
Abstract
Abstract
Superficial-zone protein (SZP), also known as lubricin, is a key mediator of boundary lubrication and plays an important role in the functional integrity of the diarthrodial joint. The aim of this investigation was to examine the role of transforming growth factor beta (TGF-β) and interleukin-1 beta (IL-1β) on the expression of SZP in various compartments of the bovine knee joint: the superficial zone of articular cartilage, synovium, meniscus, and anterior and posterior cruciate ligaments. The effects of TGF-β1 and IL-1β on SZP expression were examined in explants and cells from the different tissue compartments. TGF-β1 up-regulated the expression of SZP in cultured explants, but IL-1β down-regulated it. Quantitative analysis of secreted proteins in the medium of the cells demonstrated significant stimulation by TGF-β1 and inhibition by IL1-β of the accumulation of SZP protein in all four tissues. Real-time polymerase chain reaction analysis revealed that TGF-β1 significantly up-regulated SZP expression and that IL-1β down-regulated it. These results revealed the modulation of SZP expression in various compartments of the knee joint by TGF-β1 and IL-1β. In addition, SZP was found to be immunolocalized at the surface layer of cells in histological sections of all four tissue compartments. Collectively, results of the current study on regulation of SZP expression by TGF-β and IL-1 help provide new insights, into tissue engineering strategies to repair and regenerate the different tissue compartments in the articular joint with optimal lubrication.
Introduction
Joint injury and arthritis result in dramatic changes in the concentration of some growth factors and cytokines such as transforming growth factor beta (TGF-β) and interleukin-1 (IL-1) in synovial fluid.19–22 IL-1 is the main pro-inflammatory cytokine that enhances catabolism in chondrocytes, 22 whereas TGF-β plays a significant role in anabolism of chondrocytes and stimulates cartilage function. 23 IL-1 and TGF-β can have significant effects on expression and secretion of SZP and lubricin by chondrocytes. IL-1 has been shown to reduce the expression and secretion of cartilage-derived SZP and lubricin, whereas TGF-β causes up-regulation of SZP and lubricin synthesis.13,24–29 However, the regulation of SZP and lubricin mRNA and protein synthesis by growth factors and cytokines in other compartments of the knee joint, such as the synovium, meniscus, and ligaments, is not fully understood. A better understanding of the modulation of SZP and lubricin synthesis in these compartments is important not only for appreciation of the biological processes governing joint homeostasis and pathophysiology, 30 but also for future functional tissue engineering approaches to fabricate functional engineered joint tissues with proper lubricating properties. The clarification of the regulatory effects of SZP expression by growth factors and cytokines may contribute to establish optimal lubrication conditions in engineered joint tissues.
The main objective in this study was to investigate the effects of TGF-β1 and IL-1β on SZP mRNA and protein expression in cultured cells from four different bovine intraarticular tissues, namely, the superficial zone of the articular cartilage, synovium, meniscus, anterior cruciate ligament (ACL), and posterior cruciate ligament (PCL) in serum-free chemically defined media using monolayer cell cultures. The second objective was to examine SZP immunolocalization in cultured tissue explants of the four distinct compartments, depending on culture conditions.
Material and Methods
Tissue acquisition for explant culture
Stifle (knee) joints from 3-month-old calves were obtained from a local abattoir and dissected under aseptic conditions. For explant cultures, full-thickness osteochondral plugs were obtained from femoral condyles using a 2-mm-diameter cork borer (Fisher Scientific, Hampton, NM). The synovia, menisci, ACLs, and PCLs were also carefully dissected. The menisci were dissected free of the ligamentous attachments, synovium, and adipose tissues. The synovial sheath and the periligamentous tissue were stripped away from the ACL and PCL. Menisci were then cut into slices (∼5 mm in thickness) along the radial-axial plane from the central aspects of the tissues using a #11 blade on a scalpel handle. Each sample was rinsed three times in Dulbecco's phosphate buffered saline (DPBS, Gibco, Grand Island, NY). For each experiment, tissues were obtained from six different animals. There were two pieces of tissue from each compartment for each animal for a total 12 samples. Tissues were cultured as explants in 24-well culture plates with Dulbecco's modified Eagle medium:nutrient mixture-F12 (Ham) 1:1 (DMEM/F-12, Gibco) containing 50 μg/mL ascorbate-2-phosphate (Sigma, St. Louis, MO), 0.1% bovine serum albumin (Sigma), and antibiotics (Medium-A) and 10% fetal bovine serum (FBS, Gibco) and incubated at 37°C in a moist atmosphere of 5% carbon dioxide and 95% air. After 24-h equilibration in the culture medium, the media were switched to serum-free fresh Medium-A with insulin transferrin selenium (ITS) + Premix (BD Bioscience, Bedford, MA) in the presence or absence of 10 ng/mL of recombinant human TGF (rhTGF)-β1 or rhIL-1β (R&D Systems, Minneapolis, MN) and incubated for 3 days.
Immunolocalization of SZP in the intraarticular tissues
For immunohistochemical localization studies, freshly isolated and explant-cultured tissue samples from the articular cartilage, synovium, meniscus, and PCL were fixed in 4% paraformaldehyde (Fisher Scientific, Fair Lawn, NJ) or Bouin's solution (Sigma), embedded in paraffin, and sectioned at 5-μm intervals. The sections were probed for SZP using monoclonal antibody (mAb) S6.79 (a generous gift from Dr. T. Schmid, Rush Medical College, Chicago, IL) with a modification of a previously described method.28,31,32 The mAb S6.79 was made in the mouse against human SZP. 32 It shows strong cross-reactivity with a variety of mammalian SZP molecules, including bovine, dog, and rabbit. Sections were deparaffinized, and endogenous peroxidase was blocked with 3% hydrogen peroxide. Thereafter, sections were blocked with normal horse serum. The sections were then incubated overnight at 4°C with a 1:5000 dilution of SZP mAb. The sections were incubated with a biotinylated horse anti-mouse immunoglobulin G (IgG) (Vector Laboratories, Burlingame, CA) for 1 h followed by 30-min incubation with VECATATIN ABC reagent (Vector Laboratories). Visualization was achieved using diamino benzine/peroxidase reaction (ImmPACT diamino benzine peroxidase substrate, Vector Laboratories), resulting in a brown precipitate. No counterstaining was performed.
Cell isolation and culture
The superficial zone of articular cartilage of femoral condyles (approximately 100 μm thick) was harvested using a dermatome. The synovium, meniscus, ACL, and PCL were manually cut into 1- to 2-mm pieces. For the isolation of cells, these tissues were digested with 0.2% (0.03% for ACL and PCL) collagenase P (Roche, Indianapolis, IN) in Medium-A containing 3% FBS for 5 h (superficial zone of articular cartilage), 1.5 h (synovium), 14 h (meniscus), and 3 h (ACL and PCL) at 37°C. The cells released from the tissues using digestion were collected using centrifugation and rinsed with DMEM/F-12, and the suspension was filtered through a cell strainer (70 μm, Falcon, BD Bioscience). Isolated cells were plated as monolayers at a density of 1 × 105 cells/well (approximately 2.5 × 104 cells/cm2) in 12-well culture plates in Medium-A with 10% FBS and incubated. Because preliminary studies had shown no significant difference in SZP expression between ACL- and PCL-derived cells, ACL- and PCL-derived cells were pooled for the present study. After 24 h equilibration in the culture medium, cells were switched to fresh Medium-A with ITS+ Premix plus graded levels (0.0, 0.1, 1.0, and 10.0 ng/mL) of rhTGF-β1 or rhIL-1β and incubated. These concentrations were based on values commonly used for other studies.25–29
Enzyme-linked immunosorbent assay analysis of SZP protein
Because most SZP is secreted into the culture medium, 1 the spent medium from the cultures of the four kinds of cells was harvested after the 3-day treatment and quantitatively analyzed in duplicate for SZP protein using a sandwich enzyme-linked immunosorbent assay, as previously described.27,28 Briefly, each well of MaxiSorp 96-well plates (Nalge Nunc International, Rochester, NY) was coated with 1 μg/mL of peanut lectin (EY Laboratories, San Mateo, CA) in 50 mM of sodium carbonate buffer, pH 9.5. To minimize non-specific binding, the wells were blocked with 1% bovine serum albumin in the same buffer. Aliquots of culture media were incubated overnight in the wells at 4°C. The wells were next incubated for 1 h with 2 μg/mL of S6.79 mAb. This step was followed by a 1-h incubation with goat anti-mouse IgG conjugated with horseradish peroxidase (Bio-Rad, Hercules, CA) and finally TMB Peroxidase EIA Substrate Kit (Bio-Rad) was used to develop color and to quantitate the absorbance. The wells were washed extensively with DPBS containing 0.05% Tween-20 (Sigma) between each step. The absorbance was measured at a wavelength of 655 nm. SZP levels were calculated using a SZP standard that was purified using affinity chromatography on a peanut lectin column (the purity verified according to immunoblot analysis) and quantified using a Micro BCA Protein Assay Kit (Pierce, Rockford, IL).
Real-time reverse transcriptase polymerase chain reaction analysis
Total RNA was extracted from the cell cultures after 24 h of stimulation using an RNeasy mini kit (Qiagen, Valencia, CA) with on-membrane DNase I (Qiagen) digestion to avoid genomic DNA contamination. Total RNA was reverse transcribed into single-strand cDNA using a high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA). Real-time polymerase chain reaction was performed in triplicate on the cDNA with an ABI PRISM 7700 Sequence Detection Systems and SYBR Green regents (both from Applied Biosystems) following the recommended protocols. SZP mRNA levels were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) levels and expressed relative to the control (untreated) culture levels (ΔΔCT methods; Applied Biosytems 33 ). The primers for bovine SZP (forward: 5’-AGAAAACCCGA TGGCTATGA-3’; reverse: 5’-TCGCCCATCAGTCTAAGGAC-3’), and GAPDH (forward; 5’-GGCGCCAAGAGGGTCAT-3’; reverse: 5’-GTGGTTCACGCCCATCACA-3’) were designed using Primer Express software (Applied Biosystems). These primers were designed to span more than 1 intron on genomic DNA and thus avoid non-specific fluorescent emission derived from contaminating genomic DNA.
Statistical analysis
All the quantitative data were presented as means ±standard deviations. A one-way analysis of variance and Fisher's protected least significant difference post hoc test were performed using StatView statistics (SAS Institute, Inc., Cary, NC) to determine the effects of TGF-β1 and IL-1β on SZP expression. A paired t-test was performed to determine the difference between the control and treated cells. P-values less than 0.05 were considered significant for all comparisons.
Results
The distribution of SZP in freshly isolated intraarticular tissues
SZP was immunolocalized in all the tissues tested. In articular cartilage, strong SZP staining was localized to the uppermost surface layer and in cells in the superficial zone (Fig. 1A). No SZP staining was seen in the middle and deep zones of cartilage. Sections of synovium showed SZP staining in the cells covering the synovial villi (Fig. 1B). In the meniscus, SZP staining was observed along the tibial and femoral surfaces (Fig. 1C). The immunoreactivity of both the surfaces was evident through these meniscal surfaces, encompassing the central (avascular) and peripheral (vascular) regions. There was no staining in deeper layers of the meniscus. In the PCL, SZP was identified on the surface. SZP staining was also observed at the interface between collagen fiber bundles within the PCL (Fig. 1D), where it was mostly concentrated, and inside the bundles in varying amounts. Sections treated with the non-specific mouse IgG showed no staining (data not shown).
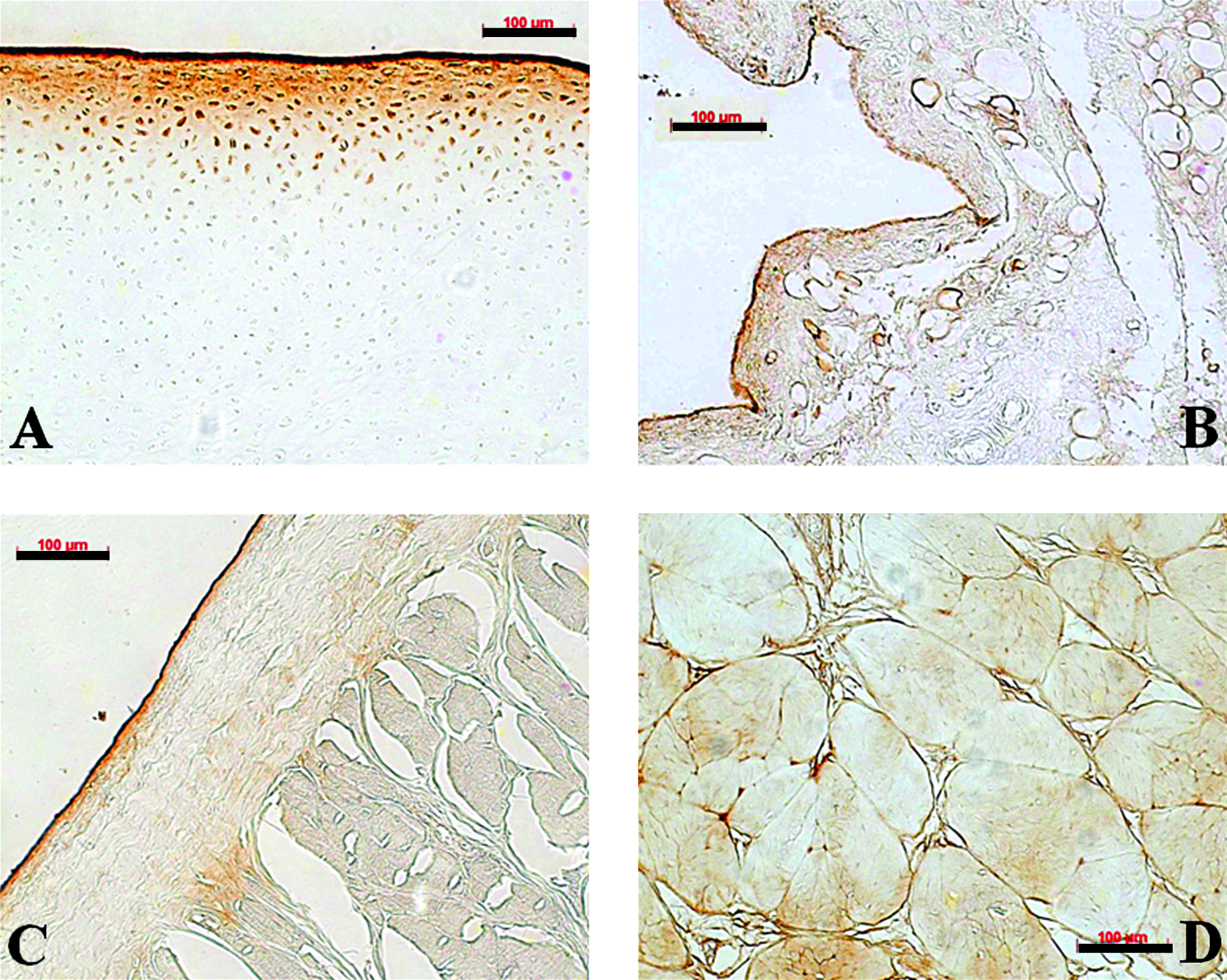
Immunolocalization of the superficial zone protein (SZP) in bovine articular cartilage (
TGF-β1 and IL-1β modulate the SZP expression in cultured explants
SZP was immunolocalized differentially in cultured tissue explants, depending on the culture conditions (Fig. 2). The distribution of SZP in the non-treated cultured explants (Fig. 2A, D, G, and J) was similar to that observed in freshly isolated tissues. The explants treated with 10.0 ng/mL of TGF-β1 for 3 days exhibited increased signal intensity of SZP at the surface (Fig. 2B, E, H, and K). In the cartilage, synovium, and PCL explants, the localization extended more deeply (Fig. 2B, E, and H). In contrast, 3-day treatment of the explants with 10.0 ng/mL of IL-1β resulted in decreased signal intensity of SZP at the surface and reduction of SZP retained within the tissues (Fig. 2C, F, I, and L).
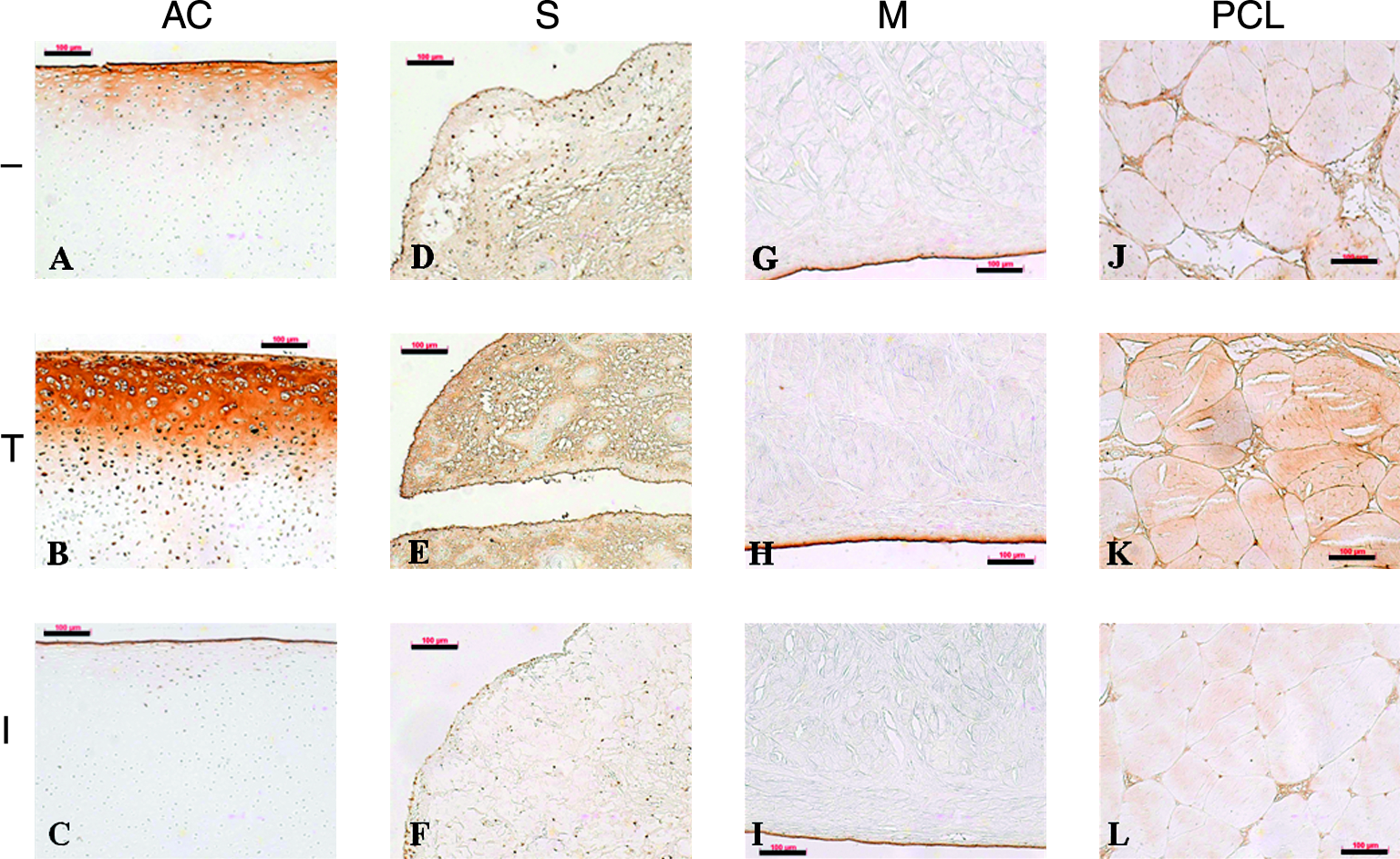
Effects of transforming growth factor beta 1 (TGF-β1) and interleukin-1 beta (IL-1β) on superficial zone protein (SZP) expression in explants of articular cartilage (AC) (
TGF-β1 and IL-1β modulate the SZP accumulation in culture medium in monolayer cells
Because the accumulation of SZP into the culture medium increased linearly in a time-dependent manner in our previous studies,26,27 the culture media were analyzed on day 3. In the control culture, the SZP protein level in the culture medium was much lower in meniscus-derived cells (∼0.5 μg/mL) and ACL- and PCL-derived cells (∼1.3 μg/mL) than in synovial cells (∼9.2 μg/mL) and superficial-zone chondrocytes (∼4.4 μg/mL) (Fig. 3). SZP accumulation in the cultured chondrocytes of the middle and deep zone was barely detected even when treated with TGF-β1 (data not shown). 27 TGF-β1 stimulated SZP accumulation in the four different cells (Fig. 3A). In these four kinds of cells, a dose-dependent (0.1 to 10.0 ng/mL) increase of SZP protein was observed with TGF-β1. The maximal effect was observed with 10.0 ng/mL of TGF-β1. There were seven times as many superficial-zone chondrocytes as in controls, eight times as many synovial cells, 10 times as many meniscus-derived cells, and eight times as many ACL- and PCL-derived cells. In contrast, treatment with 0.1, 1.0, and 10.0 ng/mL of IL-1β reduced SZP protein levels in all four cell types (Fig. 3B). Relative differences in SZP protein levels between untreated control and treatment groups were statistically significant (p < 0.05). In these cells, IL-1β at a concentration of 10.0 ng/mL exhibited the greatest inhibitory effect. With 10.0 ng/mL of IL-1β there were 48.1% fewer superficial-zone chondrocytes than in controls, 27.3% fewer synovial cells, 33.9% fewer meniscus-derived cells, and 37.7% fewer ACL- and PCL-derived cells.
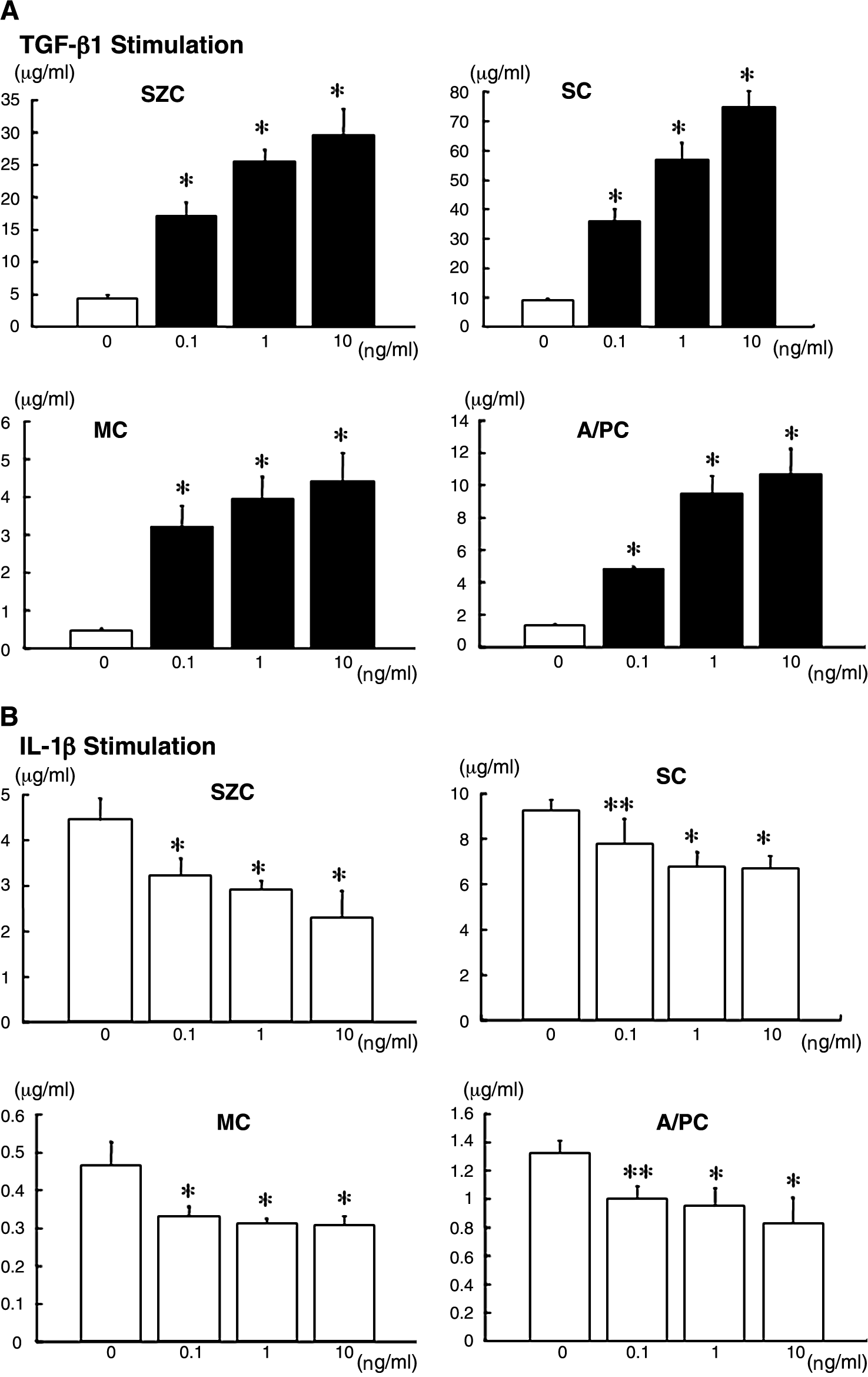
Effects of transforming growth factor beta 1 (TGF-β1) (
The effect of TGF-β1 and IL-1β on SZP mRNA expression
Treatment with TGF-β1 at 0.1, 1.0, and 10.0 ng/mL for 24 h dramatically enhanced the expression of SZP mRNA by all cell types (Fig. 4A). The degree of stimulation was approximately 22 times in meniscus-derived cells, 11 times in ACL- and PCL-derived cells, six times in synovial cells, and five times in superficial-zone chondrocytes. In contrast, 24-h stimulation of all cell types with 0.1, 1.0, and 10.0 ng/mL IL-1β significantly reduced SZP mRNA expression to 50% to 70% of the untreated controls (Fig. 4B).

Effects of transforming growth factor beta 1 (TGF-β1) (
Discussion
The present study investigated the localization and regulation of SZP expression by TGF-β1 and IL-1β in the various compartments of the knee joint: the superficial zone of articular cartilage, synovium, meniscus, and the ACL and PCL. As observed in previous studies using various animal models,2,9–13,31,34 immunohistochemical staining revealed that SZP was detected on the surface of all four kinds of freshly isolated tissues. Analyses were also performed on the cultured explants exposed for 3 days to the anabolic growth factor TGF-β1 and the catabolic cytokine IL-1β. In all four kinds of cultured explants, treatment with 10.0 ng/mL of TGF-β1 enhanced SZP expression at the surface of the tissues. In cartilage and PCL cultured explants, TGF-β1 stimulated cells deeper in the tissues to express SZP. TGF-β1 also induced cells in the sub-synovium to express SZP. On the other hand, explants treated with 10.0 ng/mL of IL-1β showed a reduction in SZP expression. Our results demonstrate how TGF-β1 and IL-1β in the synovium, meniscus, ACL, and PCL modulated SZP expression in explant culture.
We also investigated the effect of TGF-β1 and IL-1β on SZP mRNA expression and protein synthesis of cells from those tissues using a monolayer culture system. The current study is the first report to investigate the effect of TGF-β1 and IL-1β on SZP expression in cells from the meniscus, ACL, and PCL. Treatment with TGF-β1 dramatically increased SZP mRNA expression in these cultured cells. A corresponding robust increase in SZP protein accumulation by TGF-β1 was also observed. Significant differences between the control and treatment groups were observed in mRNA and protein levels. In contrast, IL-1β significantly decreased SZP mRNA expression and protein synthesis in those culture cells. These trends in regulation of SZP expression in cells from the meniscus, ACL, and PCL were similar to those from the superficial zone of articular cartilage and synovium. The quantitative data were consistent with the immunohistochemical findings in the cultured explants treated with TGF-β1 and IL-1β. That the increase and decrease in SZP secretion and accumulation were corresponding changes in SZP message levels suggests that regulation of SZP synthesis may, at least in part, be modulated transcriptionally. Taken together, our findings demonstrate that TGF-β1 stimulated SZP synthesis and expression by the intraarticular tissues and that IL-1β inhibited them.
SZP is considered to be operative and functionally important in tissues that need to slide against opposing surfaces.15,38 Cartilage slides against cartilage, as well as against other joint tissues, such as meniscus, ligament, and synovium. The presence of SZP along the surface of the meniscus suggests that it plays a role in maintaining lubrication at the articular cartilage–meniscus interface. SZP also was observed at the interface of collagen bundles within ACLs and PCLs. Because sliding between fibrils and bundles within the ACL and PCL occurs as mechanical loading is applied to ligaments, SZP may lubricate the collagen fibrils and bundles to facilitate friction-free motion of these structures within ligaments during locomotion. In addition, SZP at the interface of collagen bundles may serve another function related to ligament elasticity. 12 Patients with camptodactyly-arthropathy-coxa vara-pericarditis syndrome, in which the SZP gene is defective, lose significant tendon elasticity. 15 The presence of SZP at the surface of these tissue compartments of the knee joint could support the paradigm that SZP facilitates boundary lubrication at multiple tissue surfaces in the joints.
The total accumulation of SZP into the culture medium was higher in the culture of superficial-zone chondrocytes and synovial cells than in that of the meniscus, ACL, and PCL. However, the underlying cellular mechanisms for these differences are not clear. The rates of degradation of SZP in cells from these four tissues may be different. Another potential reason may be the use of a mixed population of cells from synovium, meniscus, ACL, and PCL for the analysis. In the current study, we isolated only superficial-zone chondrocytes from articular cartilage and cultured and analyzed them. Articular cartilage consists of three different zones (superficial, middle, and deep) evidenced by different cell morphologies, cell arrangements, biochemical compositions, and mechanical properties.35–38 Superficial-zone chondrocytes selectively synthesize and secrete SZP, whereas the middle- and deep-zone chondrocytes of the articular cartilage have limited or no capacity for SZP accumulation,1,2,27,31,38 which was compatible with the results of immunohistochemical staining in articular cartilage. In the meniscus, it has been reported that three distinct cell populations are present: the fibrochondrocytes, fibroblast-like cells, and superficial zone cells. 39 Because our immunohistochemical findings revealed that SZP was found only at the surfaces of the meniscus, it is highly probable that cells of the superficial zone selectively synthesize and secrete SZP, similar to superficial-zone chondrocytes. In the synovium, the immunohistochemical result revealed that SZP was localized only on lining layers that covered the synovial villi. We isolated synovial cells by digesting whole synovium with sub-synovium consisting of a fibrous or an adipose connective tissue. In addition, the synovium contains two types of cells: macrophage-like synoviocytes and fibroblast-like synoviocytes.
The present findings have important implications for tissue-engineering strategies that aim to replicate the lubricant function of bioconstructs formed in vitro. The articular cartilage, meniscus, ACL, and PCL are often damaged in sports injuries. Serious knee injury includes ruptures of the ACL or PCL, meniscal tears, and articular cartilage damages. These injuries can permanently affect joint function, leading to progressive posttraumatic osteoarthritis, because of the limited capacity for self-repair. Novel techniques such as mesenchymal stem cell–mediated repair and tissue engineering have shown promise and may become effective biologic therapies for such injuries in the future.40–42 However, there is little documentation of efforts to engineer joint tissues with optimal lubricating properties. The general paradigm of tissue-engineering strategies is to mimic the structure of a tissue and native environment as closely as possible to encourage the restoration of its structure and function. Gleghorn et al. suggested that controlling localization of SZP in engineered tissues may be critical for proper lubricating function. 43 Fabrication of a functional tissue-engineered joint tissue may be dependent on localizing SZP secreting cells in the surface zone or at the surface of fiber bundles. Our findings, which revealed the regulatory effects of SZP expression by TGF-β1 and IL-1β, could aid in the establishment of optimal lubrication conditions in engineered articular cartilage, menisci, and ligaments. In the current study, despite their inherent heterogeneity, synovial cells had a greater potential to synthesize and secrete SZP in cells from the intraarticular tissues tested. It has been previously demonstrated that the synovium is a promising tissue source for regenerative medicine. 44 Recently, we demonstrated that synovial cells could synthesize SZP even after chondrogenic differentiation in vitro. 45 Therefore, chondrogenically differentiated synovial cells could be used to create a superficial zone of tissue-engineered cartilage. Synovial cells could also be employed to supply SZP-producing cells at the surface of tissue-engineered meniscus and ligaments.
IL-1β is the major catabolic cytokine and is highly expressed in arthritic joints.21,22 The inhibition of SZP secretion in intraarticular tissues by IL-1β under pathological conditions may result in the loss of boundary lubricant, which under such conditions, probably leads to the further progression of osteoarthritis. In addition, this inhibition of SZP synthesis would remove many of the protective functions of SZP in intraarticular tissues. Alterations in SZP content at the surface of intraarticular tissues may lead to alterations in joint lubrication and deterioration of joint function. Conversely, the stimulation of SZP secretion in intraarticular tissues by TGF-β stimulation, as demonstrated here, may lead to functional improvement of damaged intraarticular tissues and ameliorate the pathology of joint function in arthritis, providing an efficient means to improve joint lubrication. Artificial control of SZP accumulation by TGF-β and IL-1 may lead to the cure or protection of joint dysfunction caused by arthritis.
In conclusion, we demonstrated the distribution of SZP in the articular cartilage, synovium, meniscus, ACL, and PCL and the effect of TGF-β and IL-1 on SZP expression in cells from the various tissue compartments using explant cultures and monolayer cultures. In this report, we provide novel insights into the role of TGF-β and IL-1 in the accumulation and expression of SZP in the meniscus, ACL, and PCL. Synovial cells had the highest potential to synthesize and secrete SZP in the cells from various compartments of the joint. This information will be useful in tissue-engineering strategies to repair and regenerate the different tissue compartments with optimal lubrication in the articular joints.
Footnotes
Acknowledgments
This study was funded by the Lawrence J. Ellison Endowed Chair. We thank Dr. Thomas Schmid (Department of Biochemistry, Rush Medical College, Chicago, IL) for his generous gift of the mAb S6.79 and Maggie Chiu (Department of Pathology and Laboratory Medicine, University of California at Davis, Davis, CA) for excellent technical assistance in histology.