
Abstract
Despite the rapidly growing body of work on stem cell–based adipose tissue engineering, there remains much to be learned about the role of the scaffold and culture environments in directing the adipogenic differentiation of cells. The present study examined how various culture environments and differentiation stimuli (traditional differentiation medium [DM] and coculture with mature adipocytes) impacted the adipogenic differentiation of human preadipocytes, with studies progressing from two-dimensions (2D) to three-dimensions (3D) in vitro. Assays for adipogenic markers (leptin, adiponectin, and glycerol) and Oil Red O staining were used to assess differentiation. After 16 days of 2D culture, adipogenesis was substantially greater when preadipocytes were cocultured with adipocytes rather than treated with DM. In a 3D in vitro environment, the production of adipogenic markers was significantly elevated relative to 2D conditions, and the coculture condition continued to stimulate greater adipogenesis. Alterations in 3D scaffold physical properties had only a minimal effect on the function of mature adipocytes, but significantly impacted the ability of preadipocytes to undergo adipogenic differentiation in vitro. These alterations in scaffold environment and in medium conditions, particularly the application of adipocyte/preadipocyte coculture methods in lieu of traditional DM, may provide further means for optimizing adipogenic outcomes in vitro and in vivo.
Introduction
One strategy to generate adipose tissue involves the application of tissue engineering techniques. Adipose tissue is derived from embryonic mesoderm, much like mesenchymal bone marrow stem cells. 14 Preadipocytes are unipotential precursors of adipocytes that can be isolated from the stromal vascular fraction of excised or lipoaspirated fat.15,16 Preadipocytes are readily available for use as a cell source; upon differentiation, these cells develop lipid vacuoles and morphological changes characteristic of adipocytes.6,17
Several research groups have used a scaffold-based tissue regeneration approach for adipose tissue engineering with variable success. 18 Multiple carriers of either preadipocytes or mesenchymal stem cells have been used, including poly(lactide-co-glycolide acid), matrices,19–21 collagen scaffolds,22–26 hyaluronic acid (HA),27–31 Matrigel,32,33 fibrin matrices,34,35 or adipose-derived basement membrane. 36
Yet, despite this rapidly growing body of work on stem cell–based adipose tissue engineering, there remains much to be learned about the roles of the scaffold and culture environment in directing the adipogenic differentiation of cells. The large differences between scaffolds, treatments, and culture conditions across previous studies often make it difficult to directly compare these works against each other or draw conclusions about the impact of the culture environment on adipogenic differentiation. Moreover, much of the work in optimizing conditions for preadipocyte differentiation into adipocytes has been performed only in two dimensions,37–40 although cell behavior in two-dimensional (2D) environments tends to be very different from that in three-dimensional (3D) environments.41–43
In the current study, we explored the effects of varied scaffold and culture conditions on the differentiation of preadipocytes to adipocytes in a 3D environment. To accomplish this analysis, it was necessary to identify a base scaffold material that was appropriate for soft tissue applications, but also created a very defined, controllable, and characterizable environment. Thus, hydrogels of poly(ethylene glycol) (PEG) were employed as scaffold materials, and the adipogenic differentiation of preadipocytes was investigated as a function of scaffold properties, culture dimensionality, and culture medium treatments.
Materials and Methods
All reagents were obtained from Sigma-Aldrich (St. Louis, MO), unless otherwise specified.
Preadipocyte and adipocyte isolation
Adipose tissue samples were collected from adult patients undergoing elective plastic surgery operations according to human subjects protocols approved by the UW-Madison Hospital Institutional Review Board. Adipose samples of excised and lipoaspirated fat were transported under sterile conditions to the lab and digested within 30 min of harvest.
Adipose tissue was processed according to the protocol used by Clavijo-Alvarez et al. 12 Briefly, adipose tissue was washed three times in phosphate-buffered saline and then minced and digested in 1 mg/mL of type 1 collagenase (Worthington Biochemical, Lakewood, NJ) for 1 h at 37°C under constant shaking. The digest was then filtered through a 100 μm nylon mesh and centrifuged at 1000 rpm for 10 min, which separated the mature adipocytes (upper layer) from the stromal vascular fraction (pellet). The pellet was resuspended in erythrocyte lysis buffer (10 mM potassium bicarbonate, 1 mM ethylenediaminetetraacetic acid, and 154 mM ammonium chloride), and then centrifuged again at 1000 rpm for 10 min, with the resulting preadipocyte-rich pellet suspended and cultured in basal medium (phenol red-free Dulbecco's modified Eagle's medium:Ham's F12 basal medium containing 10% fetal bovine serum, 2 mg/mL L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin). Mature adipocytes were collected after the first centrifugation and cultured in either an inverted T-flask (adapted from Tholpady et al. 44 ), on a glass slide with silicone spacers, or on a transwell (TW) insert, depending on the requirements of the specific assay. Fresh medium was added to all cells 18–24 h after isolation and then every 2–3 days thereafter. The viability of freshly isolated preadipocytes and adipocytes was measured via Trypan blue staining.
Generation of differentiation medium
In both 2D and 3D experiments, two different culture conditions were explored for the adipogenic differentiation of preadipocytes: use of conventional adipocyte differentiation medium (DM) and use of direct coculture of adipocytes with preadipocytes in a TW plate. DM provides a standard, proven way of differentiating preadipocytes into adipocytes, while the TW condition was investigated because it may better mimic the complex differentiation stimuli provided in vivo. Use of direct coculture of adipocytes to stimulate adipogenic differentiation of preadipocytes in 3D in vitro environments has not previously been described.
The conventional adipocyte DM adapted from Cho et al. 34 consisted of phenol red-free Dulbecco's modified Eagle's medium:Ham's F12 supplemented with 3% fetal bovine serum, 2 mg/mL L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, biotin (33 μM), pantothenate (17 μM), human recombinant insulin (1 μM), dexamethasone (1 μM), isobutylmethylxanthine (0.1875 mM), and indomethacin (0.2 mM). The last two drugs were in the medium during only the first 3 days of culturing. TW plates were used to generate a coculture system of adipocytes and preadipocytes. TW inserts (polytetrafluoroethylene, (PTFE) 0.4 μm pore size) in 24-well plates were coated with collagen by soaking the inserts in a 28 μg/mL collagen solution overnight at 37°C under constant shaking. Mature adipocytes were attached to the undersurface of the TW inserts by pipetting 250 μL of a 10 × 106 cells/mL adipocyte suspension into a well and then placing the insert into the well. Since adipocytes float, they attached to the underside of the insert. After 18–24 h, these inserts with their attached adipocytes were placed in wells containing either the 2D or 3D preadipocyte cultures; this experimental setup is depicted in Figure 1.
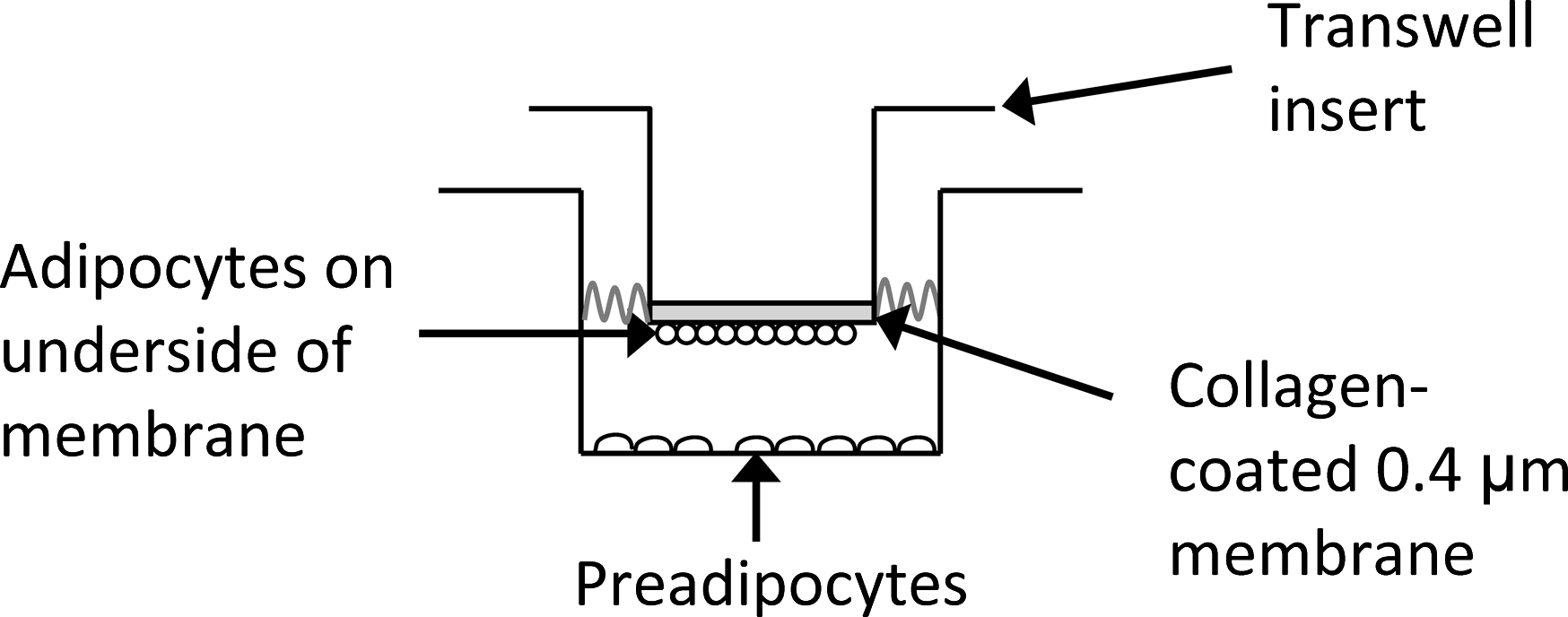
Experimental setup for preadipocyte/adipocyte transwell (TW) coculture condition. Because adipocytes are buoyant, they adhere to the collagen-coated underside of the TW insert.
Adipogenic differentiation—2D culture
First, the effects of the culture medium conditions on the adipogenic differentiation of preadipocytes were analyzed in a 2D environment. Preadipocytes were harvested and cultured as described above, and then seeded onto 24-well plates at a density of 1 × 105 cells/cm2. After treatment with DM or coculture with adipocytes in a TW, the adipogenic differentiation of the cells was analyzed via the quantification of several markers (leptin, lipolysis, adiponectin, and Oil Red O), which are described in subsequent sections. Samples were harvested at time points of 4–5, 9–10, 12–13, and 15–16 days, with medium changes every 2–3 days. Negative and positive controls consisted of preadipocytes or mature adipocytes, respectively, cultured in normal growth medium.
To account for glycerol, leptin, and adiponectin released by mature adipocytes in the coculture condition, mature adipocytes were attached to collagen-coated TW inserts (see above) and cultured with 300 μL of basal medium (in the absence of preadipocytes). The medium was collected every 3 days for 15 days, and the values for glycerol, leptin, and adiponectin (n = 12) were subtracted from TW sample values.
Adipogenic differentiation—3D cell encapsulation
Preadipocytes or freshly isolated adipocytes (positive control) were encapsulated in diacrylated PEG (PEG-DA) or methacrylated HA (HA-MA) scaffolds at a final concentration of 10 × 106 cells/mL. Methods outlined by Mann et al. 45 were followed for the acrylation of 3.4, 6, and 8 kDa PEG; each product was analyzed via 1H NMR to confirm acrylation. Encapsulation was performed by combining 1.5 mL of sterile-filtered 20 wt% PEG-DA, 0.5 mL of a 40 × 106 cells/mL suspension, and 0.1 mL of 1% Irgacure-2959 photoinitiator (Ciba Specialty Chemicals, Basel, Switzerland). This solution (final concentration of 15 wt% PEG-DA) was placed in a 1-mm-thick mold and exposed to UV light (365 nm and 2 mW/cm2) for 5–7 min, producing hydrogels that were then cut into 12-mm-diameter discs using a sterile cork-borer. HA-MA was synthesized via the methods described by Smeds et al., 46 and cells were encapsulated in a similar fashion, except that the final HA-MA concentration in the gels was 2 wt%.
Cell-seeded gels were placed in 24-well plates, incubated at 37°C, 5% CO2, and treated with the differentiation conditions outlined in a previous section (DM or TW). At time points of 4–5, 9–10, 12–13, and 15–16 days, culture medium was collected, and scaffolds were harvested in an assay-specific manner to quantify various adipogenic markers. The negative and positive controls consisted of preadipocytes or mature adipocytes, respectively, cultured in the PEG- or HA-based scaffolds with normal growth medium.
Lipolysis, leptin, and adiponectin assays
A lipolysis assay kit (ZenBio, Research Triangle Park, NC) was used to determine the amount of glycerol released into the culture medium by differentiating preadipocytes. Medium was collected, reacted with the glycerol reagent A, and the absorbance at 540 nm was measured (Synergy HT plate reader; Bio-Tek Instruments, Winooski, VT) and compared against glycerol standards.
Leptin and adiponectin ELISA kits (RayBiotech, Norcross, GA) were used to assess the amount of leptin and adiponectin released at various time points by differentiating preadipocytes. The assays quantitatively detect the amount of human protein (leptin or adiponectin) present in the cell culture medium via a sandwich-style ELISA, with standard curves generated via absorbance readings for known concentrations of leptin and adiponectin.
Oil Red O staining
Oil Red O staining was performed to qualitatively examine adipogenesis, as Oil Red O stains lipid deposits red. For 2D experiments, the wells were fixed in 10% formalin, washed with 60% isopropanol, allowed to dry completely, and then stained with Oil Red O working solution (3:2 Oil Red O:diH2O) for 10 min. Wells were then washed multiple times, counterstained with Gill's hematoxylin, and washed again.
The Oil Red O protocol was similar for the 3D hydrogels, except the hydrogels did not have a counterstain performed with Gill's hematoxylin and the Oil Red O was extracted for absorbance quantification by adding 0.5 mL of 4% Nonidet P-40 to each hydrogel. The gels were incubated for 1 h in extraction buffer, and the absorbance of the solution was measured at 540 nm. For controls, undifferentiated preadipocytes and mature adipocytes in each type of gel were stained with Oil Red O. Since there was some background staining of the gels with Oil Red O, gels with no cells were stained and extraction was performed with 4% Nonidet P-40. These values were subtracted from sample extractions to account for background staining.
Statistical analysis
Raw data were analyzed via ANOVA with a Tukey HSD posttest, and p-values <0.05 were considered statistically significant. All data are presented as mean ± standard deviation.
Results
Preadipocyte and adipocyte viability
The viability of preadipocytes was 82.1% from the trunk, 80.9% from the extremities, and 72.4% form the breast (p > 0.05). The viability of adipocytes was 80.7% from lipoaspirated fat and 83.1% from excised fat (p > 0.05). All adipocytes used for experiments were passage 0, and preadipocytes were used from passages 0 to 4. To confirm that the preadipocytes expressed stem cell markers, passage 4 preadipocytes were analyzed using fluorescence-activated cell sorting. The preadipocytes were positive for CD73, CD44, CD54, and CD29, and negative for CD34 and CD31.
Adipogenesis in 2D environments
Preadipocytes were differentiated into adipocytes under two conditions: adipogenic DM and direct coculture with TW inserts containing mature adipocytes. Quantitative biochemical assays were performed on samples collected at various time points and controls consisting of preadipocytes or mature adipocytes grown in basal medium. Qualitatively, cultures that yielded high levels of glycerol, leptin, or adiponectin were observed to change morphology from a fibroblastic to a spherical shape and accumulate lipid vacuoles within 10–14 days, which is also indicative of adipogenic differentiation. 37
Figure 2 compares the concentration of glycerol released by preadipocytes differentiated in the DM or TW environment. Glycerol is released by differentiating preadipocytes and mature adipocytes. In general, the DM condition did not stimulate as much glycerol release as the TW condition, although both conditions stimulated glycerol levels above that of the undifferentiated preadipocyte controls (p < 0.05). The preadipocytes in the TW culture had a peak glycerol release at days 9–10 (51.5 μM; no statistical difference from positive adipocyte control). By days 12–13, the TW cultures had significantly higher glycerol release activity than the DM-treated cultures.
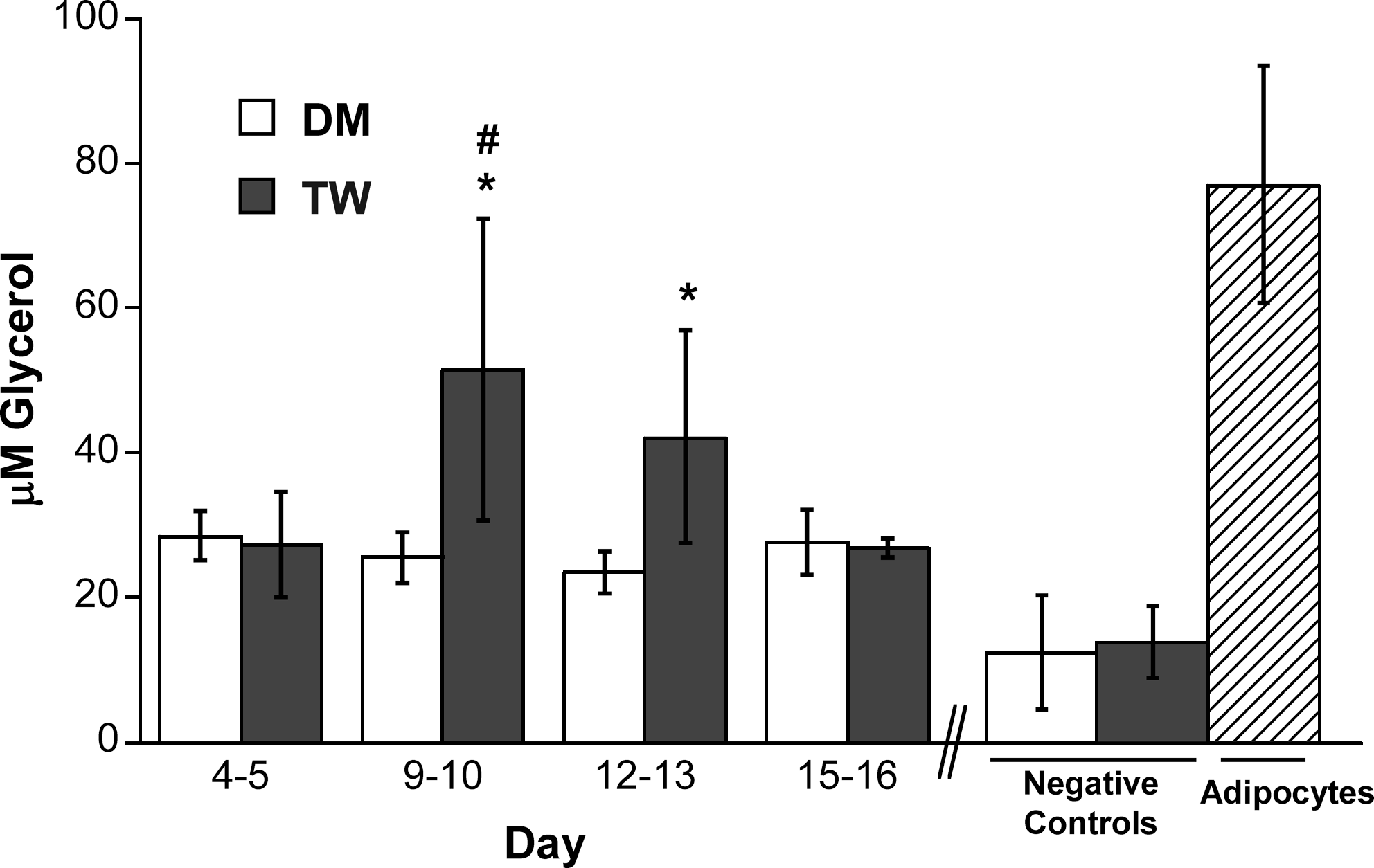
Glycerol release by preadipocytes grown in two types of differentiation media in a two-dimensional (2D) environment for 16 days (DM, differentiation medium; TW, transwell coculture). *p < 0.05 compared to negative control (undifferentiated preadipocytes); #p > 0.05 compared to positive control (mature adipocytes).
Leptin is a 16 kDa protein hormone released by differentiating preadipocytes and mature adipocytes, and is a specific indicator of adipogenic differentiation.37,47 Figure 3 demonstrates that preadipocytes differentiated in the presence of mature adipocytes produced significantly more leptin at days 15–16 than those cultured in DM (TW, 496 pg/mL vs. DM, 263 pg/mL; p < 0.05). Leptin levels in the TW condition at the final time point were statistically similar to those found in the positive control. Production of adiponectin, another important adipokine released by maturing adipocytes,37,47 followed similar trends to those seen for leptin, wherein adiponectin in the TW group was higher than in the DM group and almost equal to that seen in the positive control (Fig. 4). Lastly, Oil Red O staining of 2D differentiated preadipocytes was performed, and staining was noted in approximately 30% of the cells at days 15–16 in both DM and TW conditions (not shown).
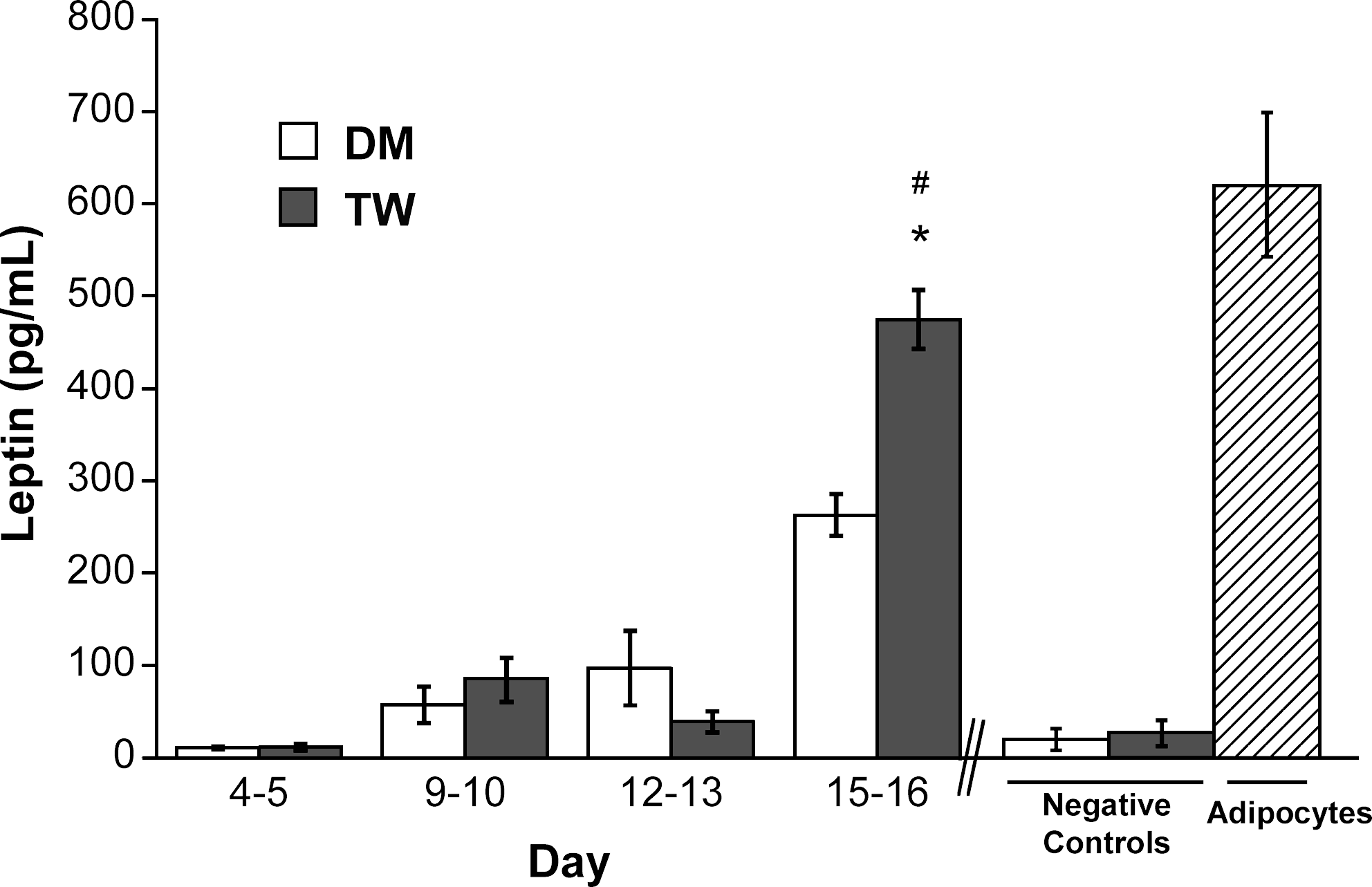
Leptin release by preadipocytes grown in two types of differentiation media in a 2D environment for 16 days (DM, differentiation medium; TW, transwell coculture). *p <0.05 compared to negative control; #p > 0.05 compared to positive control.
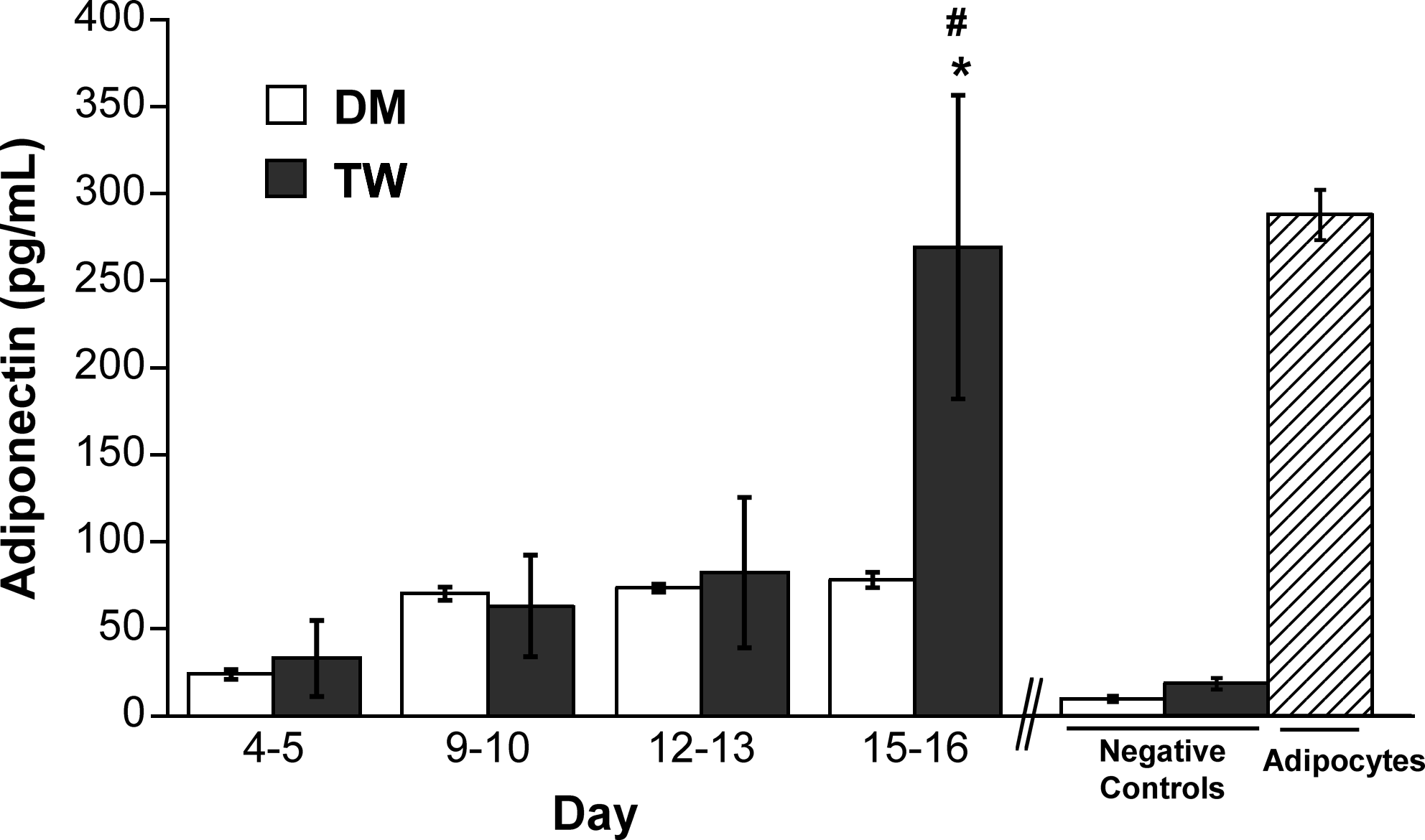
Adiponectin release by preadipocytes grown in two types of differentiation media in a 2D environment for 16 days (DM, differentiation medium; TW, transwell coculture). *p < 0.05 compared to negative control; #p > 0.05 compared to positive control.
Adipogenesis in 3D environments
After performing differentiation of preadipocytes to adipocytes in 2D environments, the differentiation of preadipocytes in 3D scaffolds was tested under the same culture conditions: traditional DM and coculture with TW inserts containing adipocytes. Further, scaffold physical properties were also altered to evaluate how properties such as stiffness affect adipogenic differentiation.
Glycerol
At all time points shown in Figure 5, significantly more glycerol was produced when the preadipocyte-seeded scaffolds were cultured in the presence of mature adipocytes (TW condition) rather than DM. By the earliest time point, the TW condition had emerged as a leader in stimulation of glycerol release, although no significant differences existed between the types of scaffolds used. At days 12–13, one scaffold condition—8 kDa PEG cultured in the TW condition—started to separate itself from the other scaffolds, with a significantly higher glycerol concentration (434.3 μM, p < 0.05). By days 15–16, the TW preadipocytes in the 3.4 and 6 kDa PEG had increased production of glycerol to catch up with the 8 kDa PEG, with glycerol values for these three scaffolds in the range of 800–1200 μM, which approached levels obtained in the positive control (∼1400 μM). Differences in glycerol production between scaffold conditions could not be attributed to differences in each scaffold's ability to support cell function, as adipocytes thrived equally in all four types of scaffolds (as evidenced by approximately equal glycerol release by the positive controls). Although the HA scaffold performed well in the initial three time points, differentiation of preadipocytes in these scaffolds appeared to stagnate, leading to a low glycerol value at the final time point.
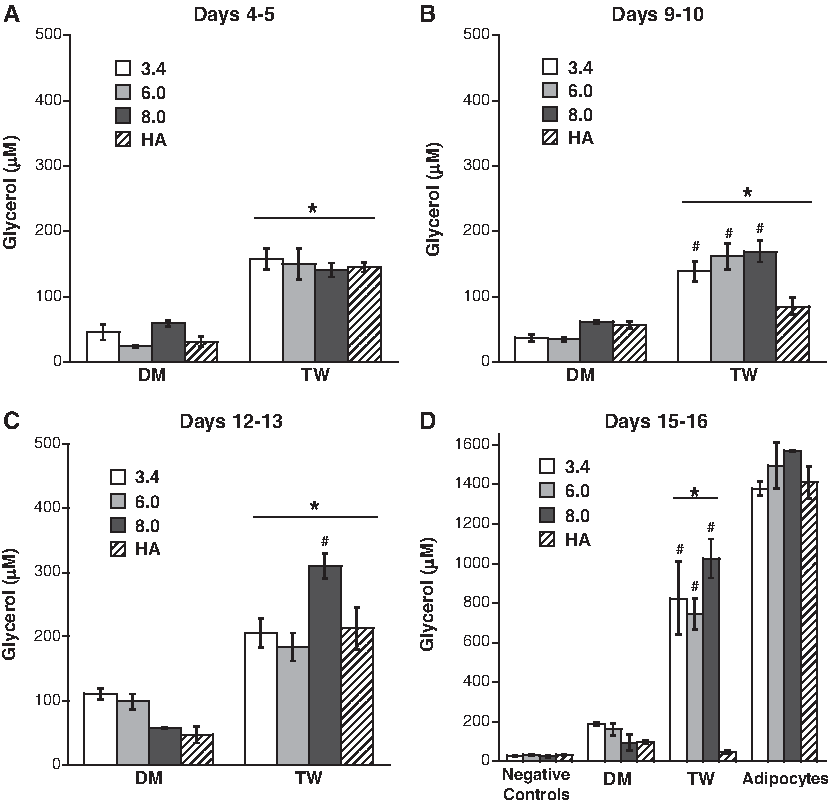
Glycerol release from preadipocytes cultured in DM or TW conditions in various three-dimensional (3D) scaffolds at time points of (
A comparison of Figures 2 and 5 also reveals significant differences in lipolysis between the 2D and 3D culture environments. Although glycerol release by undifferentiated preadipocytes (the negative control) was similar in both 2D and 3D environments, glycerol release by differentiating cells and mature adipocytes was substantially higher in the 3D scaffolds. These differences were evident by the first time point, whereupon the respective glycerol release by DM- and TW-treated preadipocytes in 3D scaffolds was 1.7- and 5-fold higher than in 2D. By the final time point, culture in 3D scaffolds resulted in roughly a 6-fold increase in glycerol release by cells cultured in DM and a 50-fold increase by cells in the TW condition relative to 2D conditions. Mature adipocytes also fared significantly better in the 3D scaffolds, with a 17-fold increase in glycerol release compared to the 2D culture condition.
Leptin
The leptin release from differentiated preadipocytes cultured within one of the three molecular weights of PEG (MW 3.4, 6, or 8 kDa) or HA scaffolds is shown in Figure 6. At the end of the time course, identification of the scaffold type that best facilitated leptin production was found to rely upon the type of differentiation treatment used. In other words, one type of scaffold (HA-MA) performed best in the DM group, while a different type of scaffold (8.0 kDa PEG-DA) performed best in the TW group. At days 15–16, the trend was that the DM condition supported higher release of leptin than the TW for all scaffolds tested. In the positive controls, leptin production by mature adipocytes varied slightly across scaffold conditions, but all samples were within a narrow range of 600–720 pg/mL.
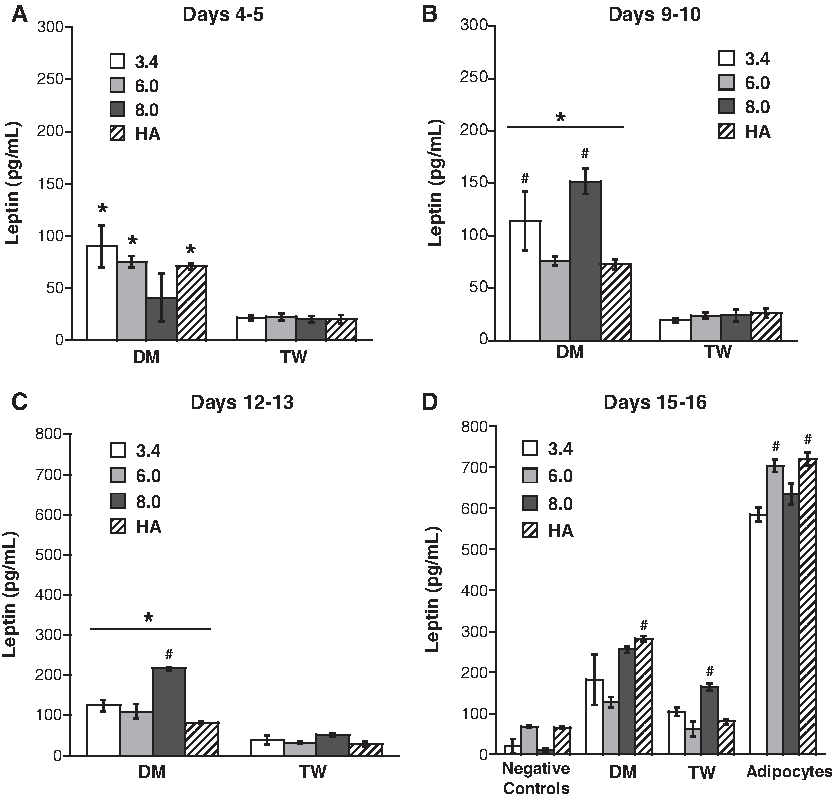
Leptin release from preadipocytes cultured in DM or TW conditions in various 3D scaffolds at time points of (
In comparing the adipogenesis in 3D (Fig. 6) to the 2D results in Figure 3, it is seen that production of leptin by preadipocytes in 3D was somewhat improved relative to culture in the 2D environment. Particularly in the DM treatment condition, culture in 3D led to a more rapid onset of increased leptin production, although, by the final time point, the values for leptin production by DM-treated preadipocytes and mature adipocytes did not significantly change in going from a 2D to a 3D environment.
Adiponectin
As shown in Figure 7, adiponectin release from the preadipocytes cultured in 3.4 and 8 kDa PEG materials for 15–16 days was higher than that seen in 6 kDa PEG and HA scaffolds for both DM and TW conditions (p < 0.05). At the final time point, adiponectin values for all 3.4 and 8 kDa conditions were not statistically different from those obtained for the positive control. However, for the 6 kDa and HA scaffolds, only the TW-treated scaffolds yielded adiponectin levels that were similar to those obtained in positive controls.
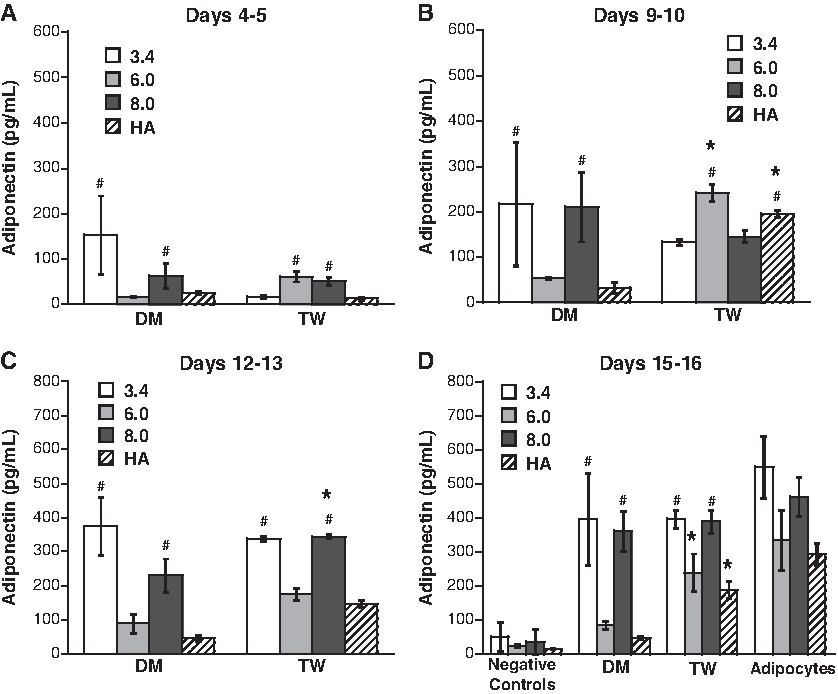
Adiponectin release from preadipocytes cultured in DM or TW conditions in various 3D scaffolds at time points of (
Quantification of adiponectin was the only experiment in which significant differences were noted in the ability of the different scaffolds to support the function of mature adipocytes. The 3.4 kDa (549 pg/mL) and 8 kDa (462 pg/mL) scaffolds performed equally well in supporting adiponectin release by mature adipocytes, but both of these conditions had higher adiponectin release than the 6 kDa PEG and HA conditions. Even accounting for the decreased adiponectin release in 6 kDa PEG and HA conditions in the positive controls, the 6 kDa PEG and HA scaffolds still did not perform as well as the 3.4 and 8 kDa PEG gels across the different medium conditions, as indicated by the only modest increase in adiponectin above negative controls for 6 kDa and HA scaffolds in DM.
With respect to the impact of culture dimensionality, a comparison of Figures 4 and 7 demonstrates that culture in a 3D environment was more conducive to adiponectin production than the 2D environment. Undifferentiated preadipocytes maintained similar levels of adiponectin production in both 2D and 3D, but differentiating and mature preadipocytes tended to produce higher amounts of adiponectin when cultured in 3D. Specifically, production of adiponectin increased much more rapidly in 3D scaffolds, reaching a level that was 5-fold higher than on 2D for the DM condition, and 1.6-fold higher than on 2D for the TW condition by the final time point. Mature adipocytes cultured in 3D scaffolds also exhibited adiponectin production that was 1.8-fold higher than in the 2D culture environment.
Oil Red O
Mirroring the adiponectin results seen in Figure 7, both 3.4 and 8 kDa PEG conditions were accompanied by the highest lipid content, as evidenced by Oil Red O absorbance (Fig. 8). Both of these scaffold conditions in the DM and TW treatment groups approached lipid content values obtained for positive controls. All values in Figure 8 are reported as a percentage of the absorbance obtained for each scaffold-specific positive control (i.e., mature adipocytes cultured in the various scaffold types were equivalent to 100% on the plot). The scaffolds themselves took up variable amounts of background stain, and this was subtracted from the readings for all corresponding conditions. The 3.4 kDa PEG, 8.0 kDa PEG, and HA constructs all stained greater than ∼2-fold compared to their respective negative controls.
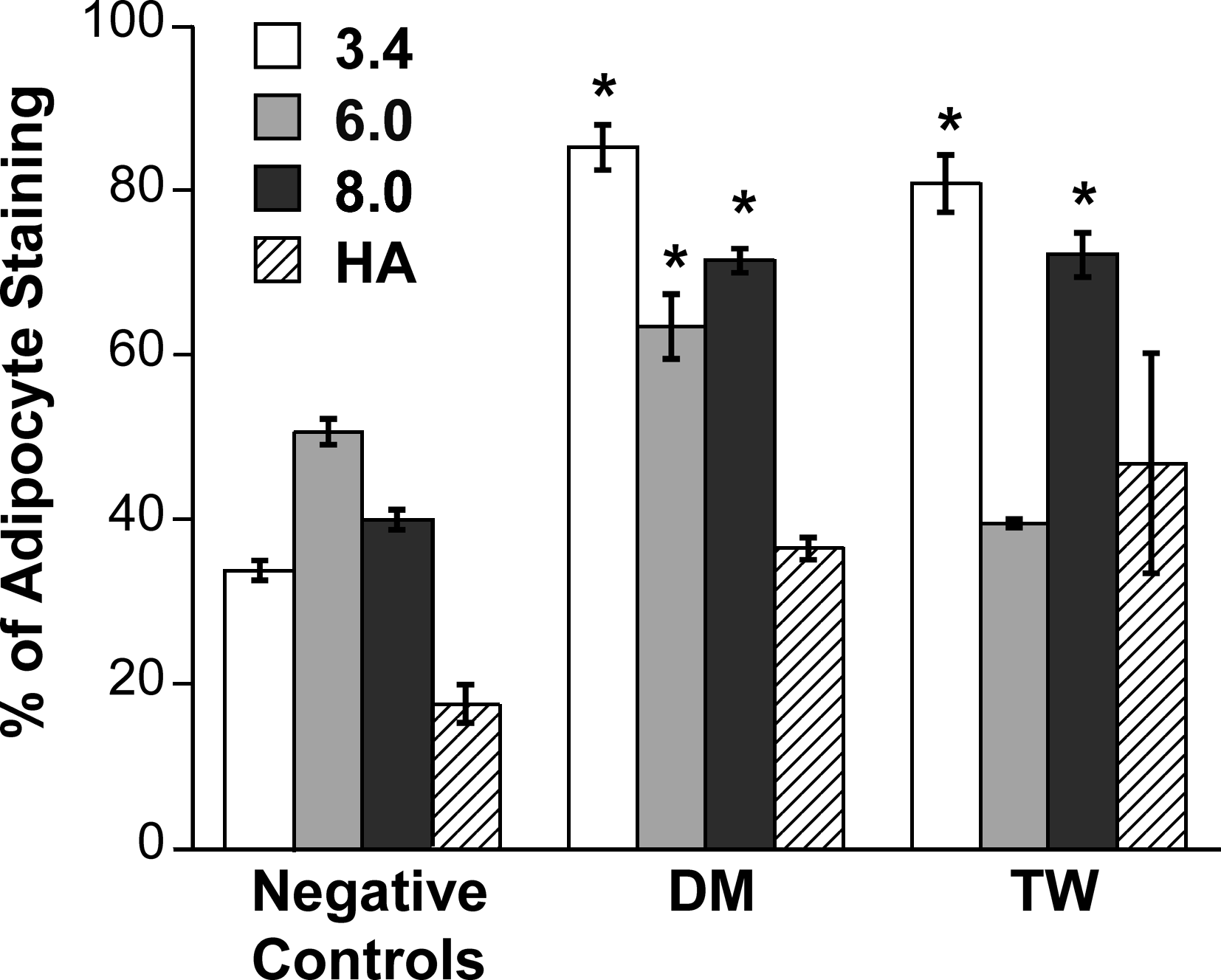
Oil Red O staining and extraction of preadipocyte-seeded 3D scaffolds after 16 days of treatment with differentiation stimuli. The amount of staining was measured at an optical density of 450 nm and is expressed as a percentage of the amount of mature adipocyte staining in matched scaffold conditions. *p < 0.05 compared to other scaffolds within same medium condition.
Discussion
Studies of preadipocyte differentiation were first performed in 2D to achieve two main objectives: (1) to compare the efficacies of two different culture conditions in stimulating adipogenic differentiation and (2) to discover whether the same culture condition that best stimulated adipogenesis in 2D would perform similarly in a 3D environment. Overall, the adipogenic performance of cells cultured in the 2D TW treatment condition was substantially improved over that of cells cultured in standard DM. In all assays, only the TW-treated condition approached levels of adipogenic marker expression observed in the positive control. The glycerol, leptin, and adiponectin data indicate that, in 2D culture, the TW preadipocytes differentiated at an earlier time point and more effectively when compared against the preadipocytes in the DM. These results are notable in that the use of DM is currently the most common method for achieving adipogenic differentiation in vitro, while the current work found DM to be far less supportive of differentiation than the TW condition.
It is also worth noting that only two previous studies48,49 have employed direct coculture of adipocytes with preadipocytes to stimulate differentiation, likely due to the challenge of culturing the buoyant adipocytes. Both of these studies were done only in 2D, meaning that 3D adipose tissue engineering experiments have not previously been conducted in coculture with mature adipocytes in vitro. Also, the modified TW setup described in this report has addressed some of the disadvantages of the coculture techniques used in the earlier 2D studies.48,49 Although the direct in vitro coculture of adipocytes with preadipocytes has not received much attention in the past, our results indicate that this method of differentiation can be more effective than other differentiation stimuli for in vitro adipogenic differentiation. Moreover, such coculture is more physiologically relevant than many other differentiation techniques, as it is one step closer to mimicking the environment that engineered tissues will experience in vivo. As a closer mimic of the in vivo environment, the use of TW culture may be beneficial for evaluating and optimizing scaffold properties and conditions in vitro before in vivo implantation.
After studying differentiation treatments in 2D experiments, the experimental setup was translated to a 3D preadipocyte culture environment using various types of biomaterial scaffolds. Previous work indicates that a mechanical scaffold is necessary for in vivo adipose tissue engineering because preadipocytes grafted alone do not generate adipocytes. 22 PEG-based hydrogels can be generated with a variety of different polymer chain molecular weights and concentrations, thereby imparting different ranges of mechanical strength, pore size, and water content upon the scaffold. A few groups have combined bone marrow–derived human stem cells with PEG-DA gels to engineer adipose tissue,50,51 but our understanding of how adipose-derived stem cells interact with the scaffold environment under different differentiation conditions has been quite limited. Patel et al. 52 encapsulated rat preadipocytes in 6 kDa PEG-based scaffolds with and without tethered peptide sequences. They found that preadipocyte death occurred over 1 week if the PEG was nondegradable and/or did not contain peptide sequences. 52 This observation is different from the results described herein, where the preadipocytes in 3D PEG scaffolds were >90% viable at day 15 according to a Live/Dead Assay (data not shown). Moreover, all of the scaffold types used in this work were able to support the viability and healthy function of mature adipocytes, which continued to secrete high amounts of relevant adipogenic markers. Thus, these current findings indicate that the 3D scaffold conditions supported the culture of viable, differentiation-capable precursor cells, as well as the culture of viable, functional mature cells, which represented our target. Possible reasons for the differences between our findings and those of Patel et al. could be that we used human preadipocytes, different seeding densities, and different growth conditions.
Meanwhile, several recent papers have described the use of HA-based scaffolds for adipose tissue engineering, although with varied results.27–31,53 Our results did not find the photocrosslinked HA-MA scaffolds to be particularly supportive of adipogenic differentiation; in many cases, the level of adipocyte-specific marker expression by cells in the HA scaffolds was not statistically higher than that found for the negative control. Although others have found HA-based scaffolds to be supportive of adipogenesis, these results may also vary with the type of scaffold structure used (i.e., hydrogel, sponge, or fibrous network), 31 as well as whether the cells were predifferentiated before seeding within the scaffolds. However, the results of the current study are consistent with a recent publication finding that a HA-based scaffold did not support the formation of mature adipocytes from preadipocytes upon in vivo subcutaneous implantation in human subjects. 53
As mentioned earlier, one purpose of these studies was to evaluate whether 2D observations and optimizations could be directly applied to 3D environments, particularly with respect to identifying culture conditions that support adipogenic differentiation. In general, the production of adipogenic markers for all culture conditions (including the mature adipocyte control) was lower in the 2D environment than in the 3D environment, which is not surprising given that 3D culture is more physiologically relevant. Production of leptin was the least affected by changes in culture dimensionality, but both glycerol release and adiponectin production were dramatically increased in 3D relative to 2D conditions. Culture dimensionality also impacted adipogenesis trends related to the application of the two different differentiation stimuli—DM versus TW. In 2D, the impact of altering the DM was fairly consistent across the different adipogenic markers, with TW coculture stimulating significantly increased differentiation. In 3D, the results became much more complex, although the TW coculture was more universally supportive of the different scaffold conditions. For instance, all TW-treated scaffold types reached adiponectin levels similar to controls, but only select types of DM-treated scaffolds were supportive of differentiation (namely, 3.4 and 8 kDa PEG, but not 6 kDa PEG or HA). Thus, there were several commonalities between 2D and 3D results, as TW treatment generally correlated with high levels of adipogenic differentiation in both 2D and 3D environments. However, selected scaffolds in the DM condition were able to perform as well as the scaffolds in the TW group, meaning that TW was not singularly the optimal differentiation condition in 3D culture. In contrast, in 2D, DM treatment did not achieve glycerol, leptin, or adiponectin levels that were close to those seen in the positive control. The observation that DM performed poorly in 2D adipogenesis but almost as well as the TW condition for 3D adipogenesis represents further evidence that the 3D culture itself provides an environment that is more conducive to adipogenesis.
There have been several recent studies directed at investigating how the mechanics of the culture environment impact stem cell differentiation. 54 In general, softer materials are more conducive to differentiation into soft tissue types, such as adipose. Thus, the current work was performed with scaffolds that represented a range of mechanics: elastic moduli ranging from 2 to 40 kPa, where the elastic modulus of native adipose tissue is ∼6 kPa. 55 Overall, the 3.4 and 8 kDa PEG-based materials tended to be more supportive of adipogenic differentiation across all medium treatment conditions. The reasons for this result—in particular, the underperformance of the intermediate PEG MW (6 kDa), which has mechanical properties in between the high-performing 3.4 and 8 kDa conditions—are not clear. The underperformance of the 6 kDa condition occurred in several different experiment trials and was not related to variations in cell harvests or problems with polymer formulation or crosslinking. Moreover, for the most part, mature adipocytes thrived in 6 kDa PEG scaffolds, producing high amounts of glycerol and leptin, indicating that these scaffolds were appropriate for culture of mature cells, but not as appropriate for differentiation of immature cells.
Conclusions
Although the use of preadipocytes as a cell source for adipose tissue engineering is attractive and practical, our knowledge of the optimal differentiation conditions for these cells, particularly in 3D environments, has been limited. The present work communicates several significant findings, including (1) establishment of a preadipocyte/adipocyte coculture technique that stimulates superior adipogenesis compared to traditional DM, (2) a direct comparison of adipogenesis in 2D versus 3D conditions showing marked upregulation of adipogenesis in the latter environment, (3) evidence that alterations in 3D scaffold physical properties significantly impact the ability of preadipocytes to undergo adipogenic differentiation, but only minimally alter the ability of mature adipocytes to function, and (4) generation of culture and scaffold conditions that can stimulate levels of adipogenic marker production similar to those found in positive controls. Of the scaffold conditions investigated here, the 8 kDa PEG-based material was found to support the greatest adipogenic differentiation both in vitro and in vivo. While truly optimized scaffolds for adipose tissue engineering will likely need to be multicomponent environments containing bioactive molecules,24,25,33 the conditions explored herein provide guidance and new information with respect to scaffold design and in vitro differentiation conditions.
Disclosure Statement
No competing financial interests exist.