMA were formalin-fixed and embedded in paraffin. Four-micrometer sections were stained with Masson's trichrome solution. Indirect immunofluorescence detection was performed on frozen sections after fixation for 10 min in methanol at −20°C using a mouse monoclonal antibody against calponin (Sigma-Aldrich) and Alexa Fluor 594-labeled-goat anti-mouse immunoglobulin (Ig)G (Molecular Probes, Eugene, OR).
Data analysis
Data are presented as means ± standard errors of the mean, and n represents the number of rings or samples. Statistical analyses were performed using a Student t-tests or two-way analysis of variance for repeated measures with a subsequent Bonferroni post hoc test. P < 0.05 was considered to be statistically significant.
Results
Human vascular constructs
We took advantage of the self-assembly approach to produce three vascular constructs: TEVM (produced from SMCs), TEVA (produced from VFs), and TEVMA (produced from SMCs and VFs). These three constructs were produced from human vascular cells and present a tubular shape. Histological analysis with Masson's trichrome staining shows that VFs or SMCs produced and organized a dense ECM in which they were embedded. The matrix was slightly denser in the TEVA (Fig. 1B) than in the TEVM (Fig. 1A). This difference was also present in the TEVMA (Fig. 1C).

Masson's Trichrome staining (
To further characterize the constructs, calponin, a SMC marker, was analyzed using indirect immunofluorescence labeling. Strong labeling was present in the TEVM (Fig. 1D) and a portion of the TEVMA (Fig. 1F), whereas the other moieties of TEVMA and TEVA (Fig. 1E) were not labeled. These results confirms that VFs and SMCs present specific characteristics.
Contraction experiments
Histamine induced a concentration-dependent contraction in TEVA, TEVM, and TEVMA not treated with MPs (controls). Contractile response of TEVA (Fig. 2A) was much weaker (by about 10 fold) than that obtained in TEVM (Fig. 2B) or TEVMA (Fig. 2C). MPs did not significantly affect response to histamine in the three constructs (Fig. 2).
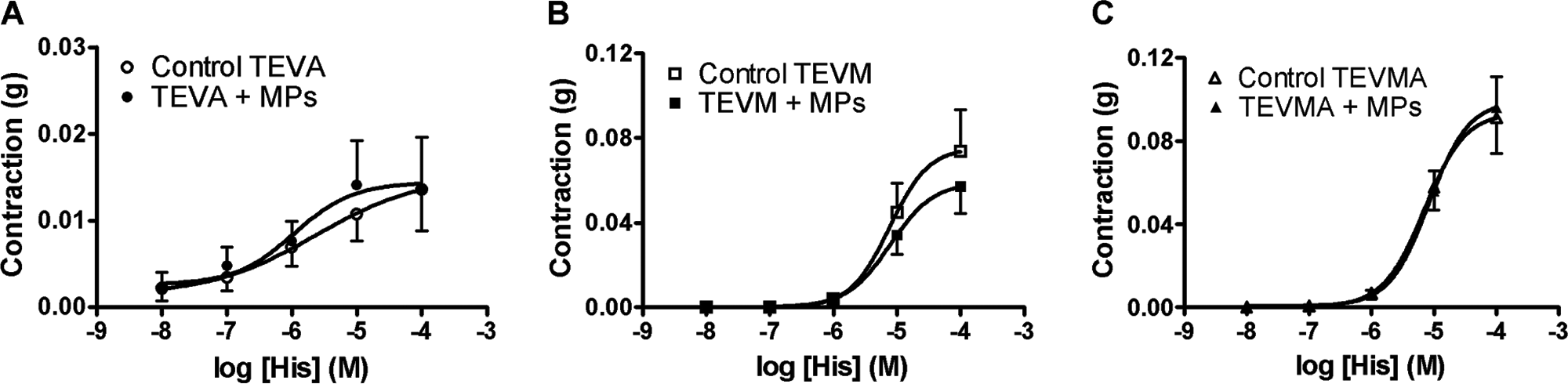
Concentration-response curve to histamine of the vascular constructs tissue-engineered vascular adventitia (
The involvement of iNOS and COX-2 was further investigated using the selective inhibitors of these enzymes: 1400W and NS398, respectively. In control TEVA, NS398 alone (Fig. 3A), 1400W alone (Fig. 3B), or the two in combination (Fig. 3C) did not significantly modify the contraction induced by histamine. In TEVA treated with MPs, NS398 (Fig. 3D) non-significantly shifted the histamine concentration-response curve to the right. 1400W (Fig. 3E) enhanced contraction to histamine, but the difference was not statistically significant. Neither inhibitor (Fig. 3F) affected the contraction to histamine in MP-treated TEVA, although the response appeared to be greater than in the control.
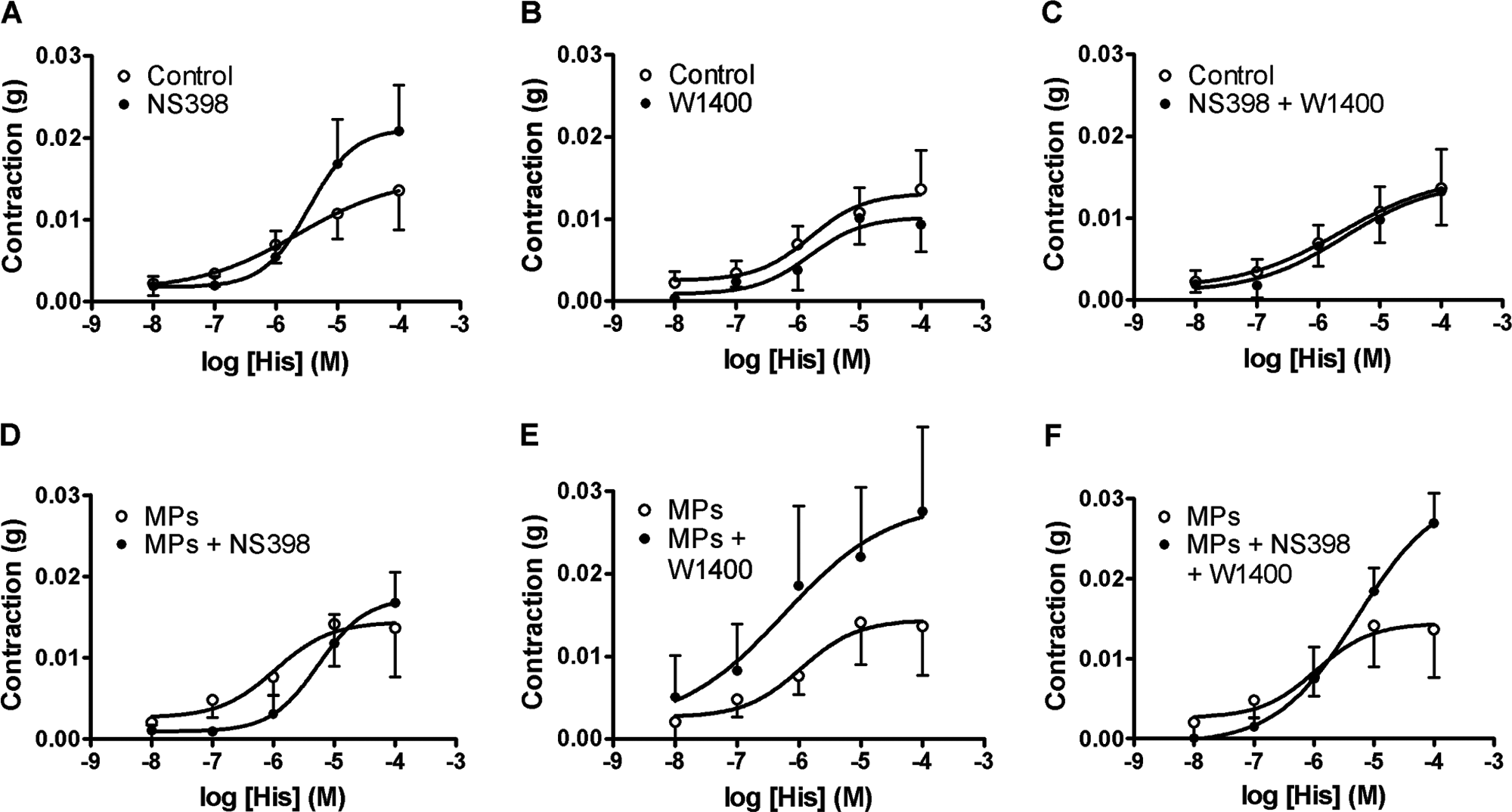
Concentration-response curves to histamine of tissue-engineered vascular adventitia incubated for 24 h in the absence (
In control TEVM, the two inhibitors, 1400W and NS398, used alone or in combination failed to modify the response to histamine (Fig. 4A-C). In TEVM treated with MPs, NS398 (Fig. 4C) or 1400W (Fig. 4D) alone produced effects identical to those obtained in untreated TEVM. Concomitant treatment with the two inhibitors (Fig. 4F) induced a non-significant increase of the contraction to histamine.
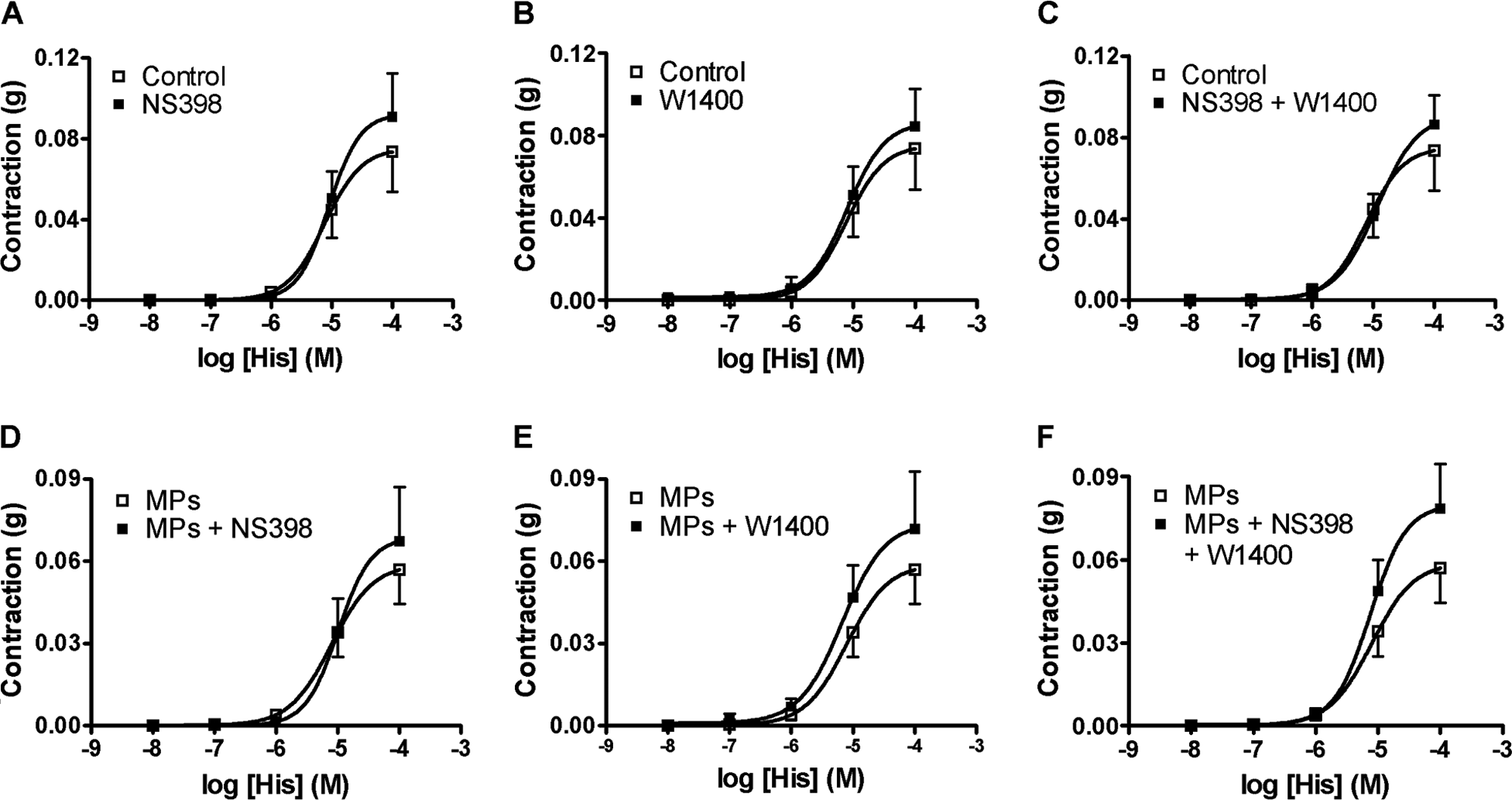
Concentration-response curves to histamine of tissue-engineered vascular media (TEVM) incubated for 24 h in the absence (
In control TEVMA and MP-treated TEVMA, neither NS398 alone (Fig. 5A, D), 1400W alone (Fig. 5B, E), nor the two in combination (Fig. 5C, F) affected the response to histamine.
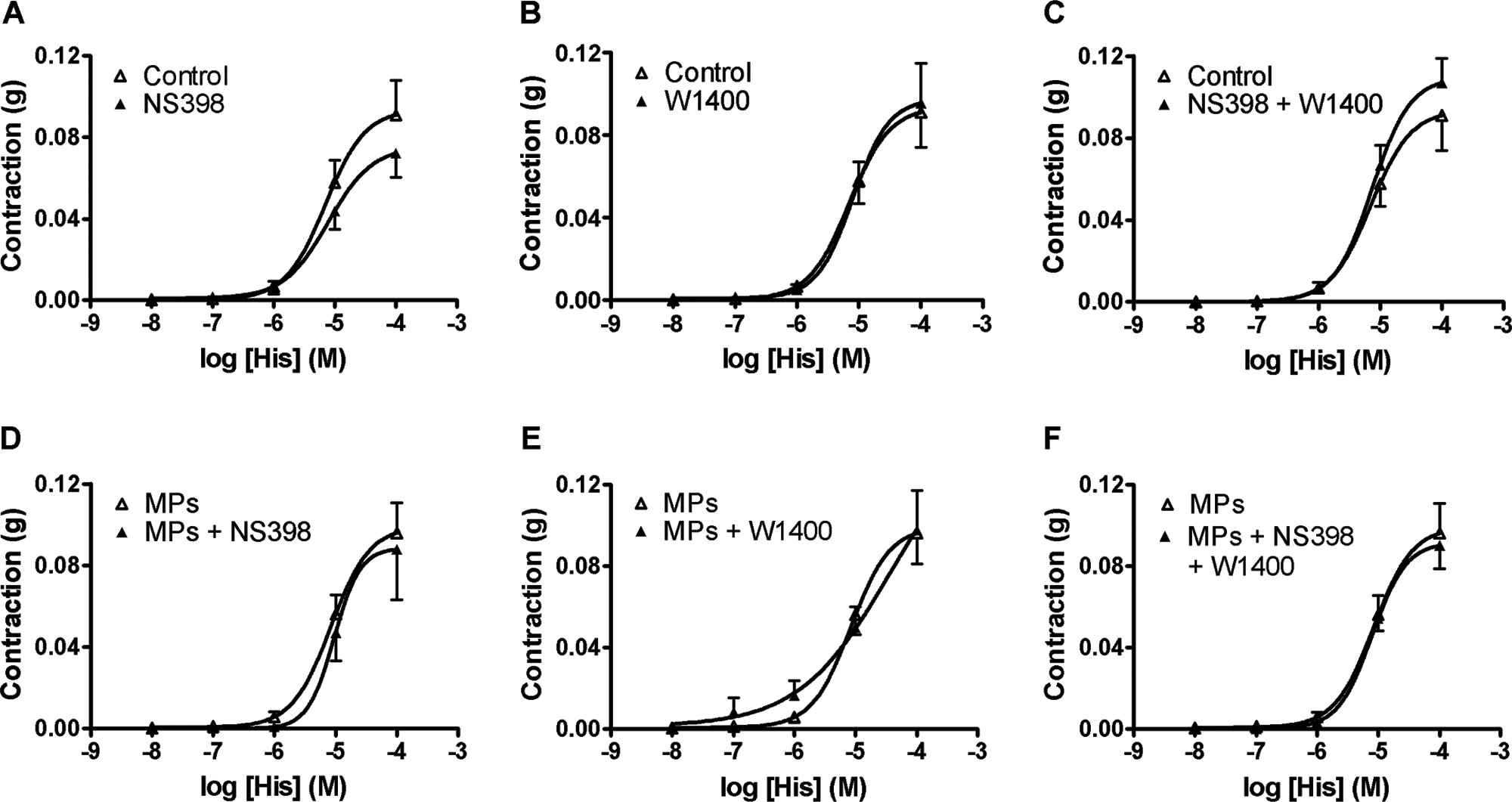
Concentration-response curve to histamine of tissue-engineered vascular media and adventitia (TEVMA) incubated for 24 h in the absence (
NO and O2− spin-trapping and EPR
As illustrated in Figure 6A, MPs significantly decreased NO content in TEVA but not in TEVM or TEVMA. With regard to O2−, MPs did not affect its release in TEVA and in TEVM, but they significantly reduced these species in TEVMA (Fig. 6B).
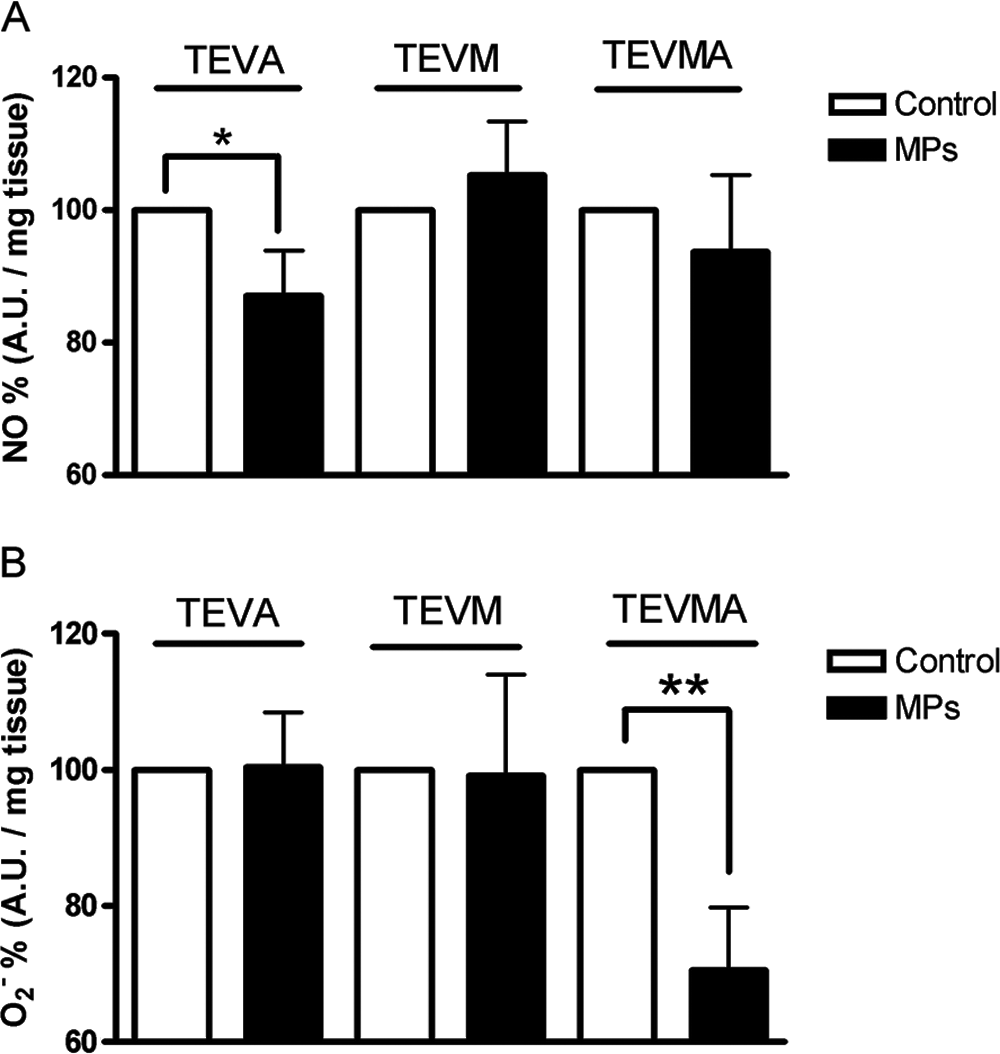
Production of nitric oxide (
Western blot analysis
Western blot of NF-κB (Fig. 7A, D) showed that MPs differentially affected its expression in the three constructs. MPs significantly reduced NF-κB expression in TEVA, whereas they markedly enhanced the expression of the same protein in TEVM. MPs failed to alter NF-κB in TEVMA. These data highlight the interaction between the adventitia and the media with respect to NF-κB. MPs did not modify COX-2 expression in TEVA or TEVM (Fig. 7B), but they significantly increased its expression in TEVMA. MPs did not modify the expression of iNOS in the three constructs (Fig. 7C). eNOS was expressed at low levels in all constructs, and MP treatment did not influence its expression. In contrast, nNOS was present in all three constructs. After MP treatment, nNOS expression was not significantly modified in TEVA or in TEVM. nNOS expression was 0.99 ± 0.37 and 1.17 ± 0.44 in TEVA (n = 4) and 1.09 ± 0.44 and 1.45 ± 0.46 (n = 5) in TEVM in control and after MP treatment, respectively, expressed in arbitrary units by taking into account β-actin level.

Western blot analysis of protein expression after microparticle treatment. Whole-protein extract was separated using 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis. Specific antibodies raised against nuclear factor-κB (
Discussion
The role of each tunicae of blood vessel is difficult to assess on natural blood vessels because it is almost impossible to mechanically separate a functional adventitia on which vasocontractile studies can be performed. Thus, we took advantage of tissue engineering that allow the production of vascular constructs made of only one tunica (TEVM or TEVA) or two tunicae (TEVMA). The aim of the present study was to investigate the role of the adventitia in the modulation of vascular tone by MPs, comparing the response of TEVA, TEVM, and TEVMA with that of histamine. The main results were that histamine was able to produce contraction in the three constructs that pharmacological blockade of iNOS and COX-2 did not affect. MP treatment did not modify the response to histamine in the three constructs. MPs decreased NF-κB expression and NO production but did not affect O2− release in TEVA. In contrast, MPs enhanced NF-κB expression, although they did not affect iNOS or COX-2 expression or NO or O2− release in TEVM. Finally, the ability of MPs to enhance NF-κB expression was blunted in TEVMA, although COX-2 expression was increased. MPs reduced O2− release in TEVMA. Thus, MPs affected the release of NO, O2−, NF-κB, and COX-2 expression in subtle fashion to keep the contractile response to histamine unchanged in TEVA, TEVM, and TEVMA.
Laflamme et al. previously characterized TEVM, TEVA, and TEVMA constructs. 9 In addition to histology confirmation of the origin of the explants, differences were observed in the time needed for the cells to migrate out of the explants and in the expression of α-smooth muscle-actin. We extended this characterization in the present study with the detection of calponin in the SMCs of the TEVM but not in the VFs of the TEVA. This confirms the validity of these constructs as appropriate models of the media and the adventitia, respectively.
Laflamme et al. 9 reported for the first time that a tissue-engineered vascular adventitia reconstructed with VFs derived from human adventitia has the capacity to contract and relax. Moreover, the adventitia was able to modulate endothelin-induced vasoconstriction. Thus, the adventitia of a blood vessel could play a greater role than expected in the modulation of blood vessel tone. The involvement of the adventitia in regulation of vasomotor tone process may require the development of strategies allowing for the administration of potentially active compounds targeting not only the media, but also the outer layer of the vessel wall. Moreover, our tissue-engineered vascular adventitia could also serve as a new model to study the vascular function of the adventitia in various pathologies. Indeed, Kleschyov et al. 21 have shown that the adventitia is a powerful source of NO triggered by lipopolysaccharide in the rat aorta. This novel source of NO has an important effect on vascular smooth muscle function and might be implicated in various inflammatory diseases. In the present study, we extended previous findings by showing that TEVA also responded to histamine, although the contraction was weaker than that obtained in TEVM. The nature of the receptor involved for this response was not determined, but it probably belongs to H1 receptor subtypes found in TEVM. 14 In contrast to the response to endothelin, TEVA did not alter the response to histamine, as indicated by the fact that the sensitivity and maximal contractile response to histamine were not significantly different in TEVM and in TEVMA.
With regard to the effects of MPs, we recently reported the effect of T-lymphocyte MPs on mice aorta without endothelium and in cultured human aortic SMCs. This type of MP promotes vascular hyporeactivity by inducing the production of vasodilatory mediators such as NO and PGI2. This effect resulted in upregulation of iNOS and COX-2 through NF-κB-dependent transcription via Fas/FasL pathway. 6 In contrast to this previous work, we found that MPs did not affect the response to histamine in the three constructs (TEVA, TEVM, and TEVMA), although they affect differentially the pathway linked to NF-κB activation associated with iNOS and COX-2 expression and the resultant changes in nitrosative and oxidative stresses. With regard to TEVM, MPs were able to increase NF-κB expression, but its increase did not result in enhanced nitrosative or oxidative stresses.
Altogether, the effects of T-lymphocyte MPs are different depending on the vessel used (i.e., mice aorta and human TEVM). These differences might be due to species or to the phenotype of vessels. Further studies will be necessary to distinguish between these possibilities. Nevertheless, our study shows that MPs can also regulate the activation of NF-κB in human TEVM.
One of the important findings of the present study was that MPs can act on TEVA in an opposite fashion than on TEVM; MPs reduced NF-κB expression and NO production in TEVA. The reason and the mechanism by which MPs produce such an effect were not directly tested. MP treatment did not affect iNOS expression. eNOS was expressed at low levels in all constructs, and MP treatment did not influence its expression. In contrast, nNOS was present in all three constructs. MP treatment did not significantly modify nNOS expression in TEVA and TEVM. Thus, the lower level of NO found after MP treatment was not associated with lower expression of iNOS or nNOS, although it cannot be excluded that MPs reduced the activity of nNOS, the regulation of which implied not only the calcium signaling, but also its association with regulatory proteins in the signaling complex: changes in intracellular location of the enzyme and phosphorylation at the activator and inhibitory residues. 22 Such an effect of the same type of MPs has been reported on cultured endothelial cells with regard to eNOS. 19 We found that MPs reduced eNOS activity but not its expression via an increase of caveolin-1 expression and enhanced eNOS phosphorylation on activator and inhibitor sites, but the eNOS phosphorylation in the latter was higher than in the former, resulting in less NO produced. Such an effect might occur in TEVA.
Nevertheless, the results underscore TEVA as one of the targets of MPs in which they can modulate vascular function. The data also highlight an interaction between TEVA and TEVM inasmuch as the ability of MPs to enhance NF-κB expression was blunted in TEVMA. In addition, MPs reduced O2− release in TEVMA and did not affect production of the same radical in TEVA and TEVM. Finally, COX-2 expression was increased in TEVMA but not in the two other constructs. Recently, we found that MPs from preeclamptic, but not from healthy, pregnant women induced ex vivo vascular hyporeactivity to serotonin in human omental arteries and mouse aortas. 23 Preeclamptic MPs induced upregulation of iNOS and COX-2 expressions, evoked NF-κB activation, and enhanced oxidative and nitrosative stresses. The MPs that most probably originated from leukocytes were responsible for the COX-2 vasoconstrictor component of preeclamptic MPs, whereas those of platelet origin were mainly involved in NO release. In the present study, using TEVMA, we strengthened the fact that T-lymphocyte MPs were able to enhance COX-2 expression, although MPs did not affect the contractile response to histamine in TEVMA despite the above changes. TEVA, TEVM, and TEVMA are reconstructed from human cells, and they allow some experimental designs that otherwise could not be achieved, for example, the cross-talk between the adventitia and the media as reported in the present study and that of Laflamme et al. 9 with regard to endothelin.
In conclusion, the results of this study obtained with a human tissue-engineered vascular adventitia reconstructed with VFs suggest that the adventitia of a blood vessel could play a greater role than expected in the modulation of vascular tone. The use of the three human vascular constructs also allows the study of the interaction between vascular cells in response to vasoactive agents. Finally, they help provide a better understanding of the effect of MPs on adventitia and media from human origin, although some differences are still present with regard to the phenotype of the media. Nevertheless, MPs regulate the interplay between vasoconstrictor and vasodilator metabolites at least from COX-2 responsible for oxidative and nitrosative stresses in human TEVA, TEVM, and TEVMA.
Footnotes
Acknowledgments
We are grateful to the members of the LOEX and Institut National de la Santé et de la Recherche Médicale (INSERM) 771; CNRS UMR 6214 laboratories, particularly to Katleen Baker, Cindy Perron, Stéphanie Pouliot, Marie-Christine Fiola, Caroline Basonie, Charles Roberge, and Abdelali Agouni for their technical support. This work was supported by the Canadian Institutes for Health Research (CIHR) and a France-Québec Exchange Program from the Fonds de la Recherche en Santé du Québec (FRSQ) and the INSERM. J.-M.B. is a recipient of a studentship from FRSQ. L.G. is the holder of a Canadian Research Chair on Stem Cells and Tissue Engineering from CIHR.