
Abstract
Appropriate three-dimensional (3D) scaffolds and signal molecules could accelerate tissue regeneration and wound repair. In this work, we targeted human basic fibroblast growth factor (bFGF), a potent angiogenic factor, to a fibrin scaffold to improve therapeutic angiogenesis. We fused bFGF to the Kringle4 domain (K4), a fibrin-binding peptide from human plasminogen, to endow bFGF with specific fibrin-binding ability. The recombinant K4bFGF bound specifically to the fibrin scaffold so that K4bFGF was delivered in a site-specific manner, and the fibrin scaffold provided 3D support for cell migration and proliferation. Subcutaneous implantation of the fibrin scaffolds bound with K4bFGF but not with bFGF induced neovascularization. Immunohistochemical analysis showed significantly more proliferation cells in the fibrin scaffolds incorporated with K4bFGF than in those with bFGF. Moreover, the regenerative tissues were integrated well with the fibrin scaffolds, suggesting its good biocompatibility. In summary, targeted delivery of K4bFGF could potentially improve therapeutic angiogenesis.
Introduction
Fibrin, converted from blood fibrinogen via polymerization and cross-linking, plays important roles in blood clotting, cellular and matrix interactions, and wound healing. 9 Recently, the fibrin-based biomaterial has been used in tissue engineering and clinical practice because of its biocompatibility and biodegradability.10,11 The fibrin polymer is a polyporous three-dimensional (3D) structure consisting of cross-linked fibers 12 providing spatial support for cell growth and binding sites for signal molecules. Furthermore, the 3D matrix would provide the scaffold for cell growth and accelerate vascular regeneration. In this work, we constructed a functionalized system by binding bFGF specifically to the fibrin scaffold to provide signal and scaffold functions for vascular regeneration.
To endow bFGF with specific fibrin-binding ability, the Kringle4 domain (K4) was fused to the N-terminal end of the human bFGF. K4, derived from human plasminogen, an enzymogen in the blood fibrin-dissolving system, has high fibrin affinity.13–15 The recombinant protein K4bFGF was expressed and purified from the BL21 strain (D3) of Escherichia coli. The binding assay in vitro showed that K4bFGF had much higher affinity for fibrin. For function evaluation, we incorporated the K4bFGF with the fibrin scaffold and implanted it subcutaneously in rats, with the result that the fibrin scaffolds with K4bFGF markedly enhanced angiogenesis and accelerated tissue regeneration.
Materials and Methods
Synthesis of K4 and construction of the expression vector
The complete complementary DNA fragment of K4 was synthesized using polymerase chain reaction (PCR) as follows. First, the primer F1: 5′-GGC ACA TCC TCC ACC ACC ACC ACA GGA AAG AAG TGT CAG TCT TGG TCA TCT ATG ACA CCA CAC CGG CAC CAG AAG ACC CCA GAA-3′ and the primer B1: 5′- AGG GGC CTT TAT CGG CAT CTG GAT TCC TGC AGT AGT TCA TTG TCA GGC CAG CAT TTG GGT AGT T
Protein expression and purification
For the expression of the recombinant proteins, single clones of the BL21(D3) strain of E. coli containing the vectors p-K4bFGF and p-bFGF were inoculated into 10 mL lysogeny broth (LB) medium as primary cultures, followed by inoculation of 4 mL of primary cultures into 200 mL of fresh LB medium and cultured at 37°C at 200 rpm until A600 reached 0.6 to 0.8. Expression of fusion proteins was induced by addition of 1 mM isopropyl-beta-D-thiogalactopyranoside (IPTG) at 30°C for 7 h at 200 rpm. The bacterial pellets were collected using centrifugation at 8000 g for 10 min and re-suspended in 15 mL phosphate buffered saline (PBS, pH 7.3) containing 0.5 M sodium chloride (NaCl), followed by ultrasonic treatment for 5 min at 100 W. The supernatants containing soluble fusion proteins were isolated using centrifugation at 13,000 g for 30 min at 4°C. Nickel chelating Sepharose (Amersham Biosciences, Piscataway, NJ) was used to purify the fusion proteins from supernatants. The supernatants were allowed to flow through the nickel column, followed by washing non-specific proteins with 40 mL of 50 mM and 5 mL of 100 mM imidazole in PBS (pH 7.3) containing 0.5 M NaCl successively. Bound fusion proteins were eluted into 15 mL of 200 mM imidazole in PBS (pH 7.3). Finally, the imidazole in the eluted solution containing fusion proteins was eliminated using ultrafiltration (Amicon Ultra-15 Centrifugal Filter Devices, Millipore, Billerica, MA). The concentration of the protein solution was quantified using the Bradford method. The purity of final proteins was assessed using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot.
Western blot
The proteins were separated using 15% SDS-PAGE under the reducing conditions and electrophoretically transferred onto polyvinylidene fluoride Western blotting membranes (Whatman, ShangHai, China). The membrane was then incubated with anti-(histidine)6 mouse monoclonal antibody (mAB) (1:2000 diluted, Sigma, St. Louis, MO) in tris-buffered saline Tween-20 (TBS-T) after blotting with 10% nonfat milk in TBS-T at 4°C overnight. Afterward, the membrane was incubated with goat anti-mouse immunoglobulin (Ig)G (alkaline phosphatase (ALP)-conjugated, 1:15,000 diluted, Sigma) followed by addition of ALP substrate nitroblue tetrazolium–5-bromo-4-chloro-3-indolyl phosphate (NBT-BCIP) in buffer solution (0.1 M Tris-hydrochloric acid, pH 9.0, 0.15 M NaCl, 1 mM magnesium chloride). The reaction was terminated by washing with distilled water 2 min later. The photos were taken using a Cannon camera (Tokyo, Japan).
Fibrin binding assay
An elzyme-linked immunosorbent assay (ELISA) method was used to evaluate the fibrin-binding ability of K4bFGF and bFGF. The cross-linked fibrin was formed using the method described by Wu et al. 17 Briefly, 100 μL human fibrinogen solution (100 μg/mL) was coated onto the 96-well plate (Jetbiofil, Guangzhou, China) at 4°C overnight. After blocking the exposed sites with 3% bovine serum albumin in PBS, the coated fibrinogen was digested with 100 μL of thrombin (1 NIH unit/mL) solution containing 30 mM calcium chloride (CaCl2) at 37°C for 2 h. Purified K4bFGF and bFGF with gradient concentrations in PBS were added to the wells (n = 4) and incubated for 2 h at 37°C. After extensive washing, K4bFGF and bFGF bound to fibrin were detected by adding anti-histidine tag mouse mAB (1:2000 diluted, Sigma) followed by goat anti-mouse IgG (ALPconjugated, 1:10,000 diluted, Sigma). The bound ALP was assessed by addition of ALP substrate para-nitrophenylphosphate disodium (2 mg/mL), and the color development was measured at 405 nm.
Preparation of fibrin scaffolds
Human fibrinogen containing factor XIII 0.125 U/mg fibrinogen was purchased from Hualan Inc. (HeNan, China). The fibrin scaffold was made by mixing the following components to obtain the final concentrations: 18 50 mg/mL fibrinogen, 40 mM CaCl2, 20 U/mL thrombin, and 4.35 nmol/mL K4bFGF or bFGF (approximately 15 μg native bFGF per 200 μL). To obtain the shaped fibrin scaffold, 200 μL of this mixture was allowed to be polymerized in a 48-well tissue culture plate for 1 h at 37°C. Scanning electron microscopy was used to observe the inner microstructure of the fibrin scaffold after fixation in 2.5% glutaraldehyde and freeze drying.
Subcutaneous implantation
All of the animal experiments were performed with the approval of the Chinese Ministry of Public Health Guidelines for the Care and Use of Laboratory Animals. Sprague-Dawley rats weighing approximately 200 g were used. Four subcutaneous pockets were created, as described previously. 16 The fibrin scaffolds incorporated with K4bFGF, bFGF, or PBS were implanted into the pockets (n = 5). Four days after the operation, the rats were killed using an intraperitoneal injection of sodium pentobarbital overdose to observe the neo-angiogenesis induced by the implants. The fibrin scaffolds with skins and subcutaneous tissues were excised and fixed in 4% formaldehyde for further histological analysis.
Immunohistochemistry
The primary antibody, anti-rat Ki-67 (Boster, WuHan, China), was used to test the proliferation cells. Briefly, formalin-fixed paraffin-embedded tissue specimens were cut into 5-μm-thick sections and mounted on poly-L-lysine-coated slides. The sections were deparaffinized and rehydrated. For antigen retrieval, the slides were digested with 0.1% trypsin at 37°C for 30 min. The tissue sections were then incubated at 4°C overnight with appropriate primary antibodies (von Willebrand factor, 1:150; Ki-67, 1:100) after eliminating endogenous peroxidase by incubating with 3% hydrogen peroxide for 10 min. The sections were then incubated with anti-rabbit IgG/horse radish peroxidase polymer for 40 min at room temperature. After extensive washing, 3,3-diaminobenzidine tetrahydrochloride was used to confirm the presence of immunocomplexes. Finally, the sections were counterstained with Mayer's hematoxylin for 1 min.
Statistical analysis
SPSS 13.0 (SPSS Corp., Chicago, IL) was used for all statistical analyses. Multiple comparisons were performed using one-way analysis of variance (S-N-K). P-values < 0.05 were considered significant.
Results
Structures of the expression vectors and the recombinant proteins
The expression plasmid p-K4bFGF contained three inserted elements, as shown in Fig. 1A. The 6-histidine is an affinity tag for purifying the recombinant protein; K4 is a fibrin-binding module, and bFGF is the functional working region. To avoid possible interference between K4 and bFGF, a 15-residue linker was inserted between them. The p-bFGF, as a control, did not contained K4 (Fig. 1).
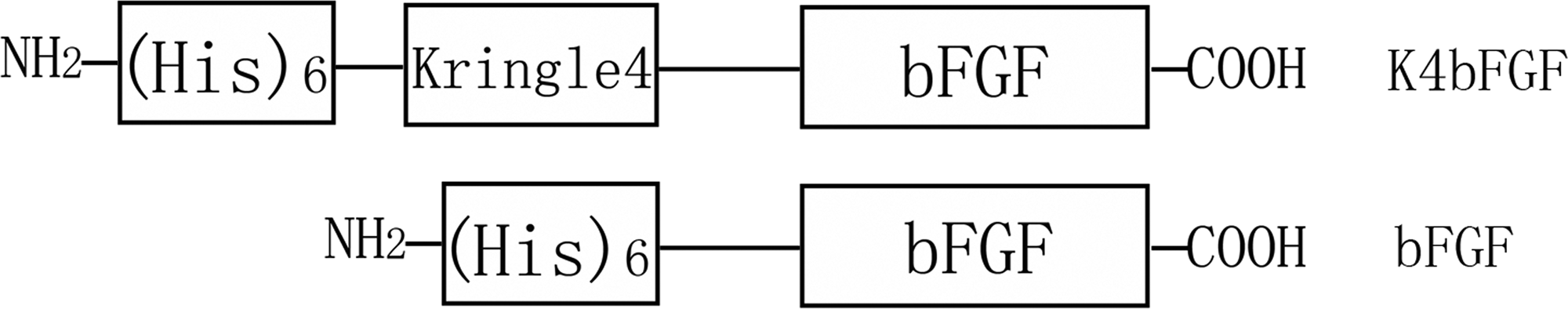
Schematic diagram of the expression plasmids of Kringle4 domain basic fibroblast growth factor (K4bFGF) and bFGF. The 6-histidine is an affinity tag used to purify the recombinant proteins; K4 is a fibrin-binding module, and bFGF is the functional region.
Production and characterization of the K4bFGF and bFGF
After induction with IPTG, E. coli BL21, containing p-K4bFGF and p-bFGF, produced an approximately 35kD (K4bFGF) and an approximately 20kD (bFGF) protein (Fig. 2A, lanes 2 and 5), which accorded approximately with the theoretical molecular weights (K4bFGF, 30.418 kD; bFGF, 19.416 kD). Their identities were further confirmed using Western blot against (His)6 tag (Fig. 2B). K4bFGF and bFGF were mainly expressed in supernatants (Fig. 2A, B, lanes 3 and 6). Both fusion proteins isolated from supernatants using one-step affinity chromatography had a high degree of purity as demonstrated by SDS-PAGE and Western blot (Fig. 2A, B, lanes 4 and 7).
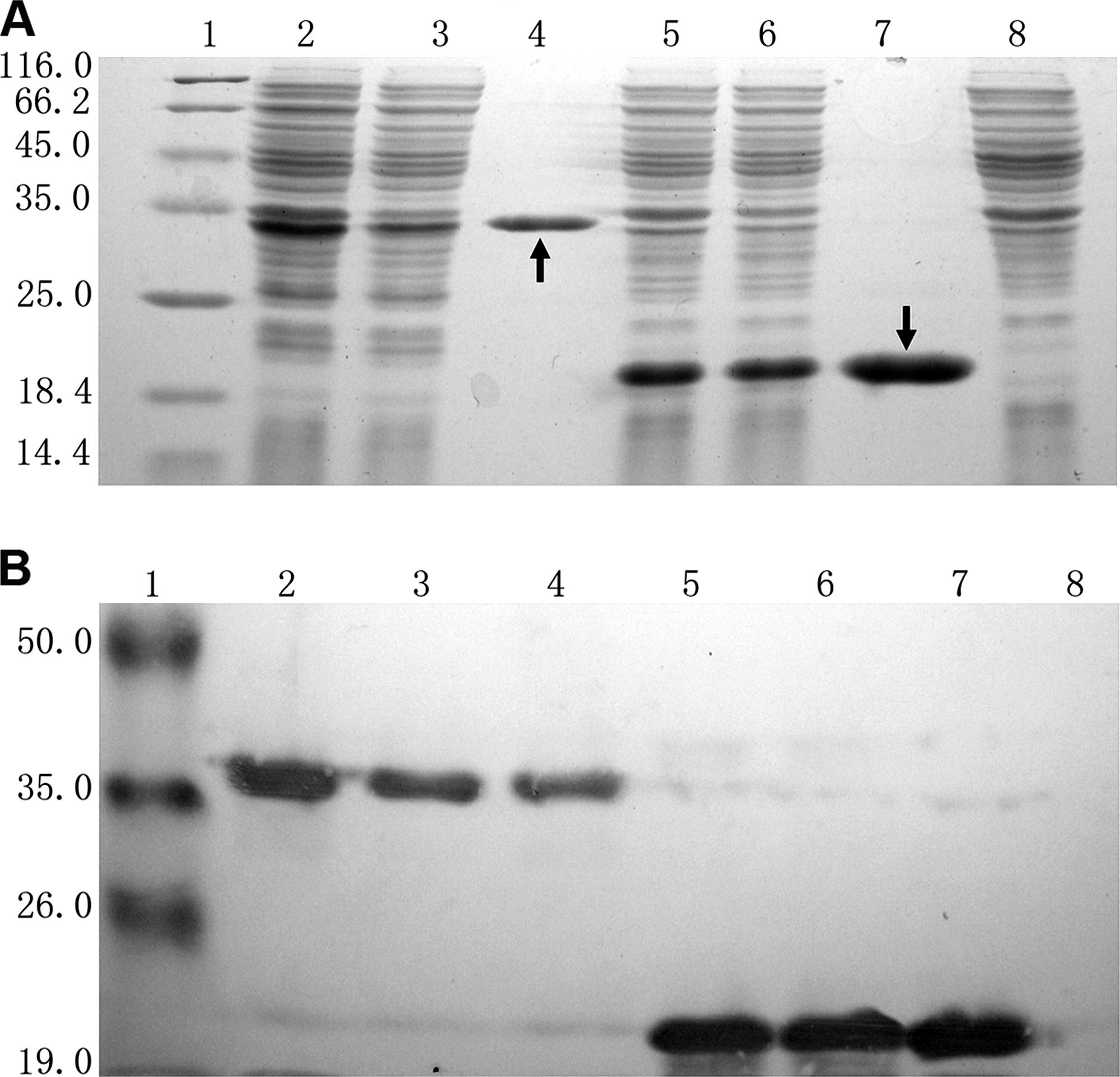
Expression, purification and identification of Kringle4 domain basic fibroblast growth factor (K4bFGF) and bFGF. (
K4bFGF showed high fibrin-binding ability
Introduction of K4 aimed to bind bFGF specifically to the fibrin. However, three pairs of disulfide bonds exist within native K4. The cystine residues have the potential to form incorrect disulfide bonds in K4bFGF and lead to the loss of fibrin-binding ability, so we performed an ELISA using the purified fibrin to determine whether K4bFGF retains fibrin-binding ability in vitro. Previous studies have reported bFGF with some fibrin affinity, 19 as demonstrated in our study (Fig. 3). At low concentrations, bFGF and K4bFGF had similar binding curves and exhibited fibrin-binding properties, whereas the amount of bFGF bound to the fibrin reached the maximum at 1 μM, and more-elevated concentrations did not lead to greater bound amounts of bFGF. In contrast, bound amounts of K4bFGF increased continuously with increasing concentrations (Fig. 3). At the 5-μM point, the optical density (OD) value of K4bFGF reached 0.560 ± 0.036, twice the OD value of bFGF (0.272 ±0.011) (p < 0.001), indicating that K4bFGF has higher fibrin-binding capacity than bFGF. Scatchard analysis further revealed K4bFGF had higher fibrin affinity (Kd = 0.470 μM) than bFGF (Kd = 5.956 μM). This assay demonstrated that fusion to K4 endowed bFGF with high fibrin-binding ability.
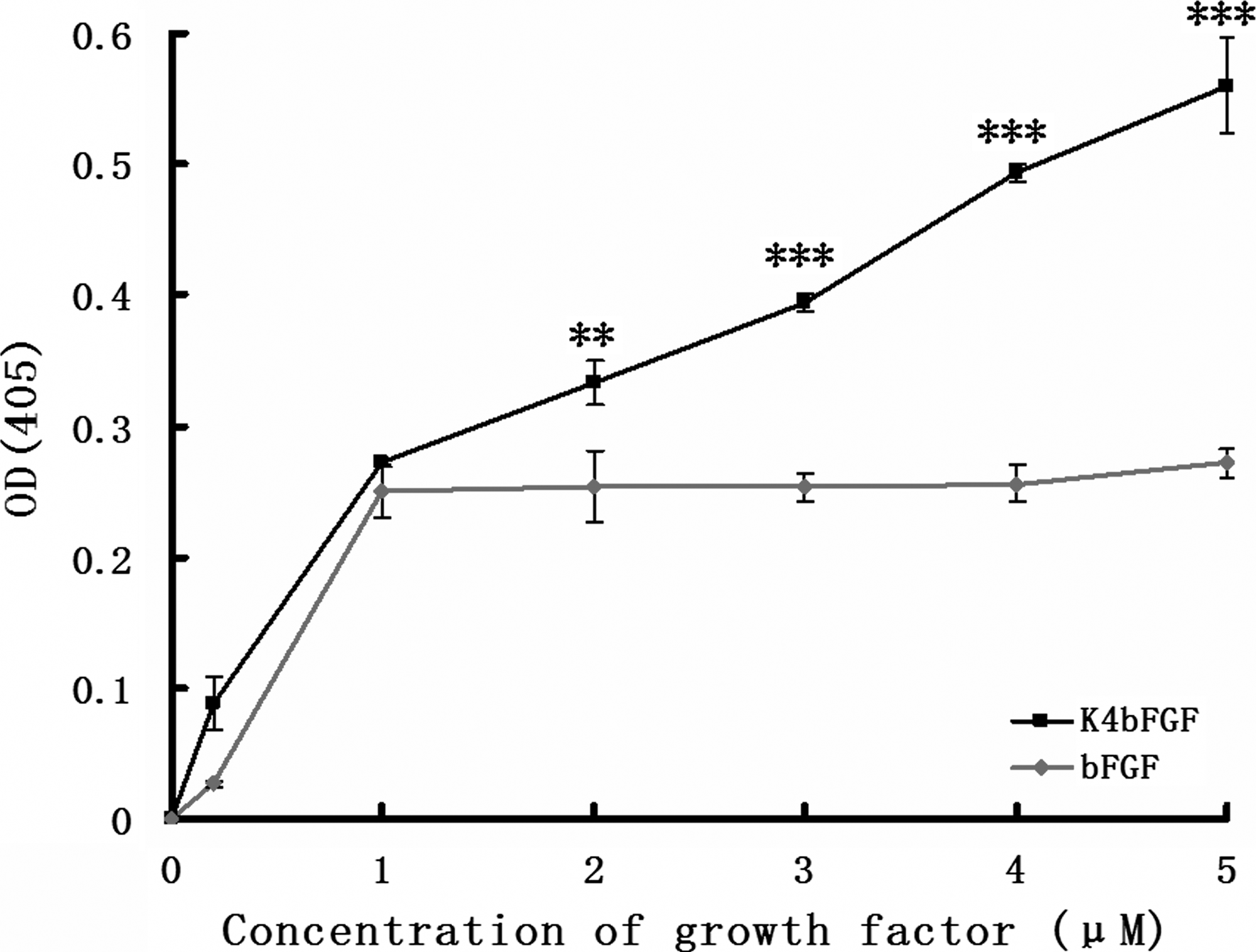
Binding curves of Kringle4 domain basic fibroblast growth factor (K4bFGF; black) and bFGF (red) for fibrin matrices. The y-axis represents optical density (OD) value, reflecting the bound amounts of K4bFGF and bFGF, and the x-axis represents the concentration of K4bFGF or bFGF applied. **p < 0.005; ***p < 0.001. Data are presented as means ± standard deviations (n = 4).
Microstructure of the fibrin scaffold
The scaffold is a shaped fibrin gel with plasticity (Fig. 4A). Fibrinogen concentration and thrombin dosage can control its mechanical strength. The interactions between the fibrin fibers and the K4bFGF molecules and the inner configuration are illustrated in Fig. 4B. The microscopic structure of the fibrin scaffold is honeycomb-like, with an average pore size of 50 μm, as revealed by scanning electron microscopy (Fig. 4C). This polyporous 3D structure was suitable for cell migration and growth. A high-power micrograph revealed that the fibrin fibers were cross-linked and distributed regularly (Fig. 4D), providing potential binding sites for signal molecules participating in tissue regeneration (Fig. 4B). This artificial fibrin scaffold made from purified human fibrinogen had a similar 3D structure to that of natural fibrin clots. 18
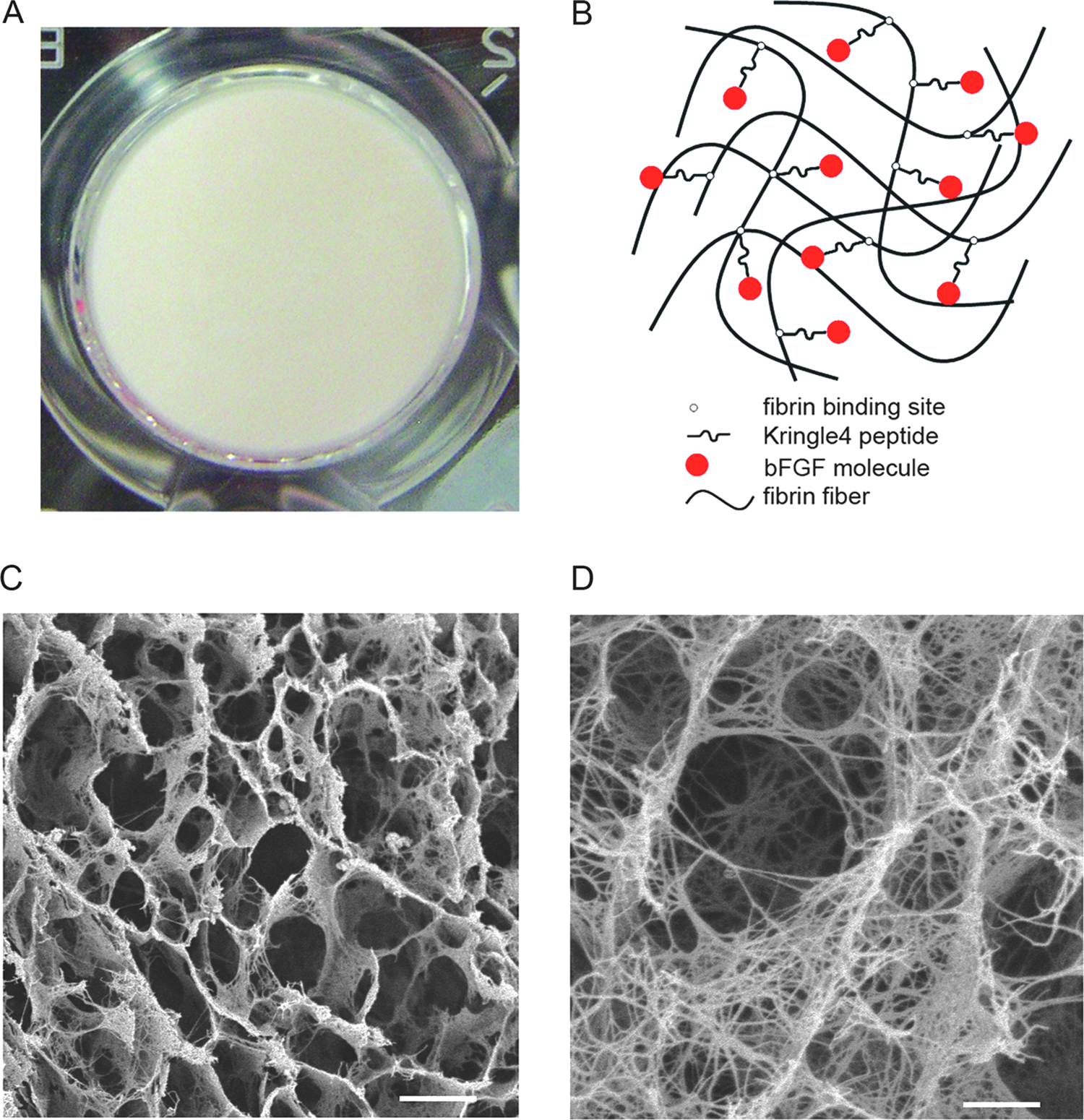
The appearance and structure of the fibrin scaffold. (
Fibrin scaffolds incorporated with K4bFGF markedly enhanced angiogenesis
In vitro assay demonstrated that K4bFGF has similar mitogenic activity to native bFGF (data not shown). When implanted subcutaneously, the fibrin scaffolds incorporated with PBS, bFGF, and K4bFGF induced different angiogenesis. On the PBS groups, only a few microvessels were observed (Fig. 5A); this effect was usually seen after surgery and was considered to be compensatory angiogenesis. 20 The scaffold with bFGF induced angiogenesis a little more than that with PBS (Fig. 5B, I). However, the scaffold with K4bFGF was much more efficient at inducing angiogenesis (Fig. 5C, I). Large numbers of newly formed blood vessels were observed, and these robust neovessels grew regularly across the fibrin scaffolds and were distributed with tree-like branches. These results suggest that K4bFGF could bind specifically to the exogenous fibrin scaffolds and enhance angiogenesis at the wound sites.

Angiogenesis stimulated by the scaffolds incorporated with phosphate buffered saline (PBS), basic fibroblast growth factor (bFGF), or Kringle4 domain bFGF (K4bFGF). (
The fibrin scaffold is a good support for cell growth and tissue regeneration. 18 Hematoxylin and eosin staining was used to observe the changes of fibrin scaffolds after subcutaneous implantation. On PBS groups, the fibrin scaffolds were almost degraded completely, and no apparent angiogenesis was observed (Fig. 5D). On bFGF groups, no marked new vessels were observed except for some cells that had invaded into the scaffold (Fig. 5E). Just like the changes on the PBS groups, an empty cavity was formed around the scaffold after its degradation, suggesting that no regenerative tissues were formed or that the new tissue formation was much slower than fibrin degradation (Fig. 5D, E). However, a large quantity of new blood vessels could be observed around the fibrin scaffolds incorporated with K4bFGF, and many of them infiltrated into the scaffolds (Fig. 5F–H). Moreover, substantial tissues were regenerated during the fibrin scaffold degradation, and these regenerative tissues invaded into the scaffold and were integrated well with it. No distinct borderlines or empty cavities were observed between the scaffold and the surrounding tissues, as demonstrated by the fibrin residual bodies in the regenerative tissues (Fig. 5F, G). Thus, only the combination of K4bFGF and the scaffold, but not the single bFGF or the fibrin scaffold, could efficiently enhance angiogenesis and tissue regeneration.
Fibrin scaffolds with K4bFGF accelerated cell proliferation
To further determine whether K4bFGF incorporated with fibrin scaffolds could accelerate cell proliferation, we performed the immunohistochemical staining of Ki-67, a nuclear antigen recognizing proliferation cells at all stages (Fig. 6). On the PBS and bFGF groups, few Ki-67–positive cells were observed in fibrin scaffolds, whereas many typical Ki-67–positive cells migrated into the fibrin scaffolds with K4bFGF (Fig. 6A-C). Also, K4bFGF bound to the fibrin scaffolds stimulated significantly more Ki-67–positive cells in the tissue surrounding fibrin scaffolds than PBS or bFGF did (p < 0.05) (Fig. 6D–F). These data further confirmed that K4bFGF incorporated with fibrin scaffolds could accelerate cell proliferation.
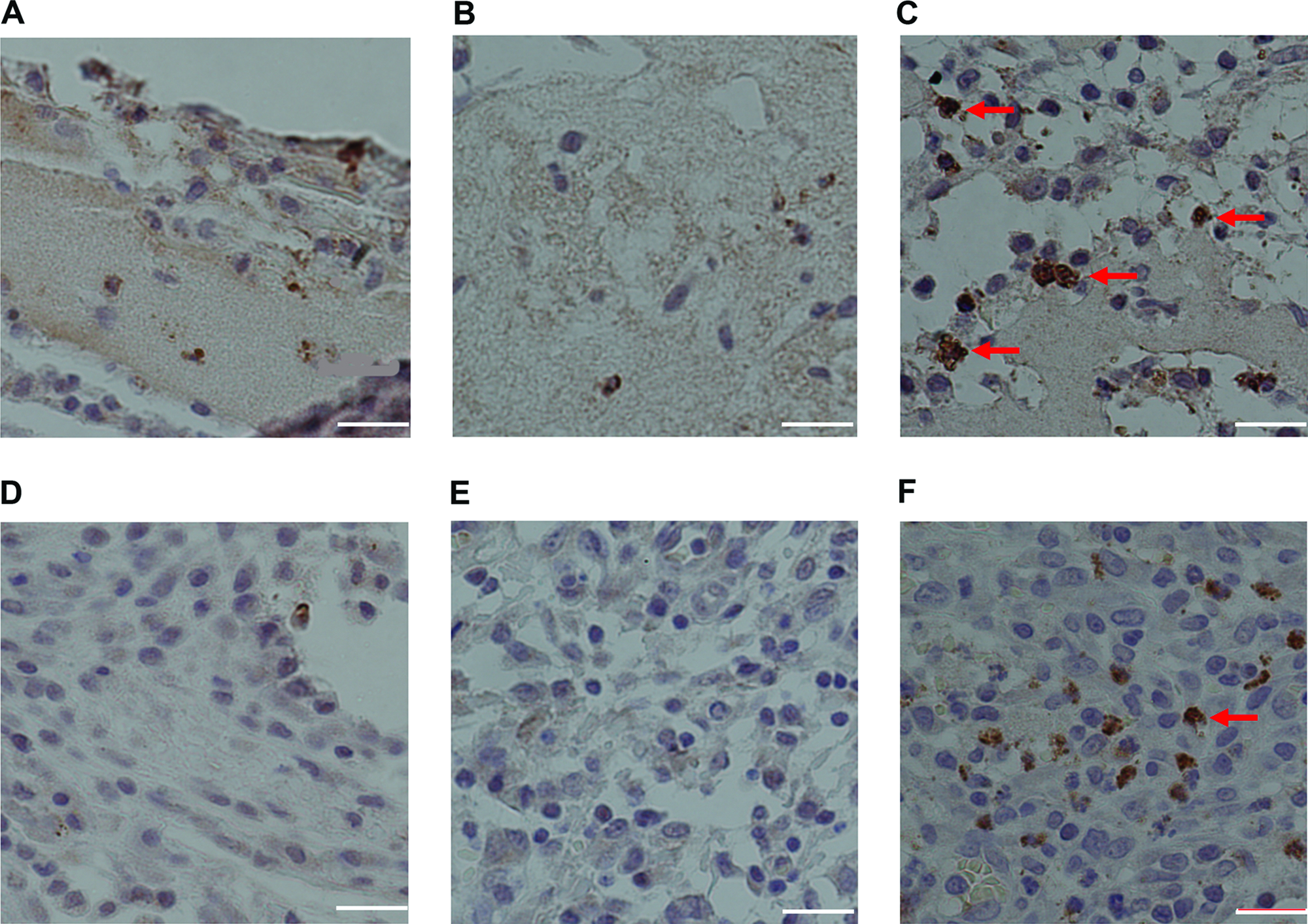
Immunohistochemical staining with Ki-67 antibody. (
Discussion
bFGF is a potent angiogenic factor and a promoter of cell proliferation. It has been used for therapeutic neo-angiogenesis and wound repair. However, traditional treatment by injection usually causes low target-specificity of bFGF at wound sites; therefore a multi-time and high-dose delivering manner is required. This leads not only to low efficacy, but also to potential adverse effects on normal tissues, because FGFs reportedly contribute to cancer angiogenesis, and over-expression of bFGF induces vascular tumors.21,22 One solution is to deliver bFGF in a site-specific manner. In this work, we targeted bFGF specifically to the fibrin scaffolds so that K4bFGF could be delivered in a site-specific manner and promote targeted angiogenesis and tissue regeneration. In addition, the artificial fibrin scaffold had a similar 3D structure to that of natural plasma clots, 23 which would provide spatial support for cell growth and tissue regeneration.
To endow bFGF with high fibrin-binding ability, we fused a fibrin-binding peptide, K4, to the N-terminal end of bFGF. The relative fibrin-binding ability of bFGF and K4bFGF was compared using an ELISA, in which the OD values and actual amounts of proteins had a linear relationship when OD values were less than 2.0 so that the OD values reflected the bound quantity. The binding curves indicated that bFGF had a binding site to fibrin that was similar to K4bFGF at concentrations lower than 1 μM. However, the additional increase for K4bFGF at higher concentrations suggested that K4bFGF had another binding site, which the binding site within the K4 might explain. Subcutaneous implantation of the fibrin scaffolds with K4bFGF induced robust angiogenesis. Cell proliferation could reflect tissue regeneration, so the Ki-67 antibody was further used to evaluate the proliferating cells. More Ki-67–positive cells in the K4bFGF group demonstrated that K4bFGF could accelerate tissue regeneration. Unlike the ones with PBS and bFGF, the fibrin scaffolds with K4bFGF were completely integrated with regenerative tissues, suggesting good biocompatibility of this system. Thus, this system holds promise for improvement of ischemic diseases and therapy for large-sarea ulcer and tissue loss, where angiogenesis is usually impaired and appropriate scaffolds are absent for cell migration and tissue regeneration.
The fibrin can be isolated from patients' own blood and will not elicit immune responses and pathogen contamination when implanted in vivo. The fibrin scaffold is biodegradable in vivo, and the degrading speed could be controlled by modulating concentrations of Ca2+, fibrinogen, and thrombin, 18 which could avoid the problems caused by too-short or too-long exposure on the bFGF. Too-short treatment with bFGF would cause insufficient angiogenesis, and too-long treatment with bFGF would be deleterious, because overexpression of angiogenic factors by gene transfer resulted in dysfunctional vessels, edema, and hemangioma formation. 24
Footnotes
Acknowledgments
This work was supported by grants from NSFC (30688002; 30600304), the Ministry of Science and Technology of China (2006CB943601), the Chinese Academy of Sciences (KSCX2-YW-R-133), and the K. C. Wong Education Foundation.